Теломер аймағының молекулалық механизмі: құрылымы, қызметі және теломеразаның рөлі


Теломерлі аймақтың молекулалық механизмі
Орындағандар: Осмонали Б. Б., Хамза А., Кисимова Г., Дузелова З., Әбілдашева А., Абдығапарова Г., Бейбітова Н., Бекболатова Г., Жалел М., Қуан Л., Ниятхан А., Сейтжан А., Оралова С.
Қабылдаған:Жунусбаева Ж. К.

ДҢҚ молекуласының толық , яғни теломерлік бөлімдерінің , алғаш рет 1971ж. А. М. Оловник айтқан болатын.
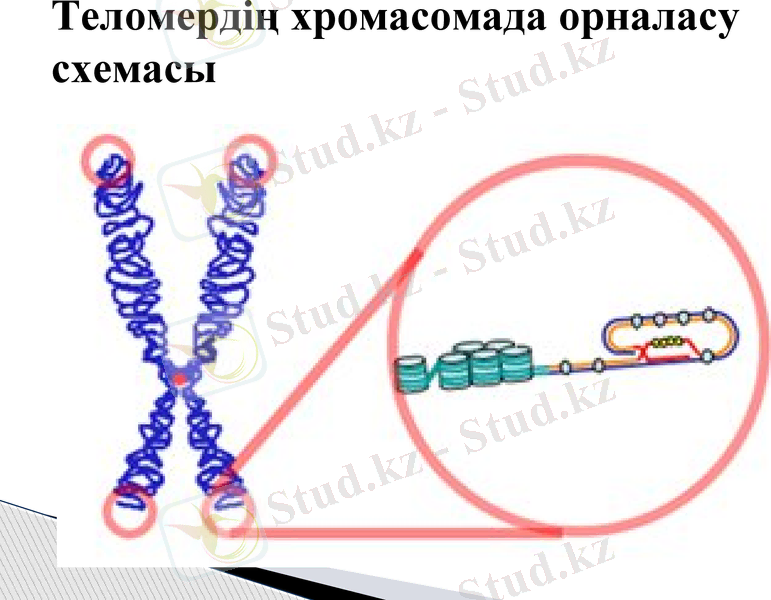
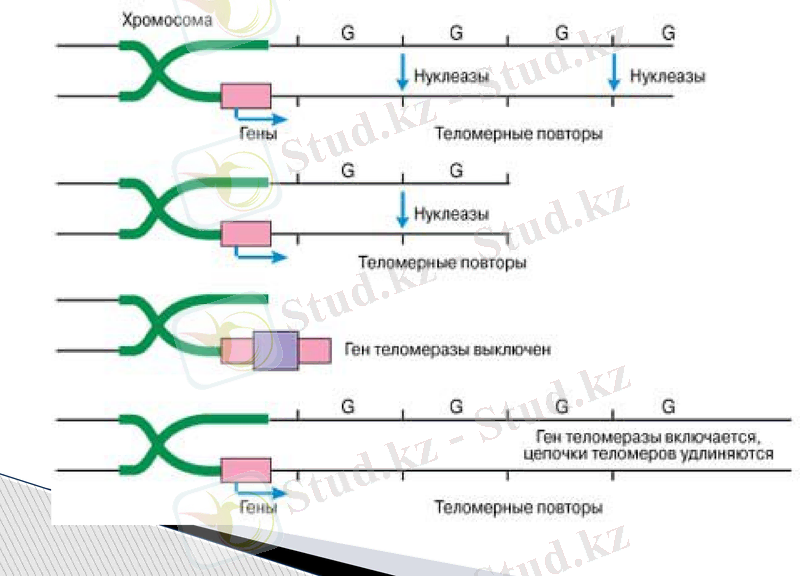
Теломердің хромасомада орналасу схемасы

Мұның мәні мынада: жоғарыда сипатталған ДҢҚ полимеразалық жүйе аналық ДҢҚ молекуласының жіпшелерінің 3 ұшын толық репликацияламайды, яғни жаңадан синтезделген ДҢҚ тізбектері 5´ ұшы жағынан қысқа болады. Себебі әрбір жаңа ДҢҚ тізбегі қысқа «РҢҚ - ұйытқыдан» (праймер) басталады. Кейін ол ерекше нуклеазалар арқылы алынып тасталады, бірақ босаған учаске толтыра алмайды, себебі ДҢҚ полимеразалар өз бетінше ДҢҚ синтезін бастай алмайды, ол тек полинкулеотидті 3´ ұшынан ұзартады. Бұл жерде ондай учаске жоқ, сондықтан жаңа тізбек матрицадан қысқа болады.
ДҢҚ молекуласының мұндай ұшын үшкір ұшы немесе оверхенга деп аталады.
ДҢҚ- ның үшкір ұшы тұрақсыз болады, себебі экзонкулезалар ұзын ұшындағы артық нуклеотидтері бір-бірлеп алып тастап, ДҢҚұшын тұйықтайды.
Қалай болғанда да, егер жасушада теломераза болмаса, оныңәрбір бөлінуінен кейін хромосома қысқарып отырады.

Әрбір репликацияда ДҢҚ молекуласы «РҢҚ-ұйытқы» ұзындығына сәйкес 10-15 нуклеотидке қысқаруы тиіс болғанымен, шындығында 50-65 нуклеотид жұбына қысқарады. Бұл ДҢҚ- полимеразалық кешенніңқасиетіне байланысты болады.
Адамның ядролық ДҢҚ-ның 1 молекуласының орташа ұзындығы 120 миллион нуклеотид жұптарына тең десек, жасушаныңәрбір бөлінуінде теломераза белсендігінсіз ДҢҚ молекуласы 0, 5% -ға қысқарады екен. Бұл әрине өте аз. Бірақ, табиғатта теломераза ұзындығын қалпына келтіріп отыратын тетіктер болмаса түбінде хромосомалар жойылып кеткен болар еді. Тек сондықтан ғана хромосомалар теломерлерінің толық и репликацияланбау проблемасының биологиялық маңызы орасан зор. Сонымен қатар, бұл құбылыс ағзалардың қартаю, канцерогенез проблемаларымен де тығыз байланысты.

Теломерлер құрылымы
ДНҚ-ның теломерлі тізбегінің, жоғарғы ұйымдастыру сатысы спецификалық белоктар есебінен жүреді.
Бұл белоктардың қарапайым гистондардан аырмашылығы олар нуклеосомды глобулаларды түзбейді. Қысқа теломерлерде нуклеосомды құрылым табылмаған. Ал тышқанның ұзын теломерлері нуклеосомды ұйымға ие.
Теломерлі белоктардың ең белгілілері - RAP 1 белогі (ашытқыларда) және оның белок аналогы TRF 1 (сүтқоректілерде) . Осы белоктардың арқасында теломерлер тығыз қалыпқа ие, яғни гетерохроматин фракциясына қатысты.
S-фазада белок TRF 1 қандайда бір сигналға жауап ретінде теломерадан диссоциацияланады-оның ұзаруы басталады
Теломерлі белоктардың көмегімен теломерлер ядролы матрикстің компоненттеріне бекітіледі, сонымен қатар, ядролы ламиналаларға да бекітілуі мумкін (ішкі ядролы мембранамен байланысқан пластинкаға) .
ДНҚ теломерасы бірнеше жіп түзеді, матриксте фиксирленген; теломерлер қысқарған сайын «жапырақшалар» саны азаяды.
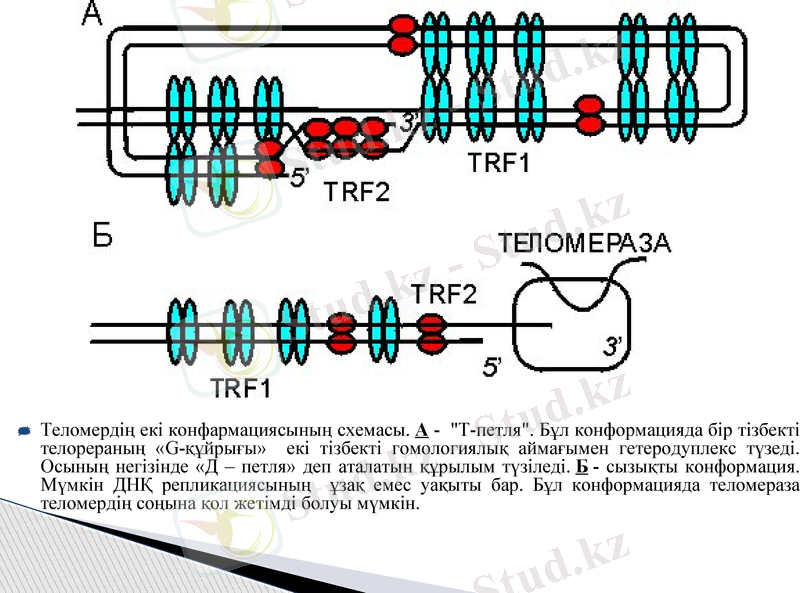
Теломердің екі конфармациясының схемасы. А - "Т-петля". Бұл конформацияда бір тізбекті телорераның «G-құйрығы» екі тізбекті гомологиялық аймағымен гетеродуплекс түзеді. Осының негізінде «Д - петля» деп аталатын құрылым түзіледі. Б - сызықты конформация. Мүмкін ДНҚ репликациясының ұзақ емес уақыты бар. Бұл конформацияда теломераза теломердің соңына қол жетімді болуы мүмкін.
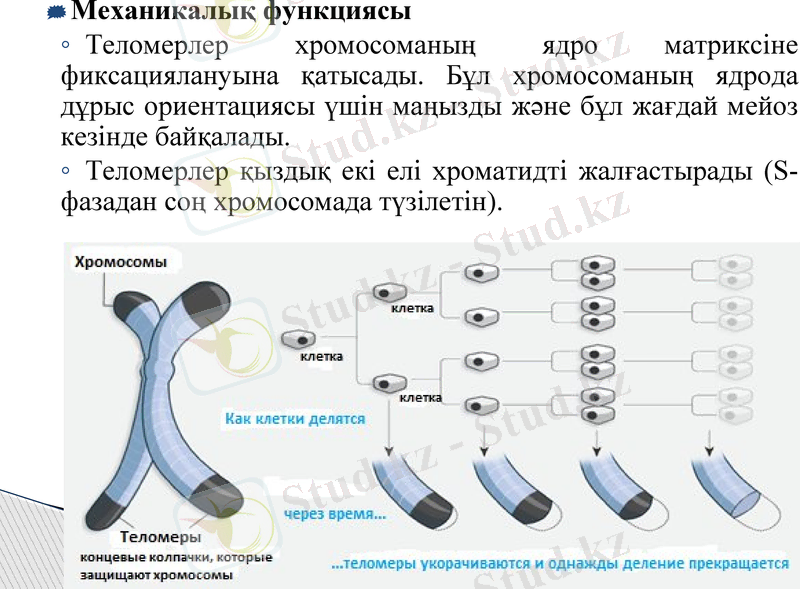
Механикалық функциясы
Теломерлер хромосоманың ядро матриксіне фиксациялануына қатысады. Бұл хромосоманың ядрода дұрыс ориентациясы үшін маңызды және бұл жағдай мейоз кезінде байқалады.
Теломерлер қыздық екі елі хроматидті жалғастырады (S-фазадан соң хромосомада түзілетін) .
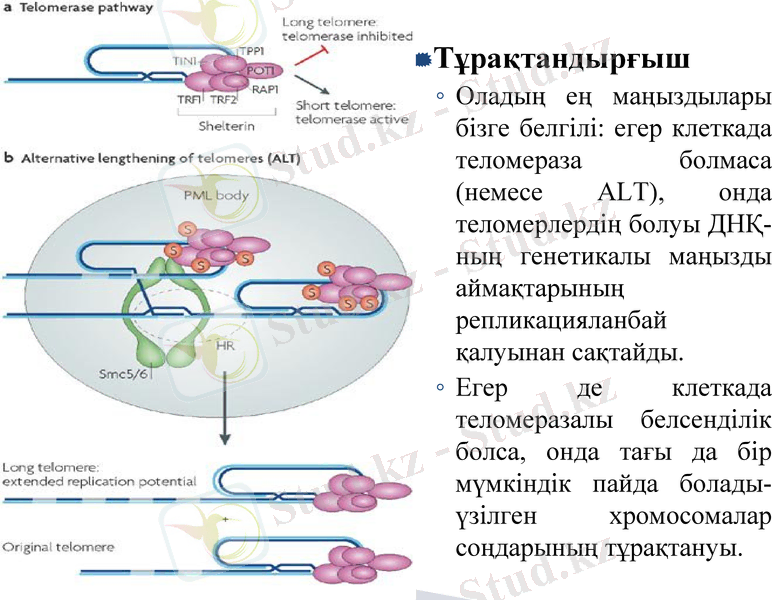
Тұрақтандырғыш
Оладың ең маңыздылары бізге белгілі: егер клеткада теломераза болмаса (немесе ALT), онда теломерлердің болуы ДНҚ-ның генетикалы маңызды аймақтарының репликацияланбай қалуынан сақтайды.
Егер де клеткада теломеразалы белсенділік болса, онда тағы да бір мүмкіндік пайда болады-үзілген хромосомалар соңдарының тұрақтануы.

Ғылыми деректер бойынша хромосома ұштарында генетикалық ақпарат болмайтын көптеген арнайы гексонуклеотид (6 нуклеотидтен тұратын) бірізділіктер қайталанып орналасқан.
ДНК - ның теломерлік бөлімдерінде осындай мыңдаған гексонуклеотидтер қайталанады. Олардың жалпы ұзындығы адам эмбрионы жасушаларында 10-15 мың нуклеотид жұптарына тең. Сонымен, хромосоманың екі теломерлік ұшы, адамның ядролық ДНК молекуласының ұзындығының 0, 02% құрайды.
Теломерлік қайталанулардан ешқандай генетикалық ақпарат болмайды, сондықтан да теломерасыз олардың біршама бөлігі түсіп қалған күннің өзінде де геном бірқалыпты қызмет ете береді. Теломердің негізгі қызметінің өзі де осы болса керек, яғни олар геномның маңызды бөлімін толық репликациялаудан қорғап, буферлік қызмет атқарады.
Әйтсе де, теломеразадан біржола бас тартуға болмайды, себебі жасушаның бөліну үдерісінде күндердің күнінде ДНК - ның теломерлік участкілері қысқарып жойылуы мүмкін. Сонымен қатар теломерлік учаскілер ереше, арнай қызметтер де атқарады. Сондықтан ол белгілі бір шекке дейін ғана қысқарады.
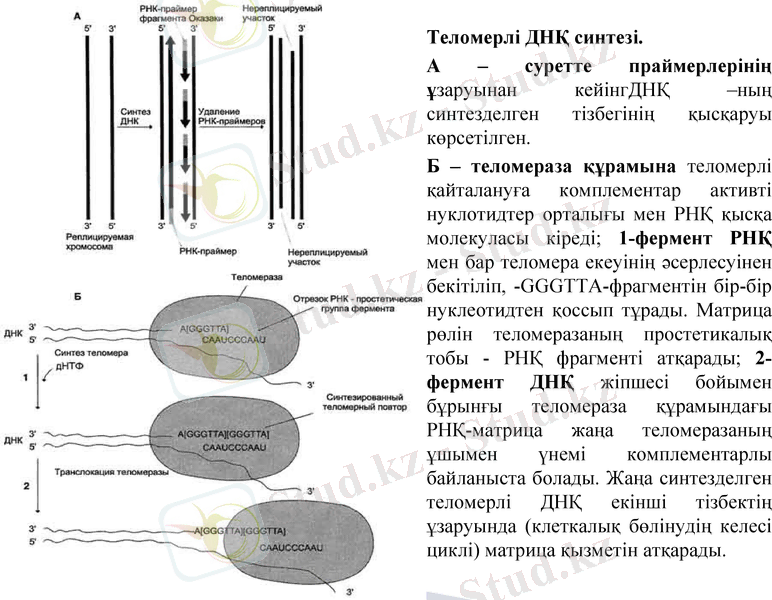
Теломерлі ДНҚ синтезі.
А - суретте праймерлерінің ұзаруынан кейінгДНҚ -ның синтезделген тізбегінің қысқаруы көрсетілген.
Б - теломераза құрамына теломерлі қайталануға комплементар активті нуклотидтер орталығы мен РНҚ қысқа молекуласы кіреді; 1-фермент РНҚ мен бар теломера екеуінің әсерлесуінен бекітіліп, -GGGTTA-фрагментін бір-бір нуклеотидтен қоссып тұрады. Матрица рөлін теломеразаның простетикалық тобы - РНҚ фрагменті атқарады; 2-фермент ДНҚ жіпшесі бойымен бұрынғы теломераза құрамындағы РНҚ-матрица жаңа теломеразаның ұшымен үнемі комплементарлы байланыста болады. Жаңа синтезделген теломерлі ДНҚ екінші тізбектің ұзаруында (клеткалық бөлінудің келесі циклі) матрица қызметін атқарады.

РНҚ-ның көмегімен толық құрылмаған жаңа ДНҚ тізбегінің 3' соңына фермент комплементарлы болып орнығады. Теломераза комплементарлық принципі бойынша ДНҚ тізбегінің 3' соңын бір гексануклеотид -GGGTTA- қа ұзартады. Синтез үнемі 5'- тан 3'-ұшына қарай. Сосын теломераза ДНҚ тізбегі бойымен бір теломерге жылжиды және жаңа -GGGTTA- фрагментінің синтезін бастап кетеді.
Көптеген соматикалық клеткаларда теломераза активті емес, өйткені соматикалық клетка теломерлі ДНҚ секілді ұзындыққа ие, ал ол клетка өзінің және оның ұрпақтарының тіршілігіне жеткілікті. Алайда өте жылдам жаңару қасиетіне ие клеткаларда (лимфоциттер, бағаналы клеткалар, эпителий, тері эпидермисінің клеталары, т. б. ) теломеразаның активтілігі байқалады.
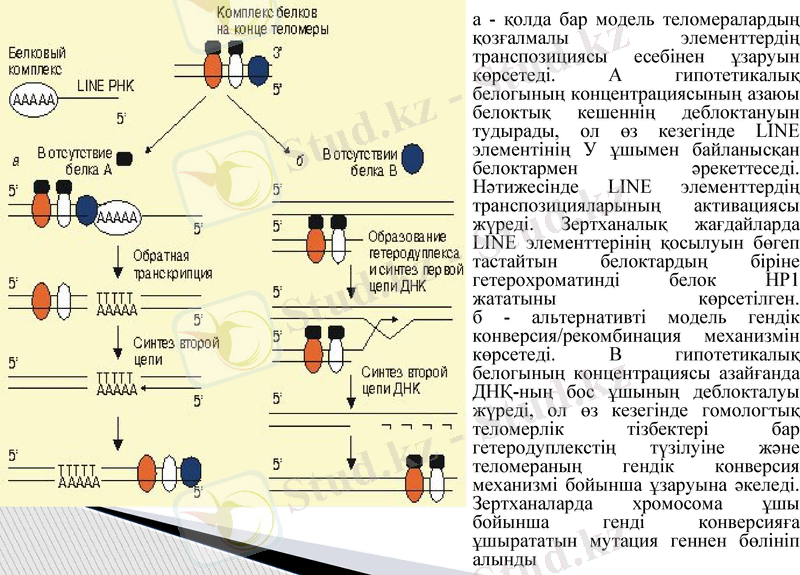
а - қолда бар модель теломералардың қозғалмалы элементтердің транспозициясы есебінен ұзаруын көрсетеді. А гипотетикалық белогының концентрациясының азаюы белоктық кешеннің деблоктануын тудырады, ол өз кезегінде LINE элементінің У ұшымен байланысқан белоктармен əрекеттеседі. Нəтижесінде LINE элементтердің транспозицияларының активациясы жүреді. Зертханалық жағдайларда LINE элементтерінің қосылуын бөгеп тастайтын белоктардың біріне гетерохроматинді белок НР1 жататыны көрсетілген. б - альтернативті модель гендік конверсия/рекомбинация механизмін көрсетеді. В гипотетикалық белогының концентрациясы азайғанда ДНҚ-ның бос ұшының деблокталуы жүреді, ол өз кезегінде гомологтық теломерлік тізбектері бар гетеродуплекстің түзілуіне жəне теломераның гендік конверсия механизмі бойынша ұзаруына əкеледі. Зертханаларда хромосома ұшы бойынша генді конверсияға ұшырататын мутация геннен бөлініп алынды

ДНҚ -ның теломерлі аймақтары сағат құралы ретінде іске қосылады - репликометр. Олар теломераза белсенділігінің жойылуынан кейінгі клетка бөлінісінің санын санайды. Әрбір бөліну теломердің 50-60 н. ж. қысқаруына алып келеді. Клетка үшін қанша рет бөлінгені емес, ал оның критикалық қысқаруының қалғаны маңызды.
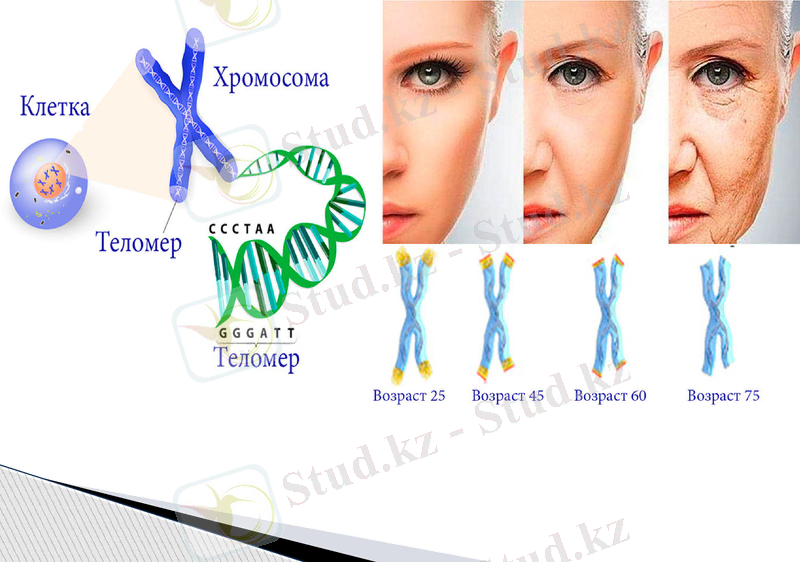
Теломердің ДНҚ-ның басқа аймақтарынан айырмашылығы: теломерлер ақуыз молекулаларын кодтамайды. Бір жағынан қарағанда бұл «геномның мағынасыз» аймақтары. Теломерлер клетканың әрбір бөлінуінің нәтижесінде қысқарып отырады. Яғни теломерлі аймақтардың ұзындығы клетканың «жасын» анықтайды - теломер «құйрығы» неғұрлым қысқа болса, соғұрлым оның жасы үлкен болады.
Теломер «құйрығы» неғұрлым қысқа болса, соғұрлым оның жасы үлкен болады деген болжамды 15 жылдан кейін тәжірибе жүзінде ағылшын ғалымы Говард Гук дәлелдеді. Бірақ ересек ағзаның жүйке және бұлшықет клеткалары бөлінбейді, ал теломерлі аймақтары қысқармайды, олар тек қана қартайып өледі.
Теломерлер клеткалық бөлінудің счетчигі ретінде қызмет атқарады: олар қаншалықты қысқа болса, алғашқы клетка соншалықты көп бөлінген.
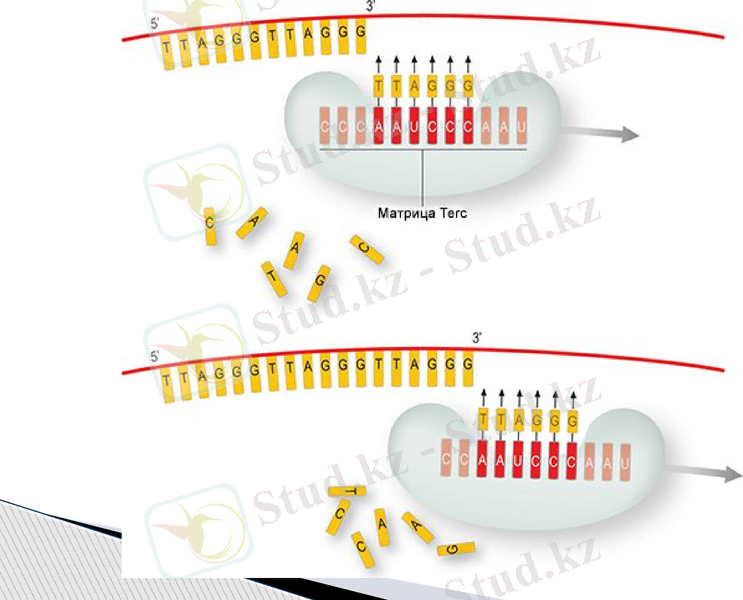

Қарапайым (сомалық) клеткаларда теломераза жұмыс істемейді. Сондықтан клетканың әрбір бөлінуінен кейін теломерлер қысқарып отырады, нәтижесінде оның өліміне әкеп соқтырады.
1997 жылы Колорадо Университетінің ғалымдары теломераза генін алды.
Ал 1998 жылы Далластағы Техас университетінің Оңтүстік-Батыс медициналық орталығының зерттеушілері адамның тері клеткаларына, көру және қантамырлардың эпителилеріне теломераза генін енгізді.
Мұндай генетикалық модефицирленген клеткаларда теломераза ферменті жұмыс қалпында болады - ДНҚ-ның соңына нуклеотидті тізбектерді тігіп отыруының арқасында клетка бөлінісінің нәтижесінде теломерлер ұзындығы өзгеріске ұшыраған жоқ. Осы әдіс арқылы ғалымдар қарапайым клеткалар өмірін бір жарым есеге ұзартуға қол жеткізді.
Сонымен қатар бұл фермент - қатерлі ісік клеткаларының туылуының басты факторларының бірі. Ісік клеткаларының жойылмауы теломераза ферментінің жұмыс істеуінің арқасында. Өлмейтін ағза теориялық тұрғыдан көп жыл жасауы мүмкін, бірақ ол сөзсіз ісік ауруынан өледі.

- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz