Мезельсон-Сталь тәжірибесі және прокариоттар мен эукариоттардағы ДНҚ репликациясы: денатурация, ренатурация және гибридизация


Мезельсон-Сталь тәжірибесі, прокариоттардың және эукариоттардың ДНҚ репликациясы, ДНҚ денатурациясы және ренатурациясы
Орындағандар: Осмонали Б. Б., Хамза А., Кисимова Г., Дузелова З., Әбілдашева А., Абдығапарова Г., Бейбітова Н., Бекболатова Г., Жалел М., Қуан Л., Ниятхан А., Сейтжан А., Оралова С.
Қабылдаған:Жунусбаева Ж. К.

Жоспар:
Мезельсон-Сталь тәжірибесі
Прокариот ДНҚ-ның репликациясы
Репликацияға қатысатын белоктар мен ферменттер
ДНҚ репликациясы
Эукариот ДНҚ-ның репликациясы
Эуариоттық ДНҚ-полимеразалар
Эукариоттық клеткалардағы ДНҚ репликациясының ерекшеліктері
Эукариоттардағы ДНҚ репликациясы
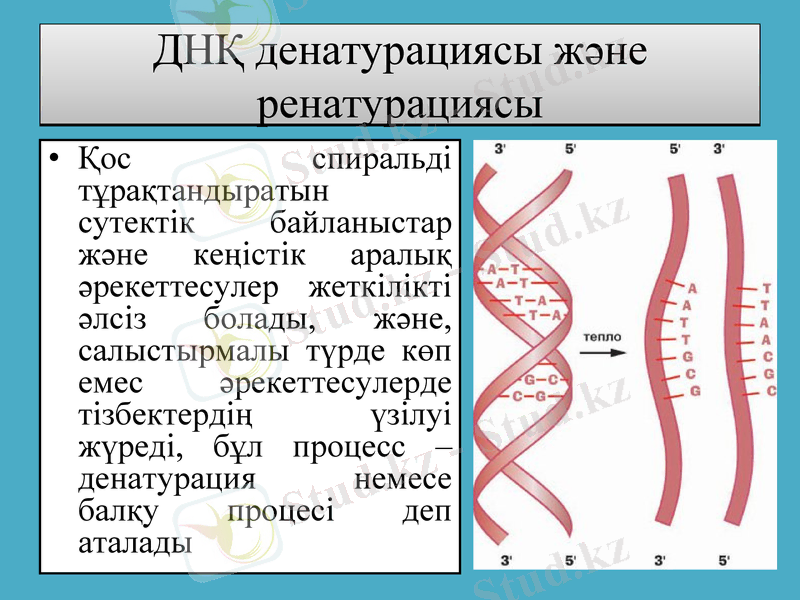
ДНҚ денатурациясы және ренатурациясы
ДНҚ-ДНҚ гибридизациясы
ДНҚ- РНҚ гибридизациясы

Мезельсон-Сталь тәжірибесі
1957 жылы Мэтью Мезелсон мен Франклин Сталь ДНҚ-ның болжамды құрылысын тексеру өте қисынды тәжірибе жүргізді.
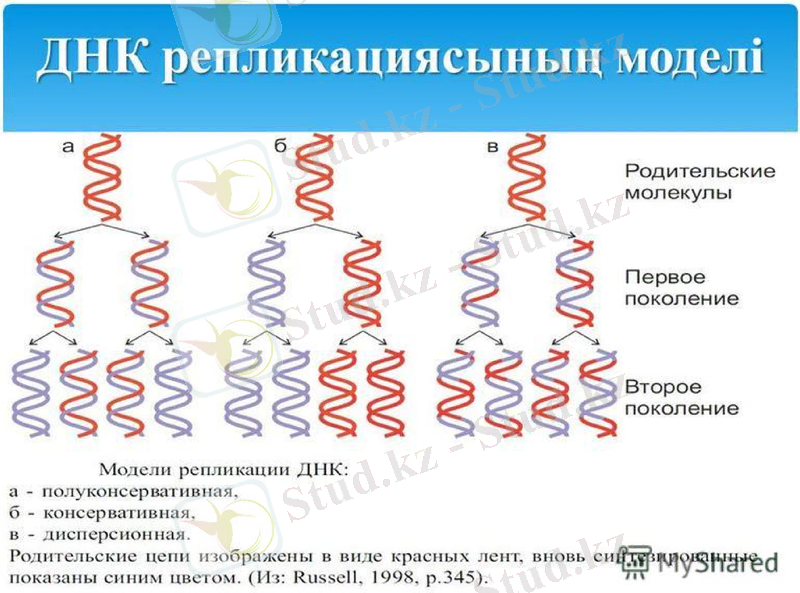

Олар ішек таяқшасы (Esherichia coli) бактериясын бірнеше ұрпаққа дейін құрамында азоттың ауыр тұрақты изотопы бар қоректік ортада өсірді. Азот көзі ретінде 15NH4Cl тұзын пайдаланды. Бірнеше ұрпақ бойы осындай азотпен қоректенгенде бактерия жасушасының ішіндегі құрамында азот атомдары бар барлық биомолекулалардың, оның ішінде ДНҚ-ның да салмағы кәдімгі азот 14N атомы бар қоректік ортада өсірілген бактериянікінен ауыр болып шығады. Әдетте, ауыр биомолекулалрдың молекулалық салмағын анықтау үшін олардың дайындамаларын хлорлы цезийдің тығыздығының градиенті арқылы үлкен жылдамдықпен центрифугалайды. Молекула неғұрлым ауыр болса - ол соғұрлым центрифуга ыдысының түбіне қарай жылдам батады.
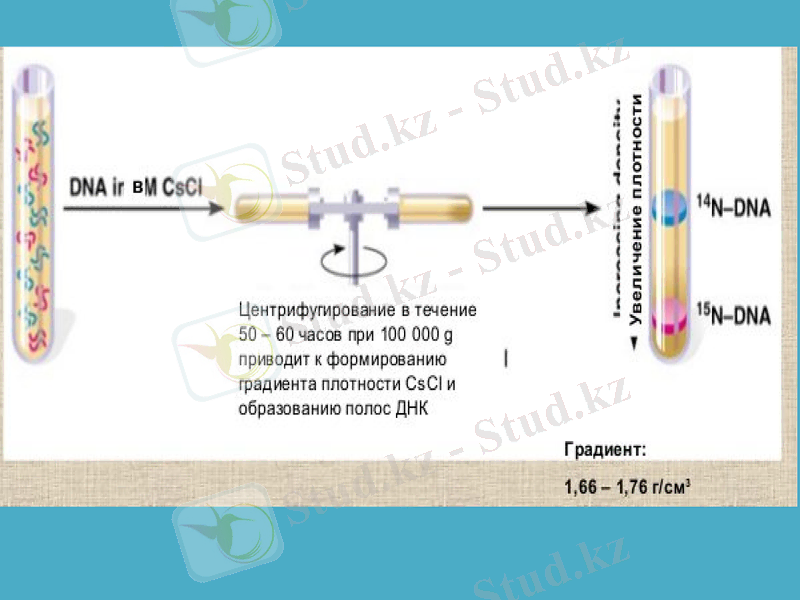

Месельсон мен Сталь осындай ауыр азотта өсірілген бактерияның ДНҚ-сын центрифугалағанда ауыр молекулалы ДНҚ-ның тек бір түрін анықтады. Одан кейін олар бактерияны құрамында кәдімгі жеңіл 14N және ауыр 15N азот атомдары бар ортада қысқалау мерзім ішінде өсірді (алынған бактерияны екінші ұрпақ деп атайық) . Ондай бактериялардан алынған ДНҚ-ны центрифугалағанда - бір ДНҚ фракциясы алынды және ол таза 15N-ортада өсірілген бактериянікінен жеңіл болып шықты.
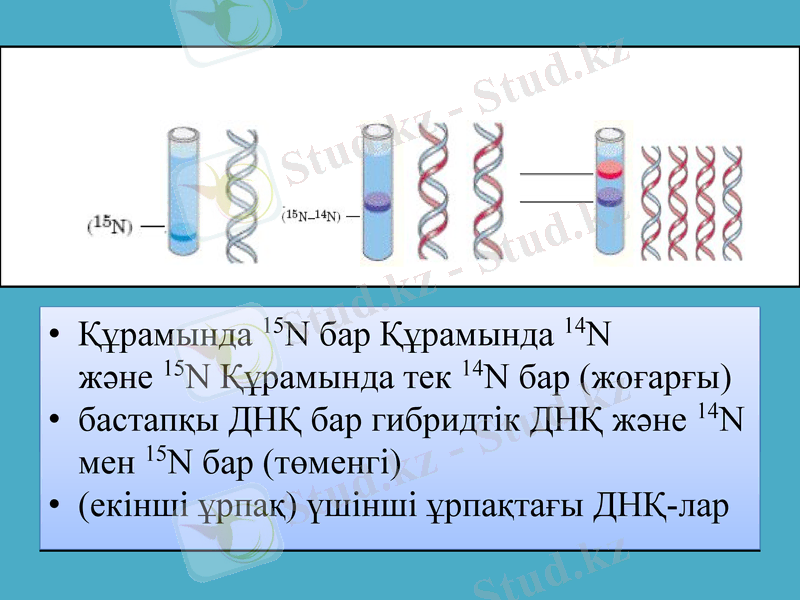
Құрамында 15N бар Құрамында 14N және 15N Құрамында тек 14N бар (жоғарғы)
бастапқы ДНҚ бар гибридтік ДНҚ және 14N мен 15N бар (төменгі)
(екінші ұрпақ) үшінші ұрпақтағы ДНҚ-лар

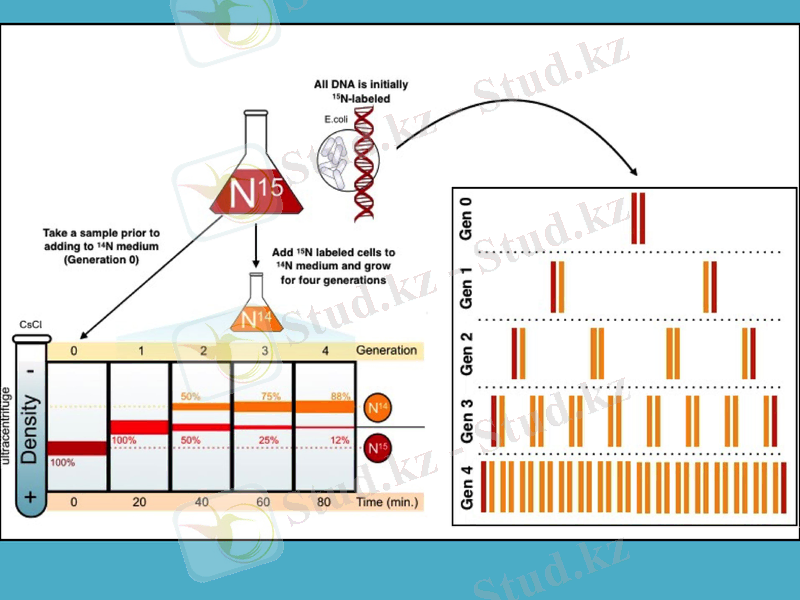

Ақырында ғалымдар келесі үшінші ұрпақты құрамында тек жеңіл 14N азот атомдары бар ортада бөлек, және құрамында екі азот атомының 14N және 15N түрлері бар ортада бөлек өсірді. Олардан алынған үшінші ұрпақтың ДНҚ-сы өте жеңіл және екінші ұрпақтағы секілді салмағы орташа екі түрлі болып шықты. Месельсон мен Стальдің алған нәтижелері бастапқы немесе аналық ДНҚ-ның әр тізбегінің бойында оған комплементарлы жаңа тізбектер синтезделетінін анық көрсетті. Сонымен, еселенудің алдындағы ДНҚ тізбектерінің әрбірі жаңа синтезделген тізбектер үшін матрица болып табылады.

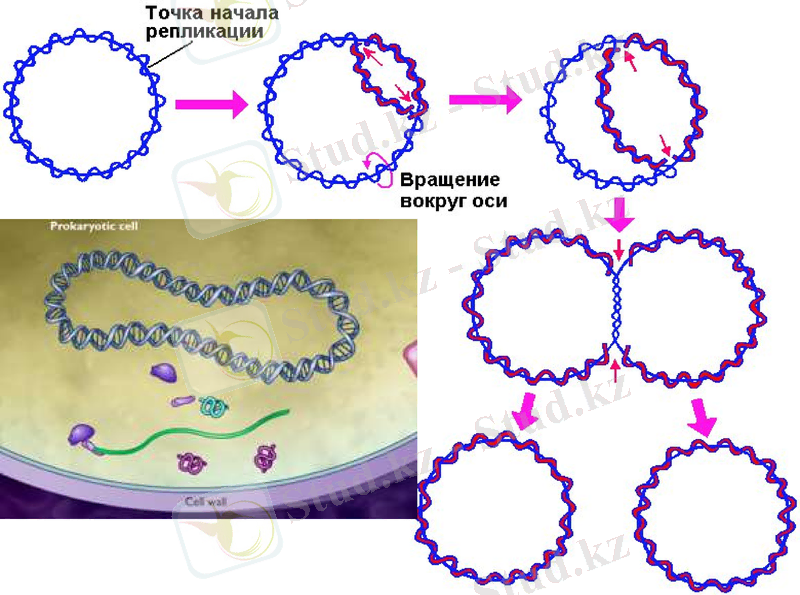
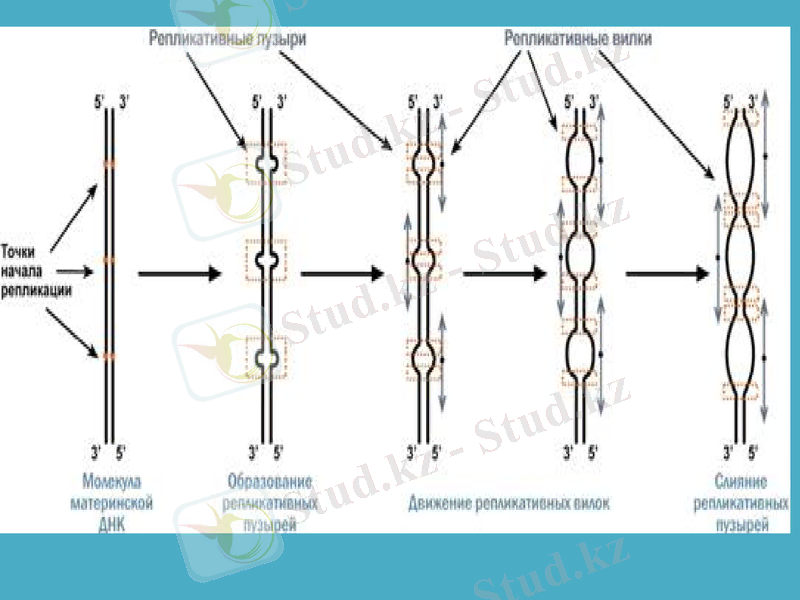
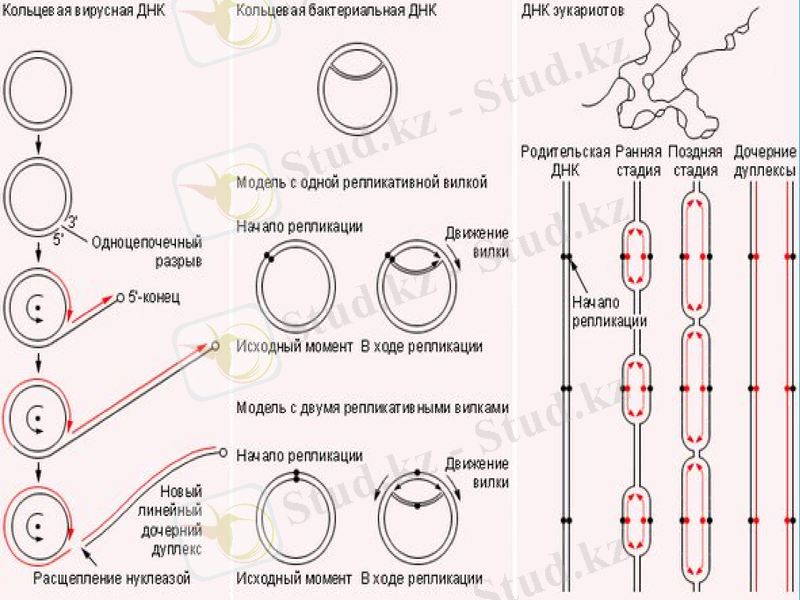
Кейінгі зерттеулер ДНҚ-ның еселенуі бірден екі бағытта жүретінін көрсетті. Сақиналы ДНҚ-ны радиоактивті 3Н тритиймен белгілеп, уақыт барысында өте жоғары шешімді электрондық микроскопты пайдаланып, бақылағанда репликацияның бір нүктеден басталатыны байқалды. Алғаш белгілі бір нүктеде ДНҚ-ның қос тізбегі бір-бірінен ажырайды. Сол нуктеден екі айыр пайда боладлы да, олар қарама-қарсы бағытта қозғалады. Пайда болған айырды рапликация айыры деп атайды. Жоғарыда ДНҚ-ның қос спиралын құрап тұрған жеке тізбектер бір-біріне антипараллельді деп айтылып кетті. Олай болса - әрбір тізбектің бойында жаңа тізбектер 5’-бағытынан 3’-бағытына қарай жүруі керек.
Прокариот ДНҚ-ның репликациясы
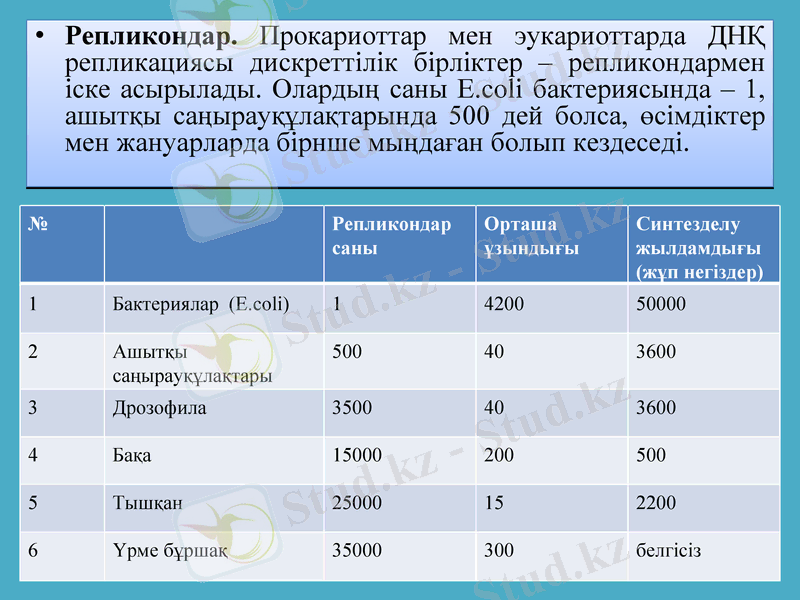
Репликондар. Прокариоттар мен эукариоттарда ДНҚ репликациясы дискреттілік бірліктер - репликондармен іске асырылады. Олардың саны E. coli бактериясында - 1, ашытқы саңырауқұлақтарында 500 дей болса, өсімдіктер мен жануарларда бірнше мыңдаған болып кездеседі.
№
Репликондар саны
Орташа ұзындығы
Синтезделу жылдамдығы (жұп негіздер)
1
Бактериялар (E. coli)
1
4200
5
2
Ашытқы саңырауқұлақтары
500
40
3600
3
Дрозофила
3500
40
3600
4
Бақа
15000
200
500
5
Тышқан
25000
15
2200
6
Үрме бұршақ
35000
300
белгісіз

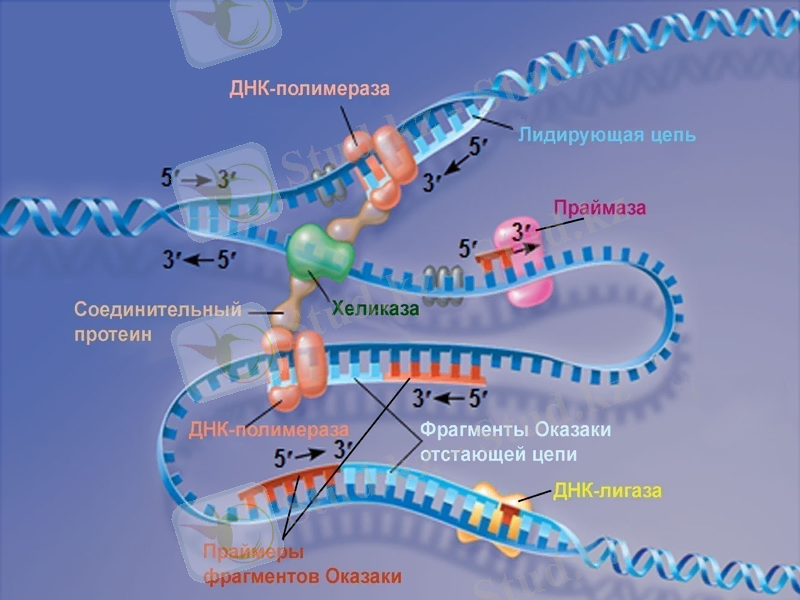
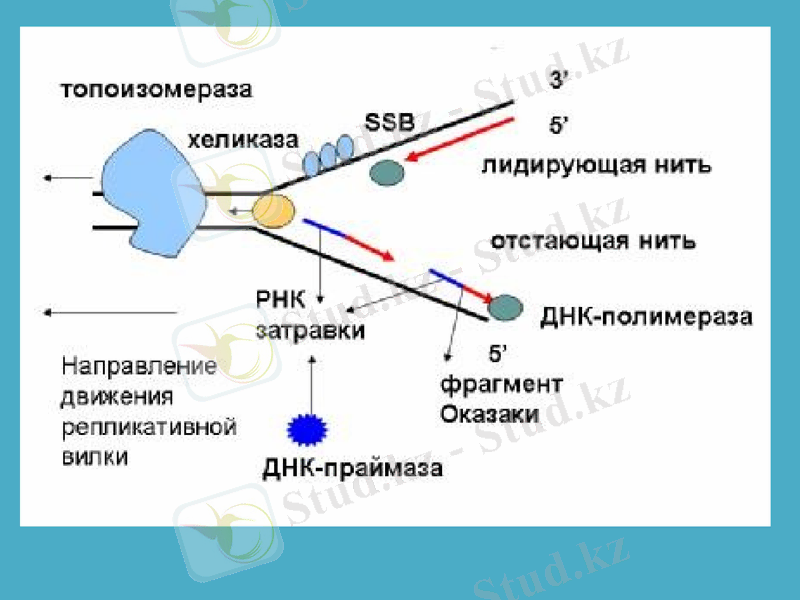
Сонымен, ДНҚ-полмераза-1 ДНҚ-ның жаңа тізбектерінің синтезін бастайтын қабілеті жоқ болып шықты. ДНҚ репликациясының басталуына қатысатын ферменттердің бірі - праймазадеп аталатын жасушаның арнайы РНҚ-полимеразаферменті екен, ол праймер деп аталатын қысқа, 10-60 нуклеотидтерден тұратын синтездейді, тек содан кейін ғана ДНҚ-ның синтезі басталады. Праймазалар құрылысы және арнайы қызметі бойынша бір-бірінен өзгеше болып келеді. Праймасома ферменттің каталитикалық қызметінде ерекше роль атқарады. Праймасома әртүрлі 7 суббірліктердің жиынтығы болып табылады. Арнайы n’ деген белоктың активтігінің нәтижесінде түзілген АТФ-ты пайдаланып, праймосома қалушы (отстающая) ДНҚ тізбегіне тез орын ауыстырады. Репликация айырына жақын жерде праймаза танитындай ДНҚ-ның арнайы екінші структурасының құрылуына қатысатын dna B және dna C белок комплекстері де прайсоманың құрамына кіреді.
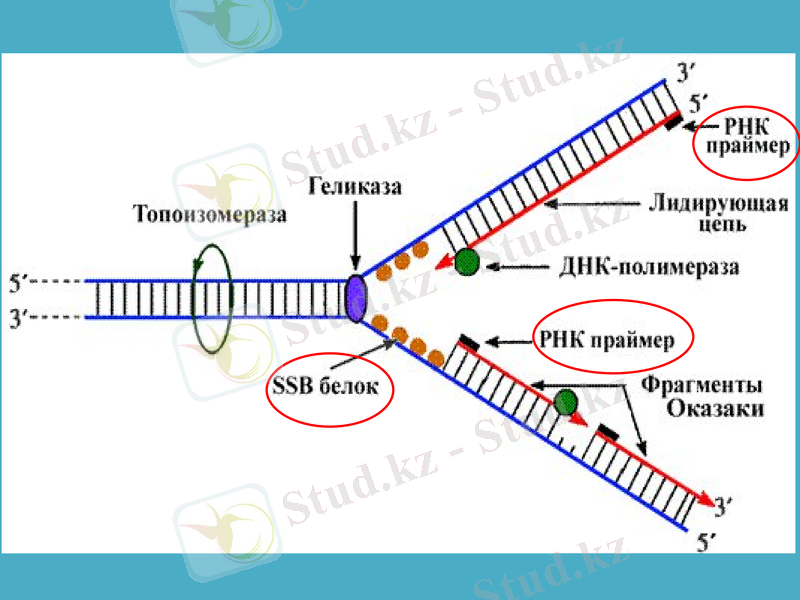

Жаңадан пайда болған ДНҚ-ның синтезін катализдайтын негізгі фермент ДНҚ-полимераза-3 болып табылады (дәлірек айтқанда, ол ДНҚ репликациясының ұзаруына жауап береді) . Ол - ДНҚ-полимеразаның өзінің және басқа белоктардың мультимерлік комплексі. Мысалы, ішек таяқшасының ДНҚ-полимераза-3 ферментінің құрамына 10 суббірлік кіреді. Репликация кезінде ДНҚ-полимераза-3-тің димерлік формасы жетекші (ведущая) және қалушы тізбектерінің бірлескен синтезін жүргізеді деген дәлелдер бар.
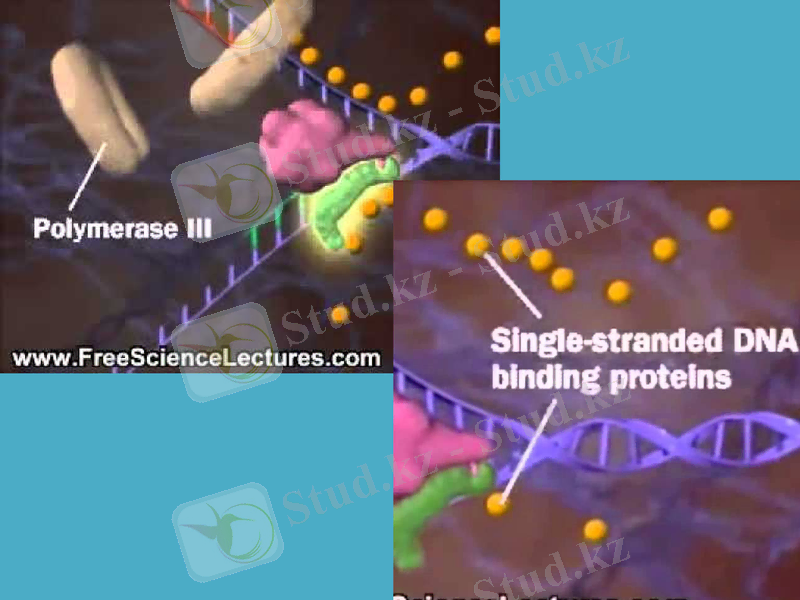
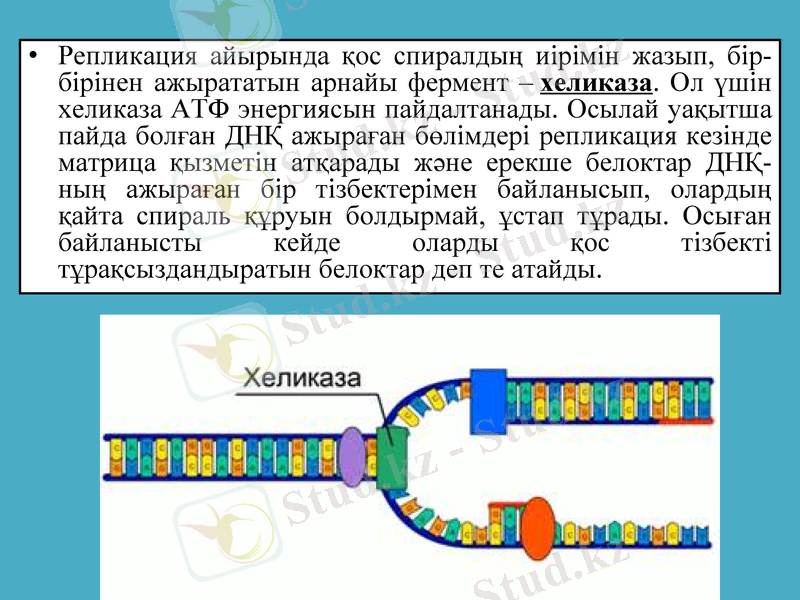
Репликация айырында қос спиралдың иірімін жазып, бір-бірінен ажырататын арнайы фермент - хеликаза. Ол үшін хеликаза АТФ энергиясын пайдалтанады. Осылай уақытша пайда болған ДНҚ ажыраған бөлімдері репликация кезінде матрица қызметін атқарады және ерекше белоктар ДНҚ-ның ажыраған бір тізбектерімен байланысып, олардың қайта спираль құруын болдырмай, ұстап тұрады. Осыған байланысты кейде оларды қос тізбекті тұрақсыздандыратын белоктар деп те атайды.
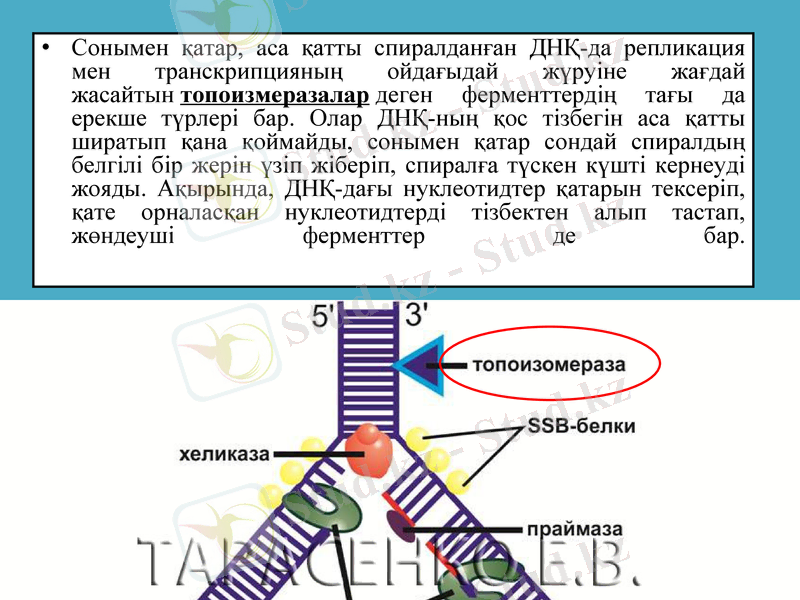
Сонымен қатар, аса қатты спиралданған ДНҚ-да репликация мен транскрипцияның ойдағыдай жүруіне жағдай жасайтын топоизмеразалар деген ферменттердің тағы да ерекше түрлері бар. Олар ДНҚ-ның қос тізбегін аса қатты ширатып қана қоймайды, сонымен қатар сондай спиралдың белгілі бір жерін үзіп жіберіп, спиралға түскен күшті кернеуді жояды. Ақырында, ДНҚ-дағы нуклеотидтер қатарын тексеріп, қате орналасқан нуклеотидтерді тізбектен алып тастап, жөндеуші ферменттер де бар.

Қазір ДНҚ-полимераза-1 бастаушы (затравка) рибонуклеотидтік праймердің бөлініп кетуін және сол босаған бөлікті толтыру процестерін катализдейтіні дәл анықталған. Ішек таяқшасы бактерияның ДНҚ-полимераза-2 ферменті ДНҚ тізбегінің бұзылған бөліктерін қалпына келтіріп, «жөндеу» қызметін атқарады. Тағы да айта кететін нәрсе - ДНҚ-полимераза-1 матрица ретінде ДНҚ-ның бір тізбекті бөліктерін пайдаланады, ал ДНҚ-полимераза-3 арасында қысқа бір тізбекті қатарлар болатын ДНҚ-ның қос тізбегін пайдаланады.
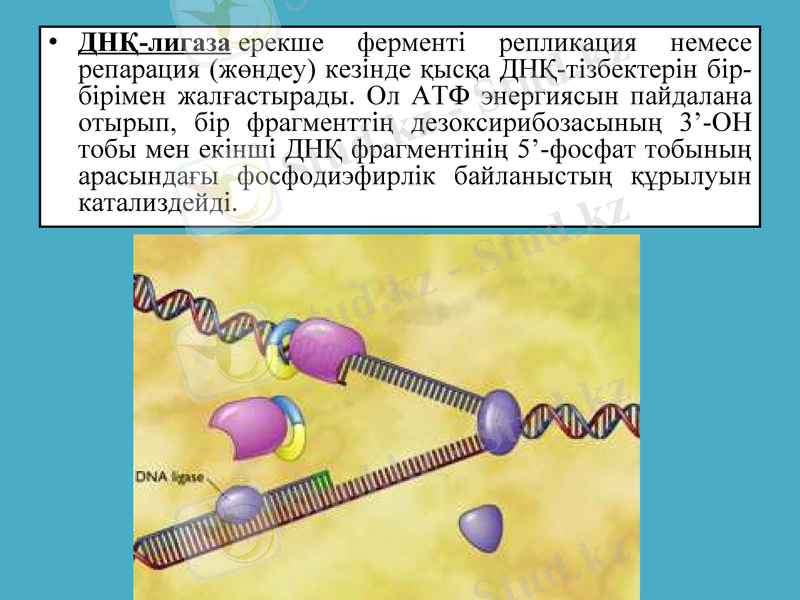
ДНҚ-лигаза ерекше ферменті репликация немесе репарация (жөндеу) кезінде қысқа ДНҚ-тізбектерін бір-бірімен жалғастырады. Ол АТФ энергиясын пайдалана отырып, бір фрагменттің дезоксирибозасының 3’-ОН тобы мен екінші ДНҚ фрагментінің 5’-фосфат тобының арасындағы фосфодиэфирлік байланыстың құрылуын катализдейді.

Прокариот ДНҚ-ның репликациясы


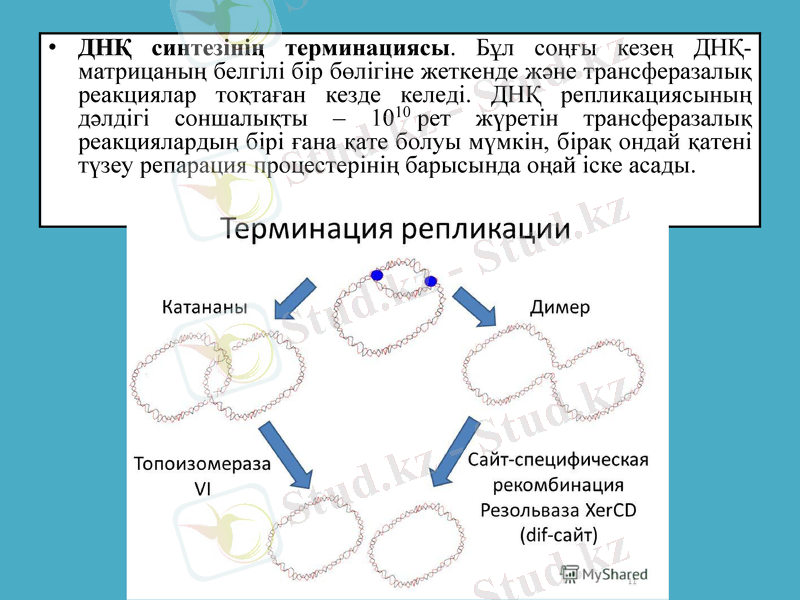
ДНҚ синтезінің терминациясы. Бұл соңғы кезең ДНҚ-матрицаның белгілі бір бөлігіне жеткенде және трансферазалық реакциялар тоқтаған кезде келеді. ДНҚ репликациясының дәлдігі соншалықты - 1010 рет жүретін трансферазалық реакциялардың бірі ғана қате болуы мүмкін, бірақ ондай қатені түзеу репарация процестерінің барысында оңай іске асады.

Эуариоттық ДНҚ-полимеразалар
ДНҚ-полимераза α (альфа) цитоплазмалық немесе үлкен полимераза деп та аталады, ол ядрода және цитоплазмада кездеседі.
ДНҚ-полимераза β (бета) - нуклеазалық полимераза немесе кіші полимераза, тек омыртқалылырға тән фермент.
ДНҚ-полимераза γ(гамма) митохондриялық полимераза.
ДНҚ- полимераза δ (дельта) - сүқоректілердің жасушасынан табылған және PCNA - белогына тәуелді; ол ДНҚ синтезінің жылдамдығы мен қарқындылығын қамтамасыз етеді.
ДНҚ-полимераза ε (епсилон) - қызметі бойынша дельта ДНҚ - полимеразаға ұқсас, ашытқы саңырауқұлақтары мен сүтқоректілердің жаушаларында кездеседі.
ДНҚ молекуласындағы репликация басталатын нүктеде А-Т жұптары көбірек, арнайы нуклеотиттер жүйесі орналасқан болады.
репликация процесі ДНҚ молекуласындағы осы арнайы нуклеотидтержүйесі мен оларды танитын белоктар (прокариоттарда - ДНҚ - А немесе А- белок) арасында байланыстың түзілуімен басталады.

Эукариоттық клеткалардағы ДНҚ репликациясының ерекшеліктері

Эукариоттардағы ДНҚ репликациясы
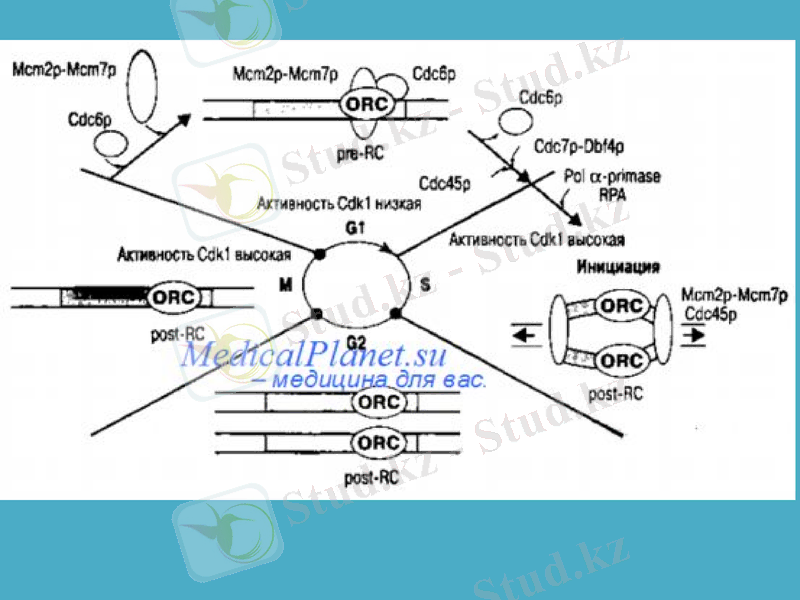
Репликация басталғанға дейін, эукариоттарда алдымен ашық ДНҚ кешені қалыптасып, іле-шала ДНҚ тізбектері тарқатылады. Бұл пресинтетикалық кезең, оның ұзақтығы 8-10 минут. Мұнда тазартылған тек үш белок: Т-антиген, RF-A және топоизомераза І мен ІІ қатысады. Т-антиген көптеген суббірліктерден тұратын кешен құрап, ДНҚ-ның қосарланған тізбектерін локальді тарқатуға жағдай жасайды.

ДНҚ жіпшелерінің одан әрі тарқатылуы RF-A фактордың және топоизомераза ферментінің, ДНҚ-геликаза-Т-ago компоненттері арқылы ДНҚ-ға жалғануына байланысты жүреді. Топоизомераза ДНҚ жіпшелерінің тарқатылуына мүмкіндік жасайды. RF-A және SSB белоктары тарқатылған тізбектерді бір-біріне байланыстырмай ұстап тұрады. Содан соң праймаза ДНҚ-полимераза α-мен әрекеттесіп РНҚ-қоздырғыштың синтезін бастайды. РНҚ-қоздырғыш, яғни РНҚ-праймер синтезделгеннен кейін 5’ - 3’бағытында Оказаки франменттері синтезделе батсайды (ДНҚ-полимераза α-ң қатысуымен) . Репликация факторы Cor RF-C циклинмен байланысып, ДНҚ-полимераза α-ны ДНҚ-полимераза δ-ға ауысуын іске асырады. ДНҚ-полимераза δ ДНҚ-ның бастаушы тізбегінің үздіксіз синтезін қамтамасыз етеді. Соңында, артта қалған және бастаушы тізбектердегі РНҚ-праймерлер босап, бөлініп кетеді. Олардың бос қалған орындары прокариоттық ДНҚ-ғы сияқты комплементарлы нуклеотидтермен толтырылады.
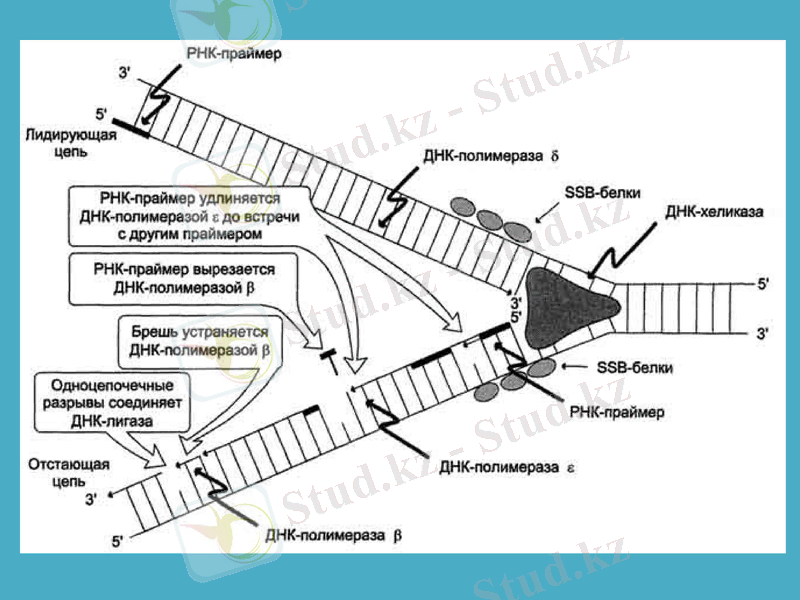

Репликацияның терминация Репликация геннің терминациялық сайттар (ter-sites) деп аталатын бірнеше арнайы нуклеотидтер жүйесінен тұратын учаскесінде. Бұл сайттар қысқа, шамамен 23 жұп нуклеотидтен құралған жүйелер. Репликация терминациясы tus генімен бақыланады. Осы ген анықтайтын tus белогы терминациялық сайтты танып, оынмен байланысып, репликациялық айырдың ары қарай жылжуына кедергі жасайды.
ДНҚ денатурациясы және ренатурациясы
Қос спиральді тұрақтандыратын сутектік байланыстар және кеңістік аралық әрекеттесулер жеткілікті әлсіз болады, және, салыстырмалы түрде көп емес әрекеттесулерде тізбектердің үзілуі жүреді, бұл процесс - денатурация немесе балқу процесі деп аталады

- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz