Гаплоидтық технология: микроспоралар мен аналық гаметофиттің in vitro дамуы және селекциядағы қолданылуы


Тақырыбы: 1. Гаплоидтық технология. 2. Микроспоралардың in vitro жағдайында дамуы. 3. Аналық гаметофиттің in vitro жағдайында өсіру.
Тексерген:

Жоспар:
Кіріспе
Гаплоидтық технология
Микроспоралардың in vitro жағдайында дамуы
Аналық гаметофиттің in vitro жағдайында өсіру
Қорытынды
Пайдаланылған әдебиеттер

Гаплоидтық технология

Гаплоидтық организмнің сомалық жасушаларында сыңар хромосомалар жиынтығы (n) болады, яғни толық жиынтықтың (2n) тең жартысы. Гаплоидтарды дағдылы селекция әдістерімен шығару (түрішілік және түраралық тозаңдану, рентген сәулесін түсіру және басқа стресс факторлармен ықпал ету) оңай емес және көп уақытты талап етеді. Ал аталық және аналық гаметофиттерді in vitro жағдайында өсіріп гаплоидтарды тез шығарып, селекция процесін жеңілдетуге болады. Бұл әдістер апомиксис процесіне негізделген. Апомиксис - организмдердің жыныссыз жолмен көбеюі.
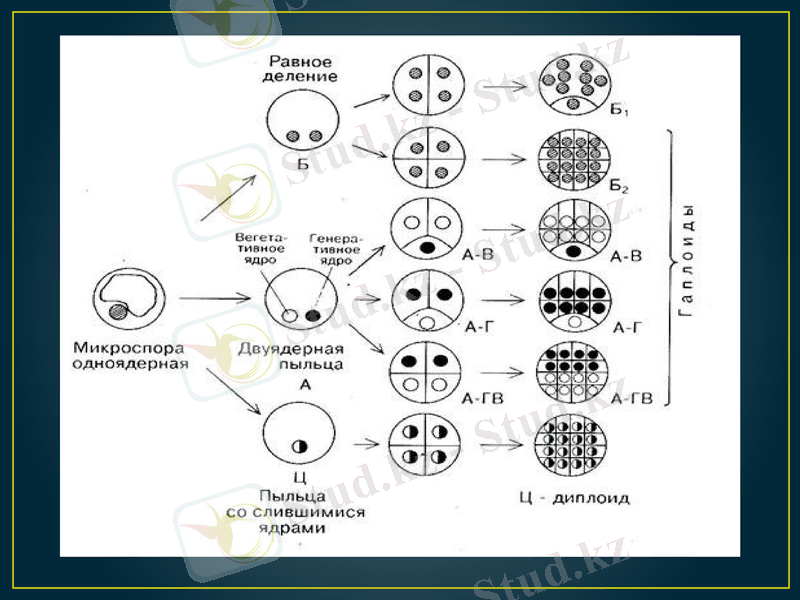
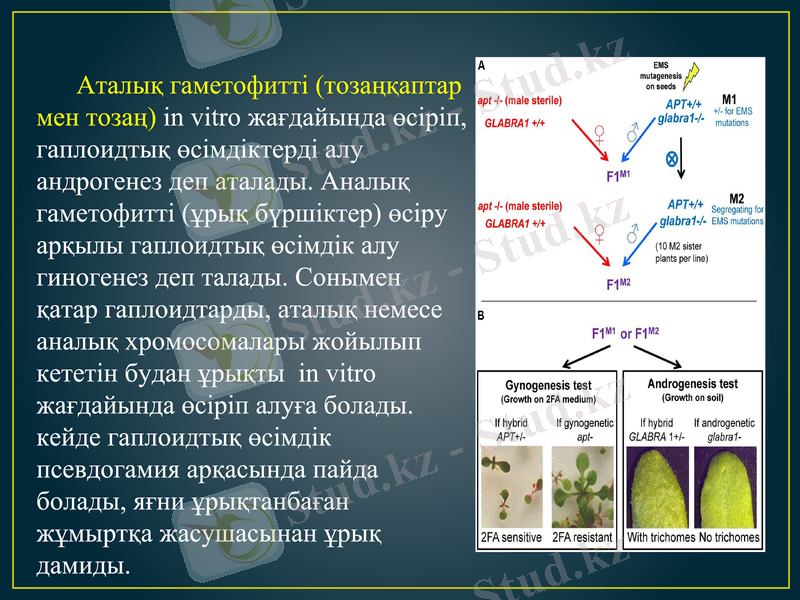
Аталық гаметофитті (тозаңқаптар мен тозаң) in vitro жағдайында өсіріп, гаплоидтық өсімдіктерді алу андрогенез деп аталады. Аналық гаметофитті (ұрық бүршіктер) өсіру арқылы гаплоидтық өсімдік алу гиногенез деп талады. Сонымен қатар гаплоидтарды, аталық немесе аналық хромосомалары жойылып кететін будан ұрықты in vitro жағдайында өсіріп алуға болады. кейде гаплоидтық өсімдік псевдогамия арқасында пайда болады, яғни ұрықтанбаған жұмыртқа жасушасынан ұрық дамиды.

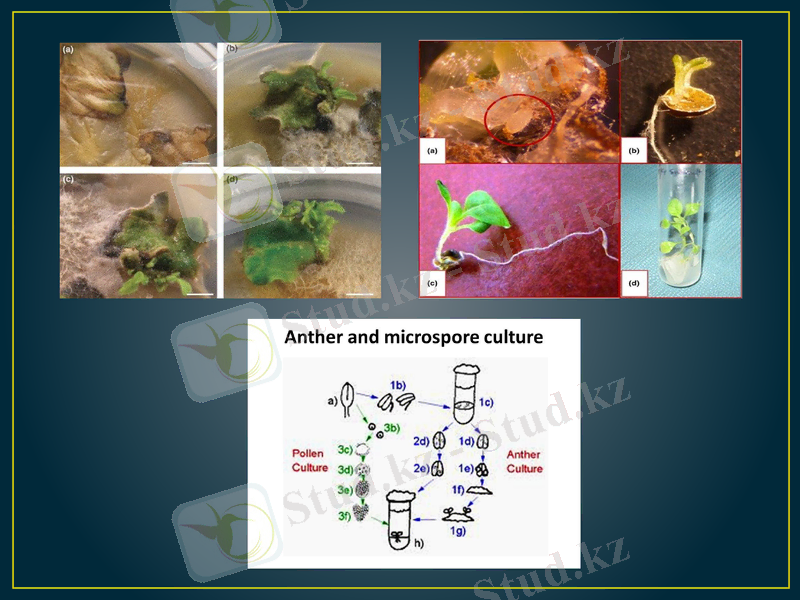
Тозаңқаптар мен тозаңдарды өсіріп гаплоидтарды алу. Бірінші рет сасық меңдуананың тозаңқаптарын in vitro жағдайында өсіріп, Индияда С. Гуха мен С. Махешвари 1964 жылы гаплоидтық өсімдіктерді алады. Содан кейін осы тәжірибені француз ғалымы К. Нич 1967 жылы темеккінің тозаңқаптарын өсіріп қайталады. Содан бері осы әдіспен гаплоидтар 200-ден астам өсімдік түрлерінен алынды, соның ішінде: бидай, арпа, қара бидай, күріш, картоп, рапс, т. б. ауыл шаруашылық дақылдары.
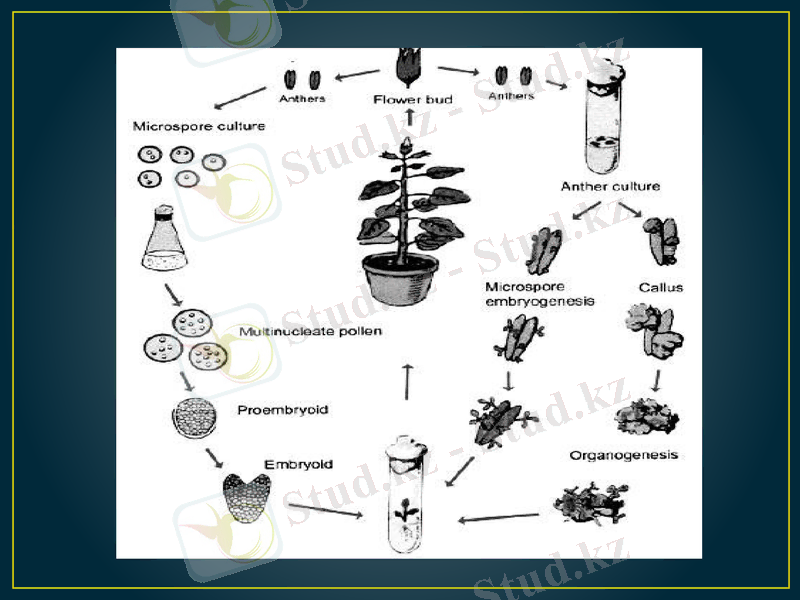

In vitro жағдайында аталық гаметофиттен гаплоидтық спорофиттің шығуы өте күрделі, әлі егжей-тегжейі анықталмаған процесс. Микроспоролардан калус немесе эмбриоидтар қалыптасып, кейін олардан гаплоидтық регенерант өсімдіктері шығады. Ол үшін микроспоралар in vitro жағдайында дамудың өте күрделі процесінен өтеді. Ал бұл процестің түрткі болар себепкерлері мен реттеушілері әлі белгісіз. Табиғи жағдайда in vivo микроспоралар (гаметофиттік) жолмен дамиды: 1) мейоздың нәтижесінде тозаңның бастапқы жасушасынан төрт (тетрада) микроспора пайда болады. ; 2) тетраданың қалың қабығынан микроспоралар босап шығады; 3) микроспора одан әрі даму жолында екі рет митоздан өтіп, соңында жетілген тозаң түйіріне айналады. Ал in vitro жағдайында микроспора әдеттегі гаметофиттік даму жүйесінің кез келген фазасынан ауытқып кетіп, сапрофиттік даму жолына түсуі мүмкін.

Микроспоралардың in vitro жағдайында дамуы

Микроспоралардың in vitro жағдайында дамуы бірнеше жолмен қамтамасыз етілуі мүмкін. Бірлі-жарымы гаметофиттік даму жолынан ауысып эмбриоидты түзеді. Басқалары дедифференцияланып, каллусқа айналады. Тағы біреулері микроспорогенез бен гаметогенез жолын жалғастырады, яғни пісіп жетілген тозаңға айналады. Ал тағы бір тобы біртіндеп ыдырап құриды. Н. Сандерленд сасық сеңдуананың микроспораларын in vitro өсіріп, олардың бастапқы кездегі даму жолдарын зерттеп, табиғаттағы in vivo даму жолынан айырмашылығын көрсеткен.

Микроспора көлемі бірдей емес екі жасушаға бөлініп, вегетативтік және генеративтік жасушаларды түзеді. Бұ табиғи жолға ұқсас. Генеративтік жасуша бірінші митоздан кейін кері кетеді. Вегетативтік жасуша көп бөлініп одан әрі зиготалық даму жолына түседі. Бұндай даму жолы Nicotiana tabacum, Datura metel, Hordeum vulgare, Triticum aestivum т. б. өсімдіктерде байқалған. Кей кезде тек қана генеративтік жасуша бөліне бастайды, немесе екеуі де (вегетативтік пен генеративтік жасушалары) бірдей бөлініп, спорофитті түзеді. Микроспораның атап өткен бөліну бағыттары көпжасушалық гаплоидтық құрылымдардың (глобула, эмбриоид) пайда болуына себеп болады. олардан гаплоидтық регенерант өсімдіктері шығады.
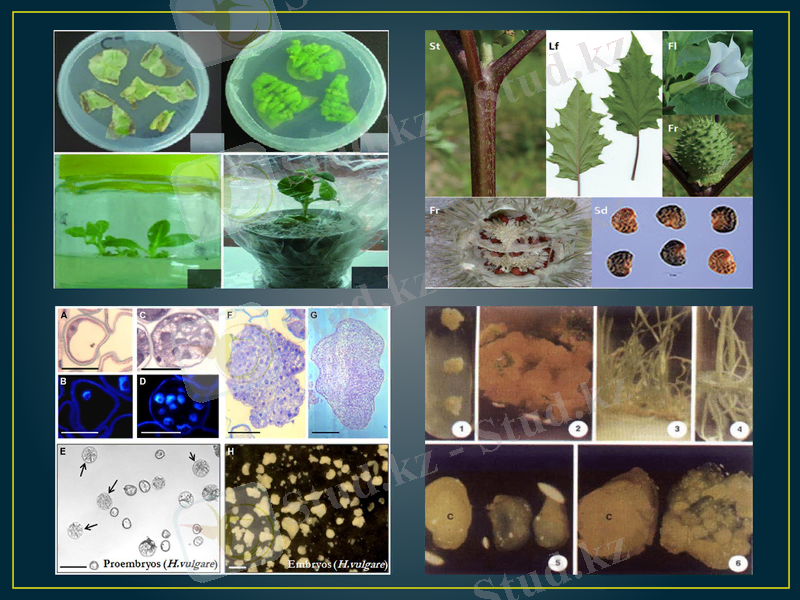

В. Ананд қызметтестерімен темекінің (Nicotiana tabacum) тозаңқаптарын өсіргенде, эмбриоидтардың үш түрі пайда болғанын байқаған. Олар вегетативтік жасушаның, генеративтік жасушаның және ол екеуінің де бірге бөлінуінен түзілген. Осы эмбриоидтардан гаплоидтық өсімдіктер алып, оларды топыраққа көшіріп, гүлденуге дейін өсірген. Олардың жапырақтары мен гүлдерінің морфологиялық құрылысын зерттегенде, бұл өсімдіктер де бір-бірінен айырмашылықтары бар үш типке бөлінген. Тозаңқаптар мен микроспораларлы өсіргенде, каллустың екі типі пайда болатыны көрсетілген: біріншісі - әр түрлі гетерогендік жасушаларынан тұрған, екіншісінің құрамында меристемалық жасушалар ошағы болған. Регенерация прцесі екінші каллуста өте жеңіл өткен, себебі меристемалық аймақтардан өркендер тез өсіп шыққан.

Аналық гаметофиттің in vitro жағдайында өсіру
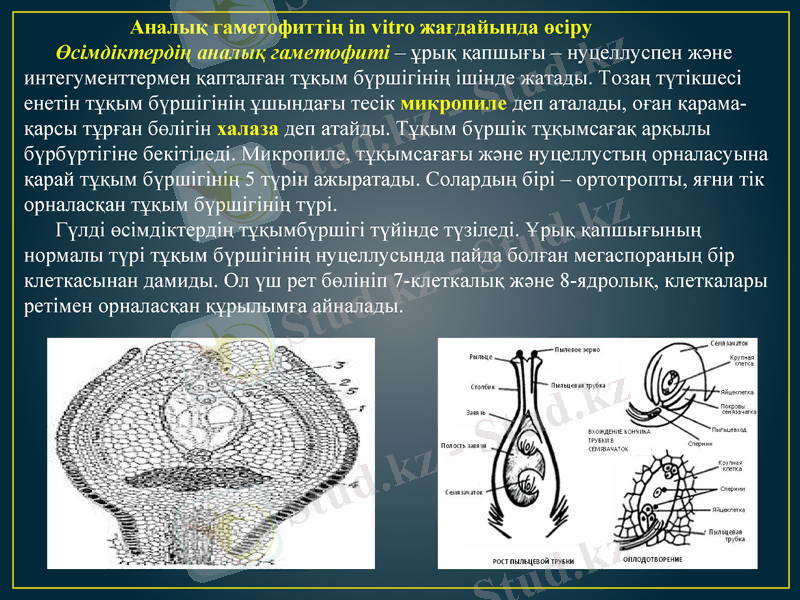
Аналық гаметофиттің in vitro жағдайында өсіру Өсімдіктердің аналық гаметофиті - ұрық қапшығы - нуцеллуспен және интегументтермен қапталған тұқым бүршігінің ішінде жатады. Тозаң түтікшесі енетін тұқым бүршігінің ұшындағы тесік микропиле деп аталады, оған қарама-қарсы тұрған бөлігін халаза деп атайды. Тұқым бүршік тұқымсағақ арқылы бүрбүртігіне бекітіледі. Микропиле, тұқымсағағы және нуцеллустың орналасуына қарай тұқым бүршігінің 5 түрін ажыратады. Солардың бірі - ортотропты, яғни тік орналасқан тұқым бүршігінің түрі. Гүлді өсімдіктердің тұқымбүршігі түйінде түзіледі. Ұрық қапшығының нормалы түрі тұқым бүршігінің нуцеллусында пайда болған мегаспораның бір клеткасынан дамиды. Ол үш рет бөлініп 7-клеткалық және 8-ядролық, клеткалары ретімен орналасқан құрылымға айналады.

- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz