Пикорнавирустар, рабдовирустар, реовирустар, ретровирустар, тогавирустар және флавивирустар: құрылымы, таксономиясы және репродукциясы


СӨЖ Тақырыбы: Пикорнавирустар. Рабдовирустар. Реовирустар. Ретровирустар. Тогавирустар. Флавивирустар. Тексерген: Бейсембаева Г. Ш. Тұрарбек Т. М. Орындаған: Мырзагалиева А. Е. Топ: БТ-507 Семей 2016 жыл
Қазақстан Республикасының Білім және Ғылым Министрлігі Шәкәрім атындағы Семей мемлекеттік университеті «Стандарттау және Биотехнология» кафедрасы

Жоспар:
Пикорнавирустар.
Рабдовирустар.
Реовирустар.
Ретровирустар.
Тогавирустар.
Флавивирустар.

Пикорнавирустар
Пикорнавирустар - құрамында РНҚ бар, липопротеид қабығы жоқ вирус тұкымдастығы, РНҚ м. с. 25-28. Жалғыз тізбекті, ауру туғызғыш. Вириондарының диаметрі 25-40 нм. Цитоплазмада көбейеді. 4 туыстыққа бөлінеді: 1) Энтеровирустар адамдарды зақымдайды, бұларға ішек вирустарынан басқа полиомиелит және А гепатиті вирустары енеді; 2) Риновирустар - адамда жәие ірі қара малдарда тыныс жолдары ауруларын туғызады; 3) Кардиовирустар - кеміргіштерден бөлініп алынған; 4) Афтовирустарға маңызы зор мал аурулары қоздырғыштарының бірі - аусыл вирусы жатады. Пикорнавирустар тасымалдаушылар көмегінсіз жуғады.
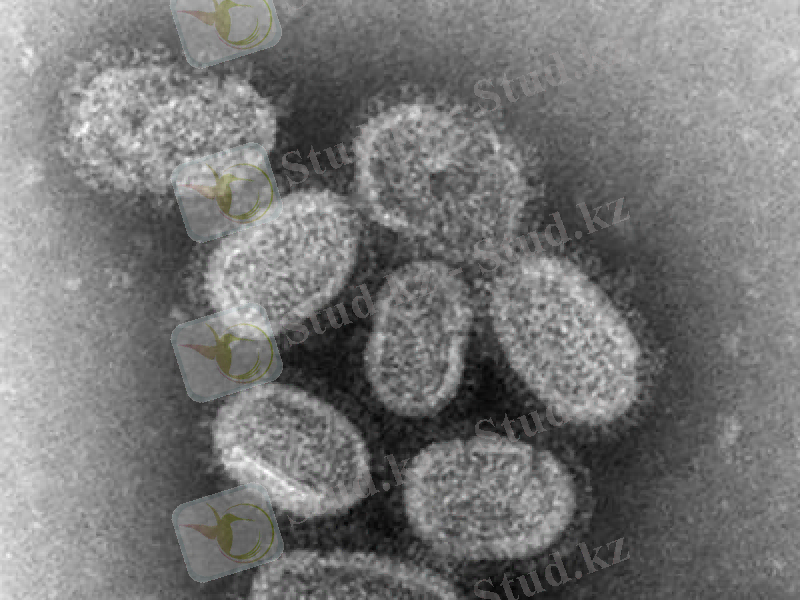

Таксономиясы
Тұқымдастығы: Picornaviridae (230-дан астам өкілдері бар) .
Туыстастығы: Enterovirus (111 сероварианттары бар) ;
Rhinovirus (105 серотиптері бар) ;
Aphthavirus (7 серотиптері бар) ;
Hepatovirus (2 серотипі - адамдарда, 1-маймылдарда) ;
Cardiovirus (2 серотипі бар) ;
Parecovirus, Erbovirus, Kobuvirus - жаңадан ашылған туыстастықтар.
Әрбір туыстастық түрлерден, ал түрлер - серотиптерден тұрады. Туыстастықтарды бір-бірінен ажыратып - анықтау (дифференциациялау) антигендік айырмашылықтарына, ортаның рН-на сезімталдығына, адамдарға патогенділігінің айырмашылығына ж. т. б. қасиеттеріне негізделген. Энтеровирустар адамдарда нерв жүйелерінің және басқа ағзалар мен тіндерде патологиялық үрдістер қоздырады. Кардиовирустар және афтовирустар көбінесе жануарларға патогенді болып келеді. Энтеровирустар құрылымы, химиялық құрамы, физикалық және химиялық факторлардың әсеріне төзімділігі ж. б. қасиеттері бойынша бір-біріне жақын келеді.

Құрылымы
Пикорнавирустар (энтеровирустар) қарапайым құрылымды ұсақ вирустарға жатады. Диаметрі 30 нм-дей, вирион массасы 8-9 МД құрайды. Вирион икосаэдрлі капсидтен тұрады, симметриясы куб типтес. Капсид 12 бесбұрыштардан (пентамерлер) тұрады, олардың әрқайсысы өз кезегінде бірнеше ақуыздық суббірліктерден - протомерлерден құралған. Оларды VP1, VP2, VP3 және VP4 деп белгілейді. Суперкапсиді жоқ. Вирус құрамында көмірсулар және липидтар болмайды, сондықтан олар эфирдің ж. б. май еріткіштердің әсеріне сезімтал емес. VP1, VP2, VP3 - капсидтің сыртқы беткейін құрайды, VP4 - вирионның ішкі жағының құрамдасы, сондықтан сыртынан ол көрінбейді. Сыртқы қабатының ақуыздары иесі жасушасының рецепторларын танып алуда, жасушаға вирионның жабысуында және жасуша ішінде вириондық РНҚ-ң босанып шығуында рөл атқарады, және де вирустың иммуногендік қасиетін анықтайды

Геномы
VPg ақуызымен байланысқан, сегменттелмеген (+) РНҚ молекуласынан тұрады. Вириондық РНҚ капсидті алып тастаған жағдайда да өзінің инфекциондық (жұқпалық) қасиетін сақтайды.

Репродукциялануы
Вирус жасуша сыртындағы рецепторлармен өзара әрекеттеседі. Вирус геномы жасуша ішінде эндоцитоз тәсілімен немесе цитоплазмаға геномдық (+) РНҚ-ның инъекциялануы арқылы түседі. Вириондық РНҚ оң жіпшелі РНҚ болатындықтан вирус - спецификалық ақуыздарға тікелей транслирленеді, яғни ақуызды синтездеу үшін вирус геномы информациялық РНҚ (иРНҚ) ретінде пайдаланылады. Трансляциялану процесі кезінде бірегей полипептидті тізбек синтезделеді, сонан соң ол протеазаның әсерінен 4 ақуыздарға (VP1-VP4) және РНҚ-тәуелді полимеразаға ыдырайды. Бұл полимераза геномдық оң-жіпшелі РНҚ-нан теріс жіпшелі РНҚ-н (матрица) синтездейді. Оң/теріс жіпшелерден вирус геномы репликацияланады. Жаңадан синтезделген вириондық РНҚ (вРНҚ) капсидке қосылады. Капсидте құрылымдық ақуыздар жинақталады. Содан кейін жаңадан синтезделген вириондар жасушадан лизистену жолымен сыртқа шығады. Жасуша ішінде бір вирионнан 150 000-ға дейін жаңа вириондар синтезделеді. Репликациялану жасушаның цитоплазмасында жүреді және цитопатиялық әсер етумен аяқталады.

Рабдовирустар
Рабдовирустар (грекше: rhabdos - шыбық, таяқ деген мағына береді) - сүтқоректілерде, балықтарда, жәндіктерде, қарапайымдыларда және өсімдіктерде әртүрлі зақымдану туғызатын, РНҚ - құрамды вирустар, олардың 80-нен астам түрлері белгілі. Rhabdoviridae тұқымдастығы өкілдерінің табиғи иесі кең ауқымды - олар әртүрлі омыртқалы және омыртқасы.

Таксономиясы
Тұқымдастығы:Rhabdoviridae
Туыстастығы:Vesicоlovirus -везикулярлы Индиана стоматит вирусы;
Lyssavirus - құтыру вирусы;
Ephemerovirus - сиырдың біркүндік қызба вирусы;
Cytorhabdoviru-салатта сарғышнекр қоздыратын вирус;
Nucleorhabdovirus - картопта сарғыш ергежейлік қоздыратын вирус;
Novirhabdovirus - гематопоэздің жұқпалы некрозын қоздыратын вирус.

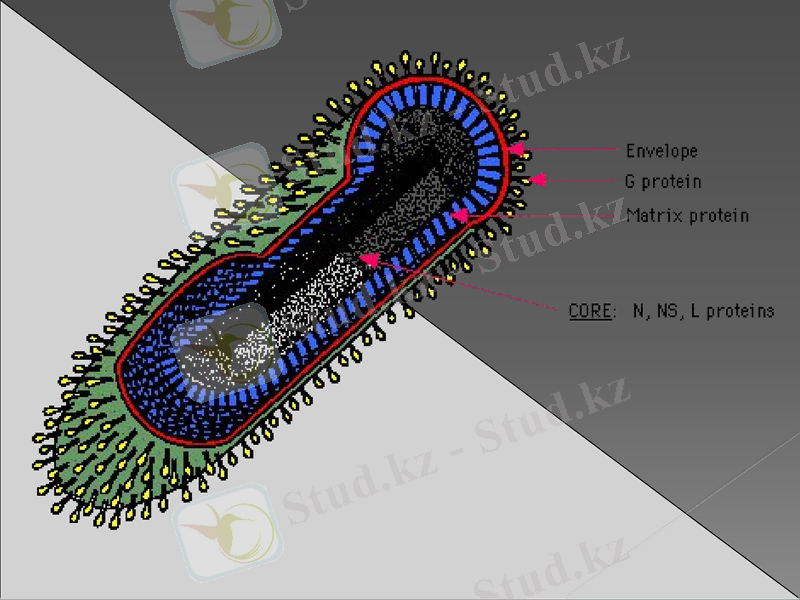
Құрылымы
Вириондары - оқ (пуля) тәрізді немесе таяқша пішінді, мөлшері 170 х 70 нм. Сыртында екі қабатты липидқұрамды қабықшасы (суперкапсид) болады, одан гликопротеинді тікенекшелер (ұзындығы 5 -10 нм, диаметрі - 3 нм) шығып тұрады. Вирионның ортасында - сыртқы қабатпен матрикстік ақуызбен (М) бөлініп тұратын, симметриясы спираль типті нуклеокапсид (РНП) орналасқан.

Репродукциялануы
Жасушаның цитоплазмасында атқарылады. Рабдовирустардың репродукциялануы (-) РНҚ - құрамды вирустардыкіне ұқсас. Рабдовирустар жасуша рецепторларына гликопротеинді тікенекшелерімен байланысады да, эндоцитоз жолымен жасуша ішіне енеді. Содан кейін вирустың қабықшасы лизистенеді, босанған РНП [РНҚ+NP-нуклеопротеин + полимераза ақуыздары [L және NS] немесе нуклеокапсид жасуша цитоплазмасына түседі. РНҚ - полимеразаның көмегімен толықсыз (неполные) оң-жіпшплі РНҚ (вирустық ақуыздардың синтезделуі үшін бес меншікті иРНҚ) және минус геномды РНҚ-ң синтезделуіне матрица болып табылатын, толықты (полные) оң-жіпшелі РНҚ синтезделеді. Рибосомалармен иРНҚ-ң трансляциялануының нәтижесінде пайда болған вирустық ақуыздар Гольджи аппаратында өзгеріске ұшырайды да, жасуша плазмолемасына, яғни қабықшасына қосылады. Геномдық (минус - жіпшелі) РНҚ -ның N, NS, L- ақуыздармен өзара әсерлесуінің нәтижесінде РНП пайда болады. Вирион құрастырылғаннан кейін бүршіктену тәсілімен жасушадан шығады.

Реовирустар
Реовирустар-тыныс алу және ішек жолдары арқылы, кейде трансмиссивті тәсілмен жұғатын, сыртқы қабаты жоқ, екі жіпшелі фрагменттелген РНҚ-құрамды вирустар тұқымдастығына жатады.
Тұқымдастығы терминін бірінші рет А. Сэбин(1959 ж. ) адамдардан бөлінген бір топ вирустарды атау үшін ұсынған. Оларды алғашқыда ЕСНО-вирустардың 10-типіне жатқызып келген болатын. Реовирус-ағылшынша respiratory enteric orphan virus-деген сөздердің бастапқы әріптерінен тұрады, қазақша -респираторлық энитериттік жетім вирус -деген мағына береді.
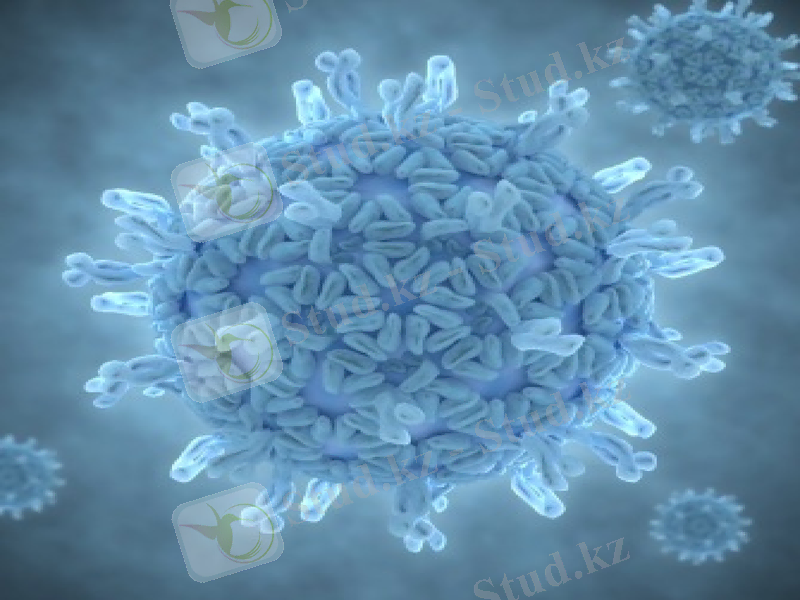

Таксономиясы
Тұқымдастығы: Reoviridae
Туыстастығы: Orthoreovirus-адамдар мен сүтқоректілердің ортореовирустары.
Orbivirus-қойлардың көгерген тіл вирусы, кемерово вирусы.
Сoltivirus-колорадалық дене қызбасының вирусы.
Rotavirus-адамдар мен маймылдардың ротовирустары және т. б.
Бұл тұқымдастықтың көптеген өкілдері табиғатта кең таралған. Олар сүтқоректілермен қатар құстарды, жәндіктерді және өсімдіктерді де зақымдайды. Адамдарға патогенді реовирустар 3 туыстастыққа жатады-меншікті реовирустар, ротавирустар, орбивирустар. Олар вирионының құрылымы бойынша бір тұқымдастыққа топтастырылған. Вирондары екі жіпшелі фрагменттелген РНҚ-нан. екі қабатты (сыртқы және ішкі) капсидтен және РНҚ-мен байланысқан вирус-спецификалық транскриптазадан тұрады.

Ретровирустар
Ретровирустар- Retroviridae тұқымдастығына кіретін 150-дей түрлерді қамтитын біржіпшелі РНҚ- құрамды кері транскриптаза (ревертаза) ферменті бар вирустар. Олардың ерекшелігі- геномының құрылысы басқа вирустарға ұқсамайды және құрамында кері транскриптаза (РНҚ -тәуелді ДНҚ- полимераза) болады. Кері транскриптаза генетикалық ақпараттың кері бағытталуын, яғни ДНҚ-нан ДНҚ-на емес, керісінше РНҚ-нан ДНҚ-на қарай жүруін қамтамасыз етеді. Осыған байланысты тұқымдастықтық ретровирустар (ағылшынша: retro-кері) деп аталған.

Таксономиясы
Тұқымдастығы: Retroviridae
Туыстастықтары: Alpharetrovirus- құстардың лейкозы, саркома вирустары.
Betaretrovirus- тышқандардың сүт бездері ісігінің вирусы.
Grammaretrovirus- тышқандар лейкомиясының вирусы.
Deltaretrovirus- сиырлар лейкомиясының вирусы, НТLV- 1, 2
Epsiloretrovirus- тері саркомасының вирусы.
Lentivirus- адамдардың иммунды- тапшылық вирусы (НІV) .
Spumavirus- адамдардың көпіршік вирустары.
Адамдар патологиясында ретровирустардың бірнеше түрлері (АИВ-1, АИВ-2, Т- жасушалық лейкоздар вирустары ( НТLV- 1 және НТLV- 2) маңызды орын алады.

Адамдардың иммундық- тапшылық вирусы (АИВ--НІV) .
АИВ- лимфоциттерде, макрофагтарда және нерв тіндерінің жасушаларында вирустың ұзақ уақыт айналымда болуының нәтижесінде иммундық жауаптың прогрессивті бұзылуын қоздыратын вирустар. Жүре пайда болған иммундық тапшылық синдромы (ЖИТС- СПИД) ерекше инфекция ретінде алғашқы рет 1981 жылы АҚШ-та пневмоцисттік өкпе қабынуының өте ауыр түріне шалдыққан жігіттерде (гомосексуалисттерде) тіркелеген. Пневмоцисттер (мысалы, Pneumocytes carinii- саңырауқұлақтарға жатады) әдетте адамдар үшін патогенді емес немесе паогенділігі төмен. Ауруға шалдыққандарға пневмоцисттік және өкпе қабынуымен қатар басқа да оппортунисттік инфекцияларға сезімталдығы жоғары екені байқалған. Осындай ауруға шалдыққандардың иммундық статусын зерттегенде лимфоциттердің, әсіресе Т-хелперлердің, күрт азайғаны анықталған. Осындай жағдай ЖИТС- СПИД- AIDS (ағылшынша: Acguired Immune Deficiency Syndrome) деп аталынады. ЖИТС қоздырғышын әр қайсысы бір- бірінен тәуелсіз екі ғалым ашқан. Л. Монтанье (францияда) өзі ашқан вирусты LAV (Lympha adenopathy associated virus) деп атаған, өйткені бұл вирус лимфоаденопатиямен ауырған науқастан табылған. Р. Галло (АҚШ-та) НТLV- ІІІ деп белгілеген (ағылшынша: Human T- lymphotropic virus III) . Лимфотропты вирустарды салыстырып зерттегенде олар бір- біріне сәйкес екені анықталып, оларды АИВ- ВИЧ- НІV (ағылшынша: Human Immunodeficiency virus) деп атау қабылданды.

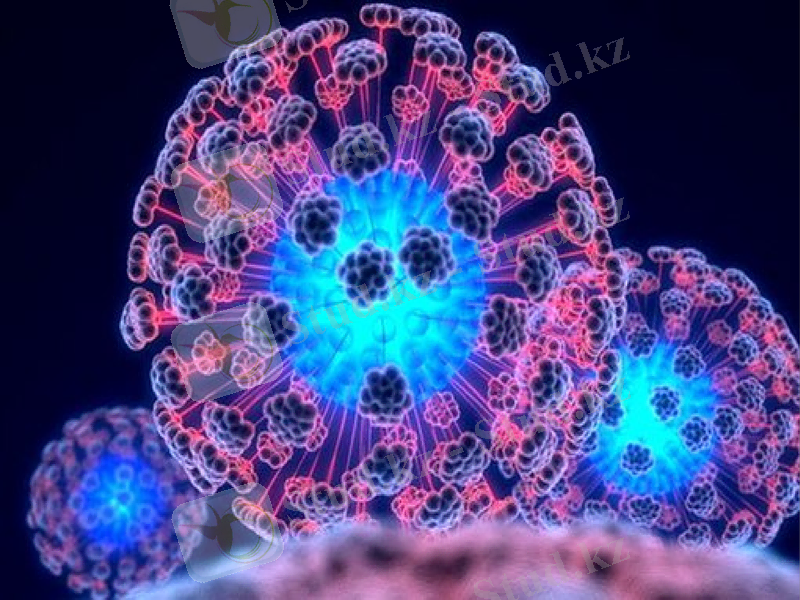
Морфологиясы
АИВ- РНҚ құрамды вирус. АИВ-тің жетілген вириондары сфера пішінді, мөлшері 100-120 нм-ден аспайды (13-сурет) . Сыртқы қабықшасы (суперкапсиді) гликопротеинді тікенекшелермен (gp 160) шаншылған липидті екі қабаттан тұрады. Тікенекшелері gp-41, gp-120ақуыздарынан құрылған. Гликопротеин gp-120 тікенекшенің капсидтен шығып тұрған бөлшегінде орналасқан және жасуша мембранасындағы CD-4 молекуласымен өзара әрекеттеседі. Гликопротеин gp41 (біріктіру ақуыздары) сыртқы қабатының ішкі жағында орналасады және жасушалық мембранасының қосылуын қамтамасыз етеді. Вирионның жүрекшесі цилиндр немесе конус пішінді, капсидтік р18 және р24 ақуыздардан тұрады. Матрикстік ақуыз (р17) вирион жүрекшесі мен сыртқы қабырғаның арасындағы қабықшаны қалыптастырады. Жүрекшеде РНҚ, ішкі нуклеокапсидтік ақуыздар (р7, р9), кері транскриптаза, интеграза, протеаза орналасады.

Геномы
Геномы (+) РНҚ- ның екі жіпшесінен тұрады, құрамында 7900-9800 нуклеотидтік қосақтар және вирустық үш фермент (кері транскриптаза, протеаза және интеграза) бар. Вирус геномы негізгі 3 құрылымдық геномдерден (gag, pol, env), 7 реттегіш және функционалдық гендерден (tat, rev, nef, vif, vpr, vpu, vpx) тұрады.
Ген gag (ағыл: group antigen-топтық антиген) - матрикстік, капсидтік, нуклеокапсидтік және протеазалық ақуыздарды кодтайды.
Ген pol (ағыл: polymerase- полимераза) - кері транскриптазаны, протеазаны, РНҚ-азаны, интегразаны кодтайды.
Ген env (ағыл: envelope- қабықша) - беткейлік gp-120 және трансмембраналық gp-41 ақуыздарды кодтайды.
Ген tat- транскрипциялануды белсендіруші, ол құрылымдық, және де реттегіш ақуыздардың транскрипциялану жылддамдығын бірнеше есе күшейтеді.
Ген vpr- ревертазаны реттейді, РНҚ және ДНҚ құрылымын тұрақтандырады, ДНҚ-н жасуша ядросына тасымалдайды және де вирустың макрофагпен өзара әректтесуі үшін маңызы бар деген болжамдар айтылған.
Ген vpx- жасушалардың онкогендік трансформациялануына жауапты.

Антигендік құрылымы
АИВ-ң негізгі антигендері- беткейлік типтік спецификалық gp41 және gp120 ақуыздар. Жүрекшелік gp24- топтық спецификалық антиген. Вирустың екі типін (АИВ-1, АИВ-2) және 10- нан астам серологиялық вариантарын ажыратады. Ерекше атап өту керек- вирус өте жоғары антигендік өзгергіштігімен айрықшаланады, оның өзгергіштігі грипп вирусына қарағанда жүздеген есе артық. АИВ-1 үшін мутациялану мүмкіндігі репликациялану циклында 10-4 -10-5 дәрежеде деп болжамданады.

Репродукциялануы
АИВ-ң тіршілік циклы 4 сатыдан тұрады
1) вирустың жасушаға адсорбциялануы және оған эндоцитоз жолымен енуі;
2) вирустық РНҚ-ң босанып шығуы, ДНҚ провирусының синтезделуі және провирустың жасуша геномымен интеграциялануы;
3) вирустың РНҚ-ң синтезделуі, трансляциялануы және вирустық ақуыздардың қалыптасуы;
4) вирион құрастырылуы, олардың жетілуі және жаңадан пайда болған вириондардың бүршіктену тәсілімен жасушадан шығуы
Вирустың толық тіршілік циклы 1-2 тәулікте іске асады, тәулігіне 1 млрд-қа дейін вирустық бөлшектер пайда болады.

Вирустың гликопротеині gp 120 Т-хелперлердің, макрофагтардың, дендритті және микроглиалдық жасушалардың CD 4 рецепторларымен байланысады. Жасушалардың басқа да корецептарлы қатысады. АИВ қабаты жасуша плазмалеммасымен қосылғаннан кейін жасуша цитоплазмасында геномдық РНҚ және вирустың кері транскриптазасы (ревертаза) босанып шығады да, олар геномдық РНҚ-ң матрицасында комплементарлы минус-жіпшелі ДНҚ-ң (сызықшалы бір жіпшелі кДНҚ) синтездейді. Соңғыдан плюс-жіпшелі ДНҚ-ң көшірмесі жасалып, сақиналы екі жіпшелі ДНҚ пайда болады. Екі жіпшелі вирустық ДНҚ (провирус) интеграза(ие жасушасының ДНҚ-ын кескілейді) ферментінің көмегімен жасушалық ядроға енеді. Иесінің жасушалық ДНҚ бөлшектенеді (кесіледі), провирус хромосоманың ДНҚ-на тіркеледі, яғни интегрирленеді де жасуша тіршілік етуін тоқтатқанша сақталады. Вирус енген Т-лимфациттердің микробтық антигендермен немесе басқа иммунды-компетентті жасушалармен белсенділігі артқанға дейін провирус тыныштық жағдайда болады. Жасушалық ДНҚ-ң транскрипциялануының кушеюі ерекше ядролық фактормен реттеледі. Бұл ақуыз жасушалық ДНҚ-ң да, әрі провирус ДНҚ-ң да транскрипциясын индуцирлейді. Нәтижесінде белсенсіздіктен белсенді (активті) жағдайға ауысады, осыған орай персистенттік инфекция өнімді инфекцияға айналады. Вирустың белсенсіз жағдайда болуы өте ұзақ мерзімге созылуы ықтимал. Жасушаның вируспен инфицирленген сәтінен бастап вирус тасымалдаушы кезеңі басталады, ол 10 жыл және одан да ұзақ уақытқа созылады. Вирустың белменділігі күшейген сәттен бастап нағыз ауру- АИВ инфекция байқалады. Реттеуші гендердің және олардың өнімдерінің көменгімен вирус белсенді көбейе бастайды. Tat -ақуыз вирустың репродукциялану жылдамдығын 1000 есе арттырады. Провирустың рекомбинантты ДНҚ бір мезгілде жаңа вириондарды қалыптастыруы үшін ДНҚ- тәуелді РНҚ-полимеразының көмегімен геномдық РНҚ-ң, және де жасушалық рибосомаларда вирустық протеиндер мен энзимдердің (gag және pol) синтезделуі үшін негізін қалаушы функциясын иРНҚ атқарады. Осындай екі протейы ұзынша полипептитты тізбекшелер түрінде цитоплазмада синтезделеді. Содан кейін олар протеазалардың көмегімен жекеленген құрылымдық ақуыздарга кескілейді. Транслирленетін Еnv ақуыз (gp 160 - тың ізашары) жасушалық протеазының әсерімен молекулаға (gp 120- және gp 41) ыдырайды. Вирустың бөлшектің кұрастырылуы жасушалық мембрананың ішкі жағында атқарылады. Вирустың қалыптасуы кезінде оған РНҚ- ның 2 жіпшесі тартылады . Вирион жасушадан бүршіктеніп шыққан кезде иелік- жасушаның бөлшегін (участкесін) ала кетеді. Мембрананың бүтіндігі бұзылады да, жасуша өледі.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz