Өсімдіктерге стресс факторлары және олардың физиологиялық және молекулалық төзімділік механизмдері


ӨСІМДІКТЕРГЕ СЫРТҚЫ ОРТАНЫҢ ҚОЛАЙСЫЗ ЖАҒДАЙЛАРЫНЫҢ ӘСЕРІ

Стресс физиологиясы
Өсімдіктерге әсер ететін коршаған ортаның колайсыз жағдайларын стрессорлар деп атайды. Өсімдіктердің жағымсыз жағдайлар әсер еткенде оларда пайда болатын күйзеліс, абыржу жағдайлары стресс деп аталады. Г. Селье. 30-ж. ж. - медицинада «стресс» деген термин ұсынды (ағылшын сөзі stress - күйзеліс) .
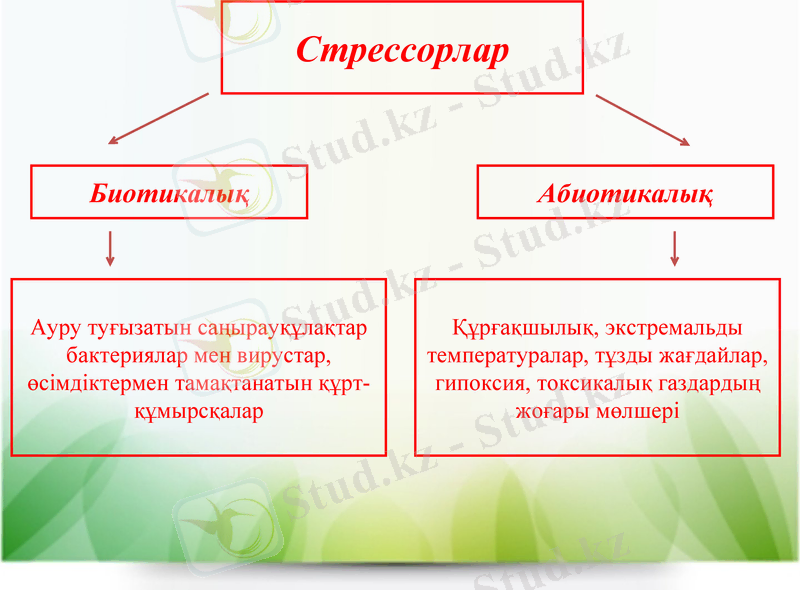
Стрессорлар
Биотикалық
Абиотикалық
Ауру туғызатын саңырауқұлақтар бактериялар мен вирустар, өсімдіктермен тамақтанатын құрт-құмырсқалар
Құрғақшылық, экстремальды температуралар, тұзды жағдайлар, гипоксия, токсикалық газдардың жоғары мөлшері
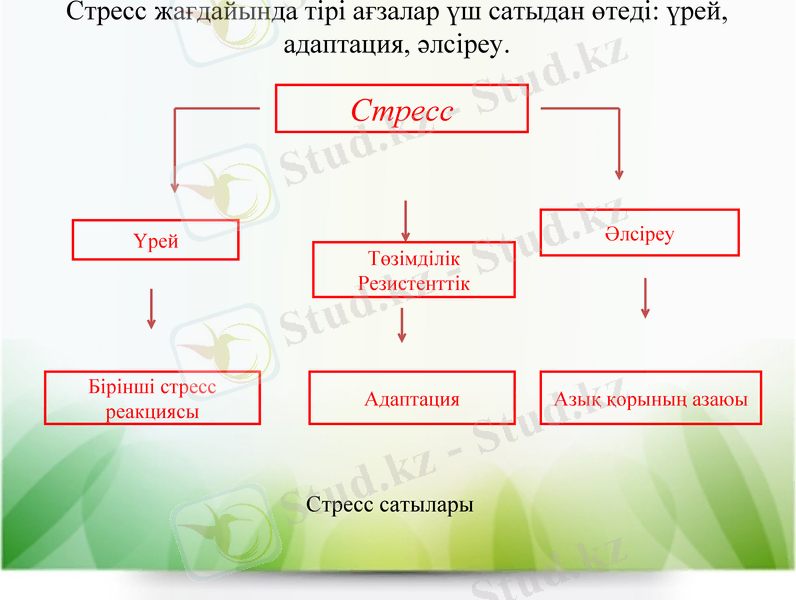
Стресс жағдайында тірі ағзалар үш сатыдан өтеді: үрей, адаптация, әлсіреу.
Стресс
Үрей
Төзімділік
Резистенттік
Әлсіреу
Бірінші стресс реакциясы
Адаптация
Азық қорының азаюы
Стресс сатылары

Стресс механизмдері. Д. Н. Насонов пен В. Я. Александров 1940 ж. бірінші рет сыртқы жағдайларға карсы жалпы, барлық стреске тән өзгеше емес реакциялар кешені деген термин кіргізді. Ол кешен пара-некроз деп аталады. Өсімдіктердің сыртқы ортаның қолайсыз жағдайлары әсер еткенде ішкі ортаның ұдайылығын сақтау қабілеті (гоместаз сақтау) және өмір сүру үдерістерін жүзеге асыруы төзімділік деп аталады.

Жалпы реакцияларға жататын үдерістер:
1. Клетканың мембранасының өткізгіштігі ұлғаяды, мембрандык потенциалы деполяризацияға ұшырайды.
2. Са2+-дің цитоплазмаға шығуы (клетканың кабығынан жэне
клетканың компартменттерінен: вакуольдан, ЭР, митохондрия-
лардан) .
3. pH мәні кышқыл жаққа жылжиды.
4. Актиндік микрофиламенттердің жиналуы ырықтанады. Ол цитоплазманың тығыздығын ұлғайтады.
5. Өсімдіктердің оттекті пайдалануы күшейеді, АТФ-ті пайда-
лануы тездетіледі, бос-радикалдық реакциялардың белсендігі
ұлғаяды.

6. Гидролиз үдерістерінің жүруі күшейеді.
7. Стресс белоктар синтезделеді де, олар белсенеді.
8. Плазмалеммада Н+-помпаның жүмысы күшейеді (мүмкін, тоно- пласта да) . Ол иондык гоместазды қолайсыз жаққа ысырылуына
кедергі жасайды.
9. Этиленнің және АБҚ-ныц синтезделуі, клетканың өсуі және
бөлінуі, тамырлардың copy белсенділігі және басқа физиоло-
гиялық және метаболикалық үдерістердің жүруі баяулайды.
Ингибиторлардың әрекеттерінің және энергетикалық ресурс-
тардың ауыстырылу нәтижесінде клеткалардың функционалдык
белсенділігі төмендейді.

2-ші фаза - адаптация. Гидролиз және катаболизм реакциялары белсендігінің төмендеуі, синтез үдерістерінің ұлғаюы. Ыдырау өнімдері зат алмасудың қайта кұруына себеп болады. Адаптация үдерісі барысында бірнеше принцип жүзеге асады: артықшылық принципі (өсімдік дәндерінің көпшілігі), бір маңызды компоненттердің гетерогендігі (ферменттердің изоформалары бірдей реакцияны катализдейді, бірақ олар әртүрлі сыртқы ортаның факторларының диапазонында жұмыс істейді.

3-ші фаза - әлсіреу фазасы. Клетка органеллалары мен құрылымдарының бұзылуы: ядроның деструкциясы, хлоропластарда граналардың ыдырауы, митохондрияда кристалдардың мөлшері азаяды. Қосымша вакуольдер пайда болады. Оларда стресс жағдайында зат алмасу өзгергеннен пайда болған зиянды заттар залалсызданады.
Негізгі энергия генераторларының - митохондриялардың және хлоропластардың ультрақұрылымы бұзылады. Ол энергияны жоғалтуға әкеледі, цитоплазманың физикалық және химиялық қалпын өзгертеді.
Регенерация фазасы (реституция) - стрессор жоғалғанан кейін пайда болады. Онтогенез барысында өсімдіктердің төзімділігі өзгереді: өсімдіктер ювенильдік фазада (өнгенде), гүлдену және жемістену фазаларында өте сезімтал, ал тыныштык фазасында төзімді болады.
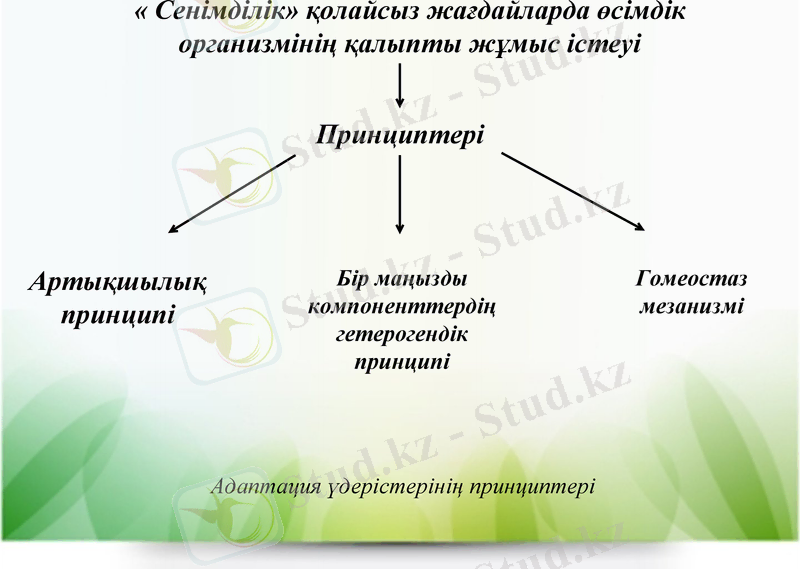
« Сенімділік» қолайсыз жағдайларда өсімдік организмінің қалыпты жұмыс істеуі
Принциптері
Артықшылық принципі
Бір маңызды компоненттердің гетерогендік принципі
Гомеостаз мезанизмі
Адаптация үдерістерінің принциптері

Стрессорлардың әсеріне қарсы өсімдіктердін бейімделу стратегиясы. Өсімдіктерде стрессорлардың әсерінен байқалатын құбылысты екі санатқа бөлуге болады:
1) зақымдану, яғни өсімдіктің әртүрлі кұрылымдық және функционалдық ұйымдасуы деңгейінде байқалады, мысалы, белоктық молекулалардың денатурациялануы, зат алмасудың бұзылуы және кұрғақшылық немесе топырақтың тұздануы жағдайында клеткалардың сусыздануынан клетканың созыла өсуінің тежелуі;
2) жаңа стресс жагдайларына бейімделуді қамтамасыз ететін
өсімдіктердің жауап реакциялары. Олар гендердің экспрессия-
сын өзгертеді, зат алмасуға, физиологиялық функцияларға және
гомеостазға әсер етеді.
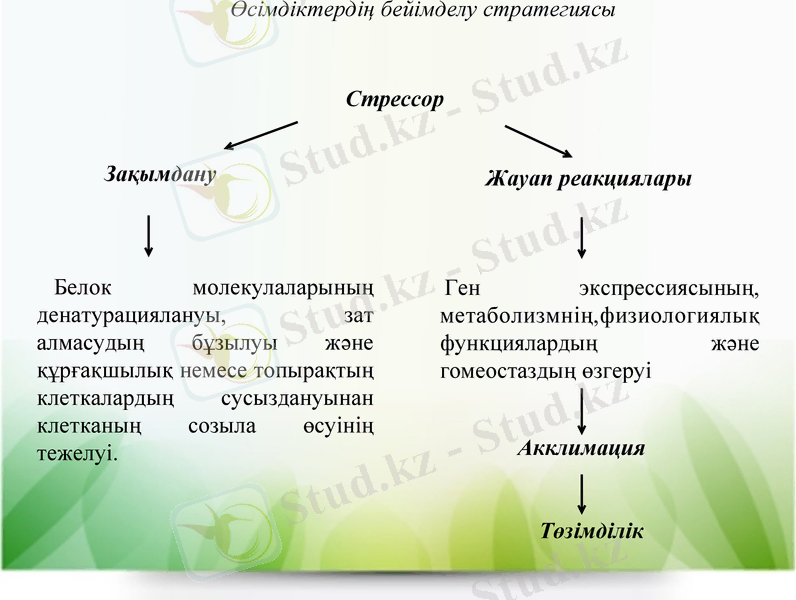
Стрессор
Зақымдану
Жауап реакциялары
Белок молекулаларының денатурациялануы, зат алмасудың бұзылуы және құрғақшылық немесе топырақтың клеткалардың сусыздануынан клетканың созыла өсуінің тежелуі.
Ген экспрессиясының, метаболизмнің, физиологиялық функциялардың және гомеостаздың өзгеруі
Акклимация
Төзімділік
Өсімдіктердің бейімделу стратегиясы

Акклимация үдерісі барысында стрессорлардың әсеріне карсы өсімдік төзімділікке ие болады. Акклимация организмнің тіршілігі кезінде пайда болады және ол тұқым қуаламайды (4-сурет) . Акклимацияда генотиптегі материалдың мүмкіндіктері негізінде жүзеге асады, яғни реакция нормасы шегіндегі амплитудасымен қамтамасыз етіледі - генотиптің жүзеге асуындағы мүмкін болатын өзгерістер. Бұл үдерісте өсімдікте төзімділік пайда болады.
Адаптациялар - тұқым куалайтын қасиет, оған өсімдік стресс жағдайына тәуелсіз ие болады. Өсімдіктердің әртүрлі стрессорлардың әсеріне төзімділігіне бейімделудің маңызы зор. Бейімделудің акклимациядан айырмашылығы - ол өсімдіктің стресс жағдайының болу-болмауына тәуелсіз өсімдікте болатын тұқым қуалайтын бекітілген конститутивті белгі. Мысалы, қоршаған ортаның кұрғақшылығына өсімдіктердің бейімделуі, әсіресе суккуленттердің, кактустардың морфологиялык ерекшеліктерін айтуға болады.

Акклимация
Адаптация
Организмнің тіршілігі кезінде пайда болады және ол тұқым қуаламайды
Өсімдіктің стресс жағдайында болу - болмауына тәуелсіз тұқым қуалайтын белгі
Шынықтыру
Күздік астық тұқымдастар
Суккуленттердің морфологиялық ерекшеліктері
Қыс кезеңінде төмен температура жағдайына төзімді болады, төмен оң және 0 С-қа жақын теріс температура әсерінен шынығады
Акклимацияның және адаптацияның айырмашылықтары
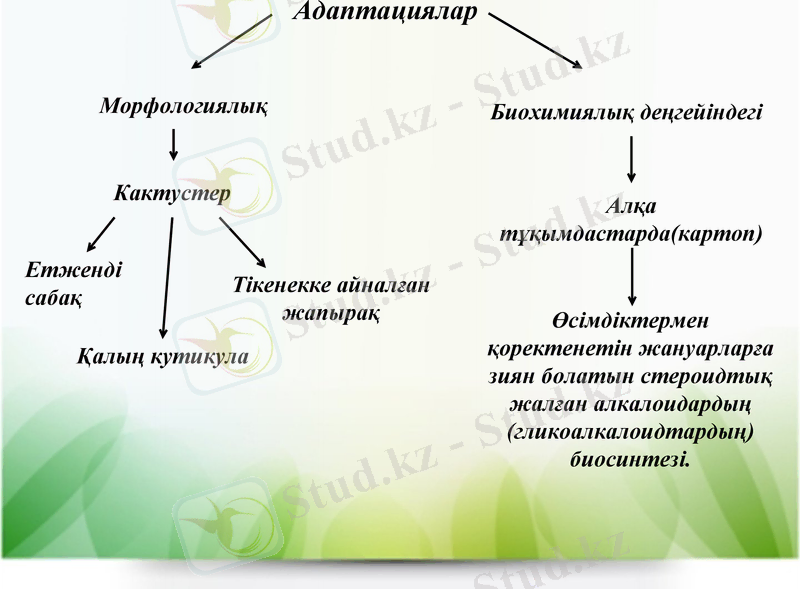
Адаптациялар
Морфологиялық
Биохимиялық деңгейіндегі
Кактустер
Етженді сабақ
Қалың кутикула
Тікенекке айналған жапырақ
Алқа тұқымдастарда(картоп)
Өсімдіктермен қоректенетін жануарларға зиян болатын стероидтық жалған алкалоидардың
(гликоалкалоидтардың) биосинтезі.

1. Қашу механизмі. Олар өсімдіктерге стрессорлардың әсерінен қашуға мүмкіндік береді. Ондай механизмдерге өсімдіктердің топырактың терең кабатына енген тамыр жүйелері аркылы суды сіңіруді жаткызуға болады. Біркатар ксерофиттерде (кұрғақшылық жерде өсетін өсімдіктер) мысалы, қара сексеуілде тамыр жүйесінің ұзындығы бірнеше ондаған метрге жетеді. Бұл өсімдіктерге жерасты суларын пайдалануды және топырақ және атмосфералық құрғақшылық жағдайында ылғалдың жетіспеушілігінен қорғайды. Осы санатқа топырақтың тұздылығына төзімді өсімдіктер цитоплазмасының иондық гомеостаз механизмін жатқызуға болады.

Конститутивті және акклимация барысындағы қорғаныс мезанизмдері
Қашу механизмдері
Резистенттік механизмдер
Өсімдіктерге стрессорлардың әсерінен қашуға мүмкіндік береді
Өсімдіктер стрессорлардың әсерінен қашпай-ақ стрестік жағдайлардан тез өтеді
Қуаншылыққа төзімді өсімдіктердің ұзын тамыр жүйесі
Тұзды жағдайға төзімді өсімдіктердің иондық гомоестаз мезанизмдері - Na - мен СІ-дың төмен концентрациясын сақтау
Бірдей реакцияны катализдейтін изоферменттердің биосинтезі

2. Резистенттік механизмдері (төзімділік) . Өсімдіктер осы механизмдер барысында стрессорлардың әсерінен қашпай-ақ, стрестік жағдайлардан өтеді. Бұндай механизмдерге, әсіресе сол реакцияларды катализдейтін бірнеше изоферменттің биосинтезі жатады. Қоршаған ортаның бірқатар параметрлерінің тар диапазон шеңберінде әр изоформаға қажетті каталитикалық қасиетке ие болады, мысалы, температураны айтуға болады. Тек бір изоферментпен салыстырғанда, барлық изоферменттердің жиынтығы, тұтастай алғанда, өсімдіктің кең температуралық диапазонда реакцияларды жүзеге асыруына мүмкіндік береді. Яғни температуралык өзгеріс жағдайына бейімдірек болады.

Су тапшылығы
Егер транспирация жылдамдығы тамыр жүйесініц суды сіңіру жылдамдығынан жоғары болған жағдайда өсімдік су тапшылығына ұшырайды. Бұндай жағдай кұрғакшылық кезінде ғана емес, топы- рақтың тұздануы кезінде де және өте төмен температура жағдайында да байқалады. Соңғы екі жағдайда дегидратация табиғаты «сусыз» стресс құрамдас болып саналады және ол өсімдікте көбінесе топырақтағы судың мөлшері шектелмеген жағдайда дамиды. Бірақ бұл судыөсімдік қабылдай алмайды. Мысалы, топырактың тұздануы жағдайында топырақтың осмостық қысымының жоғары болуына байланысты тамыр жүйесі суды сіңірмейді (су потенциалы төмен) .

Су тапшылығы кезінде клеткадағы судың мөлшерінің азаюы және сусыздануға әкелетін цитоплазмадағы иондар концентрациясының артуы әртүрлі кұрылымдық бұзылыстарға және биополимер функцияларының, әсіресе белоктардыцң денатурациялануы және оның ферментативтік белсенділігініц тежелуі байқалады.

Мембрананыц билипидтік құрылымы өзгереді және оның бүтіндігі бұзылады. Мембранадағы деструктивті өзгерістер заттардың клетка ішілік бұзылуына және электрогенездің тежелуіне әкеледі. Клетка деңгейіндегі су тапшылығы деп тургорлық кысымның азаюын айтамыз. Тұтас өсімдік деңгейінде су тапшылығы су потенциалыныц градиенті топырақ-тамыр-өркен жүйесінде және өсудің тежелуі арқылы байқалады.

Су тапшылығына төзімділік механизмдерінің индукциясы нәтижесінде өсімдіктер стрессорлардың әсеріне төзімдірек бола бастайды. Төзімділіктің дамуы қашу және тұрақтылық механизмдері индукцияларының нәтижесі болуы мүмкін. Реакциялардың екі санаты су тапшылығына өсімдіктің акклимация үдерісі катысты екенін төменде толығынан қарастырады.

Су тапшылығындағы өсімдік клеткасындағы қашу стратегиясы. Су потенциалы градиенті бойынша су пассивті түрде жоғары аймақтан төмен аймаққа карай козғалады Ψ. Егер тамыр клеткасының су потенциалы Ψкл топырақтың су потенциалынан жоғары болса, онда өсімдік топырақтан суды сіңіре алмайды. Су топырақтан тамыр клеткаларына қарай қозғалу үшін Ψopт қарағанда Ψкл өте төмен деңгейге дейін жету керек. Кейбір өсімдіктер су тапшылығына өте сезімтал болады және Ψкл өте төмен болғанда, тез солып қалады (тургордан айырылу) .

Құрғакшылыкқа және тұзға төзімді өсімдіктер клеткалары өздерінің су патенциалын клетка ішілік осмостық кысымды 7СКЛ арттыру есебінен төмендете алады. Бұл осмостық реттелу үдерісіне жатады. Клеткадағы осмостық белсенді заттардың жалпы концентрациясының артуына байланысты 7ГКЛ ұлғаяды. Оның әсерінен клетка ішілік осмос дегидратациясында заттардың пассивті жинақталуы нәтижесінде пайда болған қысымға қарағанда жоғары болады. Су тамырға πкл артуына байланысты су потенциалының градиенті бойынша енеді. Осмореттелу үдерісі өсімдіктің акклимациясында құрғақшылық және топырақтың тұздануы жағдайында маңызды рөл атқарады .

Су тапшылығы жағдайында осмореттеу төмен молекулалық заттардың синтезделуінің арқасында жүзеге асады. Цитоплазмада осмолиттер синтезделеді, вакуольде иондар жинақталады. Клетканың ішіндегі осмостық кысым ұлғаяды, судың потенциалы төмендейді. Ол клетканың суды соруына әкеледі. Ал клеткаларда егер осмореттеу үдерісі төмен деңгейде жүрсе, тургор бұрынғы қалпына келмейді. Осмостық қысым берілген көлемде еріген жалпы бөліктердің санының қызметі болып табылғандықтан, артуына төмен молекулалы органикалық заттармен қоса бейорганикалқ иондар да катысады. Клетка цитоплазмасында су және тұз стрестерінде осмостык кысымның реттелуі төмен молекулалы органикалық заттардың биосинтезі есебінен жүзеге асады және оларды осмолиттер деп атайды.
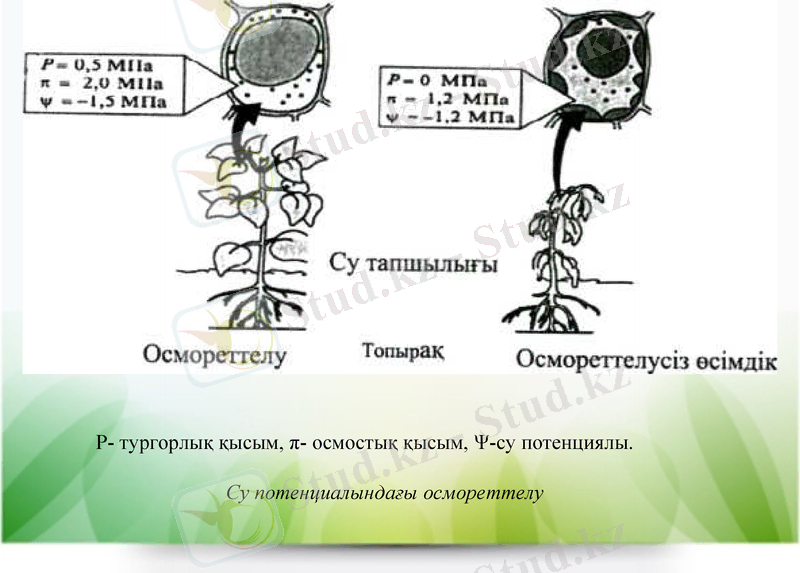
Р- тургорлық қысым, π- осмостық қысым, Ψ-су потенциялы.
Су потенциалындағы осмореттелу

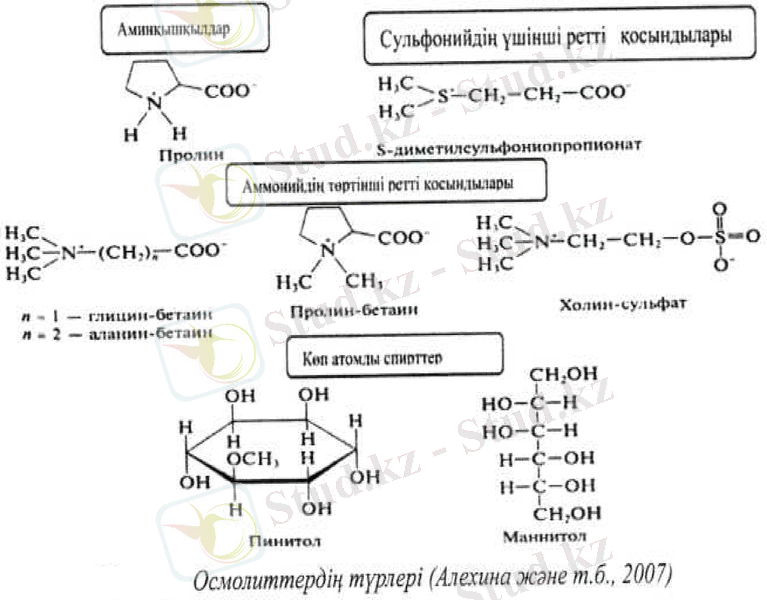
Осмолиттер. Осмолиттердің қасиеттері және қызметтері. Осмолиттер - химиялық табиғаты бойынша алуан түрлі төмен молекулалы органикалық қосылыстар. Олар суда жақсы ериді, улы емес және бейорганикалық иондармен салыстырғанда, метаболизмде өзгерістер туғызбайды. Сондықтан да олардың екінші аты - үйлесетін заттар (compatible solutes) . Клетка цитоплазмасындағы табиғаты органикалық осмолиттердің синтезі және жинақталуы - өсімдік әлемінде кең тараған құбылыс.

Үйлесетін заттар клеткада нейтральды болады. Олар цитоплазмада диссоциацияланбаған формада немесе цвиттерион түрінде, яғни теріс және оң зарядтарды алып жүретін кеңістік бойынша бөлінген молекулалар түрінде болады. Кейбір осмолиттер амфифильді косылыстар болып табылады. Амфифилъді заттар молекулаларында полярлы емес (гидрофобты), сонымен бірге полярлы (гидрофильді) топтары болады. Кең тараған осмолиттердің химиялық құрылымы келесі кестеде бейнеленген.

Осмолиттердің жалпы функциясы - осмореттеу үдерістеріне қатысу. Көптеген бейорганикалық иондар, мысалы, К+ және С1 иондарының жоғары концентрациясы улы, сондықтан өсімдік клеткасында цитоплазманың осмостық қысымын реттеуге пайдалану мүмкін емес. Алайда биополимерлермен үйлескен осмолиттер цитоплазмада грамға шаққанда бірнеше жүз микромольге дейін жинақтала алады және ол ешқандай улы әсер етпейді.

Осыған байланысты өсімдіктер цитоплазманың осмостық қысымын реттеу үшін клеткадағы бейорганикалық иондарға жатпайтын осмолиттерді пайдаланады. Осмолиттердің рөлі әсіресе кұрғақшылық пен тұздану жағдайында, клеткаға осмостық белсенді заттарды жинактау кезінде маңызды. Дегидратацияға төзімділік бойынша өсімдіктердің айырмашылығы осмолиттер биосинтезі жүйесінің тиімділік деңгейіне байланысты болады.
Ксерофиттер және галофиттер, тиісінше, - ылғалдылығы төмен және топырақтың тұздану жағдайында өсетін өсімдіктер.

Вакуольде осмореттелу, негізінен, бейорганикалық иондардың - К+, Са2+ жэне С1“ жинакталу жолымен жүзеге асады. Осмолиттер есебінен цитозолдің, вакуольдің және клетканың басқа қалган барлық органнеллаларының осмостық тепе-теңдігі орнайды. Осмореттелумен катар үйлесетін заттар дегидратацияда өте маңызды кызмет аткарады. Бұл функция цитоплазмалық биоөріске байланысты қорғаныс (протекторлық) ретінде аныкталуы мүмкін. Осмолиттердің қос функционалдык рөлі ескеріліп, олар, көбінесе осмопротекторлар деп атайды. Қазіргі уақытта олардың қорғаныс қасиеттері туралы көптеген мәліметтер жинақталуда. Бірақ айта кететін жағдай - үйлесетін заттардың қорғаныстық әсерінің дәлелі негізінен алынды. Мысалы, өсімдік жапырағы препаратында жүргізілген тәжірибеде глицин-бетаин инактивтелуін және фото жүйе II кешенде бөлінетін О, дестабилизациялануын жояды.

Осмолиттер биополимердің гидратты қабықтарын бұзбайды деп есептеледі. Стрессор әсерінен синтезделетін кейбір төмен молекулалы органикалық қосылыстар мардымсыз мөлшерде синтезделеді және ол осмореттеу қызметін атқара алмайды. Алайда олар стресс жағдайында биополимерлер протекторы ретінде маңызды рөл атқарады. Ондай қосылыстарға диамин путресцин және одан түзілетін полиамин спер- мидин және спермин жатады.
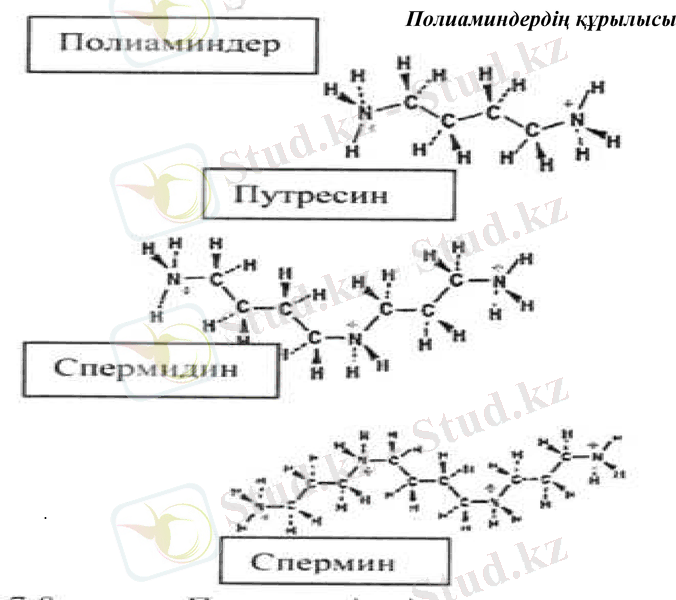
.
Полиаминдердің құрылысы

Полиаминдер (ПА) өсімдіктерде көп тараған. Олар ДНҚ, РНҚ және белоктардың синтезін карқындандырады. Поликатион ретінде, полиаминдердің биомолекулалармен әрекеттесу қабілеті өте жоғары болады, әсіресе ДНҚ, РНҚ, фосфолипидтермен және қышқыл белоктармен, сонымен қоса мембрана компоненттерінің және клетка қабығының анион топтарымен. Өсімдіктерде полиаминдер көптеген физиологиялық үдерістерге, атап айтсақ, клетканың бөлінуіне, цитоскелеттің қалыптасуына, тамырдың өсуін жеделдетуіне, эмбриогенезге және жемістердің пісіп-жетілуіне қатысады.

Полиаминдер кұрғакшылық, тұздану, төмен температура және озон әсерінен туындайтын биомолекулалардың зақымдануынан сақтайды. ДНҚ, РНҚ, рибосома, сонымен қоса, полиаминдер кешеніндегі мембраналардың құрылымын тұрақтандырады. Стресс жағдайында артқан РНҚ-аза мен протеазалардың белсенділігін төмендетеді, полиаминдер белок синтезі кезіндегі акпараттарды дәл есептейді және клеткалық құрылымдардың ыдырауын азайтады.

Дегидратация жағдайында түзілетін белоктар. Әртүрлі қызметтер атқаратын белоктар жауап реакция ретінде de novo синтезделенеді.
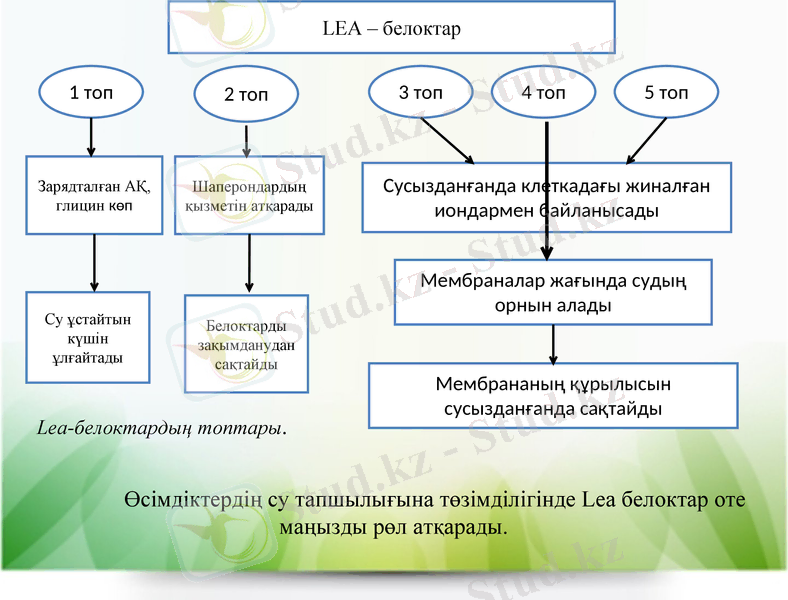
Lеа-белоктар. Кейбір су тапшылығында пайда болатын белоктар цитоплазмалық биополимерлерді және клеткалық құрылымдарды дегидратация себебінен закымданудан қорғап түрады. Оларга Lea (late embryogenesis abundant) - белоктар жатады. Олар пісіп және қурап жатқан дәндердің ішінде экспрессияланатын LEA-гендердің әсерінен пайда болғанда ашылған. Соңынан кейбір Lеа-белоктар су тапшылығы жағдайында, тұзды ортада және төмен температураларда вегетативтік мүшелерде табылған.

Lеа-белоктар гидрофильді белоктарға жатады. Олардың кұрамында аланин және глицин деген аминкышқылдар көп болады да, цистеин мен триптофан болмайды. Олар кұрамына кіретін аминқышқылдар тізбегіне және олардың кұрылысына байланысты бес топқа бөлінеді.
1 топ - зарядталған аминкышқылдар және глицин көп белоктар, суды жақсы байланыстырады. Олар цитоплазмада орналасқан, су ұстайтын қабілетін жоғарылатады.
2 топ- шаперондардың қызметін атқарады. Олар басқа белоктармен байланысып, соңғылардың закымдануынан корғайды.

3. 5 топтар - су тапшылығы жағдайында цитоплазмада жиналатын иондармен байланысады. Гидрофобтың ауданымен белоктар гомодимерлер түзеді де, зарядталған жағымен иондарды байланыстырады. 4 топ - мембрана жағында судың орнын алады. Осылай дегидратация жағдайында олар мембрананың кұрылымын сактайды.
LEA - белоктар
1 топ
5 топ
4 топ
3 топ
2 топ
Зарядталған АҚ, глицин көп
Шаперондардың қызметін атқарады
Су ұстайтын күшін ұлғайтады
Белоктарды зақымданудан сақтайды
Сусызданғанда клеткадағы жиналған иондармен байланысады
Мембраналар жағында судың орнын алады
Мембрананың құрылысын сусызданғанда сақтайды
Lea-белоктардың топтары.
Өсімдіктердің су тапшылығына төзімділігінде Lea белоктар оте маңызды рөл атқарады.

Шаперондар мен протеазалардың ингибиторлары. Ширатылып-бүктеліп, оралып (folding) жатқан полипептидтермен байланысатын белоктарды, белоктардың 3-ші және 4-ші реттік құрылым суббірліктерімен, полипептидтермен байланысып, шаперондар белоктардың қателіксіз агрегацияланбай ширатылуын қамтамасыз етеді. Кейбір шаперондар «жөндеу станцияларының» рөлін атқарады, олар белоктардың қате ширатылғанын жөндейді. Шаперондардың ең басты функциясы - мембраналардан өткенде белоктарды ширату және өздігінше ұзындап орналастыру (unfolding), және белоктарды жинақтау мен тарату етеді. Осмостық стресте протеазалардың ингибиторлары көп синтезделеді. Олар белоктардың ыдырауын тежейді де, олардың құрылымын сақтап калады.

Протеазалар мен убиквитиндер. Су тапшылығы жағдайында протекторлық қасиеттері бар қосындылар және шаперондар барына қарамастан, кейбір клеткалық белоктар денатурацияға ұшырайды. Денатурацияланған белоктар гидролизденуі керек. Ол функцияны протеазалар және убиквитиндер атқарады, олардың гендері стресс жағдайларында экспрессияланады.
Убиквитиндер - молекулалық салмағы төмен (8, 5 к Да) белоктар. Олар денатурацияланған белоктың N ұшына байланысып, соңғыларды протеазалардың жұмысымен камтамасыз етеді.

Аквапориндер. Су мембранадан каналдар арқылы өтеді. Ол каналдарды белоктар аквапориндер құрады. Каналдардың өткізгіштігі және саны өзгергеніне қарай судың трансмембрандық жылжуы реттеліп тұрады. Осмолиттер көбейгеннен клетканың суды соруы су каналдары аркылы жүзеге асырылады.
Су каналдарының белсенділігі мембраналардың су өткізгіштік қабілетін реттеп тұрады. Осындай реттелу механизмдерінің бірі -аквапориндердің фосфорлану және дефосфорлану үдерістері. Фосфорланғанда су каналдары активтеленеді, дефосфорланғанда - белсендігі төмендейді.

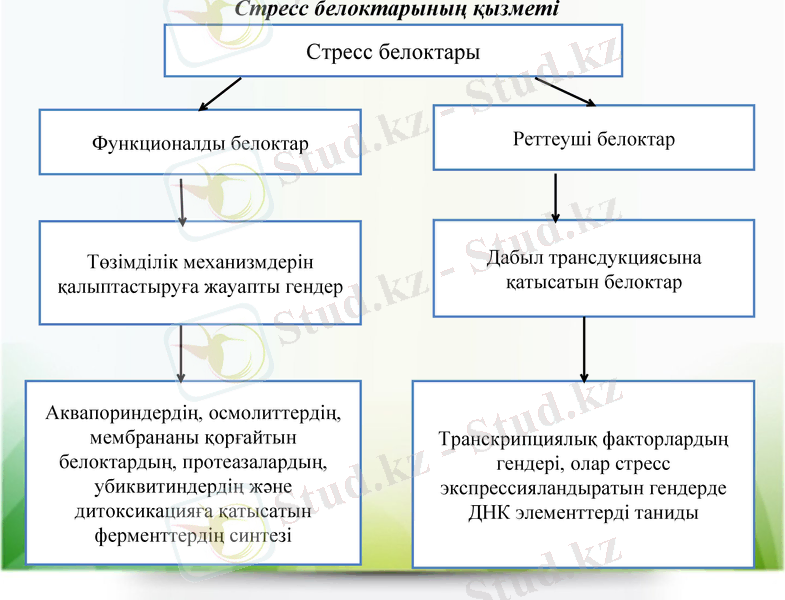
Стресс белоктардың қорғаныш және реттеу функциялары. Су тапшылығында пайда болған кейбір белоктар тұрақтылық механизміне, басқалары гендердің экспрессиясын реттеуге катысады. Соған карай су тапшылығында экспрессияланған гендер функционалды және реттеуші деп бөлінеді.
Бірінші топқа тұрақтылық механизмін қалыптастыруға жауапты гендер кіреді: аквапориндердің, протеазалардың, убиквитиндердің және детоксикацияга қатысатын ферменттердіц (СОД, аскорбат-ПО, глутатион-Б-трансфераза және т. б. ) синтезі.
Екінші топқа баска тұрақтылық механизмін калыптастыратын гендер экспрессияланғанда дабылға қатынасатын белоктардың гендері кіреді. Олар транскрипциялық факторлардың гендері, олар стресс экспрессияландыратын гендерде ДНҚ элементтерді таниды.
Стресс белоктары
Функционалды белоктар
Реттеуші белоктар
Төзімділік механизмдерін қалыптастыруға жауапты гендер
Дабыл трансдукциясына қатысатын белоктар
Аквапориндердің, осмолиттердің, мембрананы қорғайтын белоктардың, протеазалардың, убиквитиндердің және дитоксикацияға қатысатын ферменттердің синтезі
Транскрипциялық факторлардың гендері, олар стресс экспрессияландыратын гендерде ДНК элементтерді таниды
Стресс белоктарының қызметі

Гендер экспрессиясының реттелуі. Дабылдың рецепциясы. Су тапшылығында гендер экспрессиясы реттелуінің бірінші сатысы - дабылдың рецепциясы. Қабылданған дабыл трансдукция жүйесіне барады да, белоктарды белсендіреді. Олар дабыл тасымалдаушы тізбекті кұрады.

МАР-киназдық каскад. Ашытқыштарда фосфорланған белок Ssklp әртүрлі дабыл тасымалдаушы тізбектерді, мысалы, MAP (mitogen activated protein) - киназдық каскадты белсендендіреді. Олар үш бір- бірінен соң АТФ-пен фосфорланатын протеин-киназалардан: Ssk2p (МАРККК) - МАРКК-киназалар, Pbs2p (МАРКК) - МАРК-киназалар және Hoglp (МАРК) - МАР-киназалар тұрады. Ол үш протеинкиназа- дан тұрады.

Протеинкиназалар - белок молекуласында серин, треонин немесе тирозин қалдықтарына фосфатты топты кіргізуді катализдейді, ол белоктың фосфорланған түрін түзеді. Нәтижесінде молекуланың конформациясы өзгереді, белок белсенденеді немесе тежеледі.
Фосфорлану - белоктың белсенділігін бақылайтын өте қолайлы әдіс. Клеткаларда кең таралган.
Каскадтың соңында - белсенді киназа - MAPкиназа түзіледі. Бұл фермент фосфорландырады, сонымен бірнеше белок-нысанды белсендіреді. MAP-киназа фосфат қосылған соң, белсенді түрге айналады.

МАР-киназаны белсендіру үшін, клеткада ерекше киназа бола- ды - МАР киназаның киназасы /МАРКК/, ол тек МАР-киназаны фос- форландырады. Басында бұл фермент MAP-киназа сияқты ырықты емес. Оның өзі фосфат косылып белсенеді. Ол үшін клеткада басқа протеинкиназа болады: МАР-киназаның киназасыныц киназасы /МАРКК/. Ол бастапқы кезде белсенді емес. Оны сыртқы дабыл белсендіреді. МАРкиназдық каскадтың киназалары келесі жолмен белсенденеді:

Фосфорланғаннан кейін белсенденген MAP-киназа су тапшылығында және тұздық жағдайда ашытқыштардың көп гендерінің экспрессиясын, мысалы, глицериннің - ашытқыштардың ең маңызды осмолитінің - биосинтез ферментеріне жауапты гендерін индукцияландырады. Өсімдіктерде кейбір МАР-киназдық каскад компоненттерінің гомологтары табылған.
Дабыл қабылдау және тасымалдау деңгейлері:
- акпаратты сыртқы ортадан кабылдайтын тұтас өсімдік деңгейі;
- көп клеткалы организмнің ішіндегі клеткалардың қарым-катынас жасайтын деңгейі;
Олардың жүрісі клетка аралық әрекеттесуімен реттеледі: олар сыртқы мембрананың рецепторы арқылы дабыл қабылдайды.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz