Сүткоректілердің хромосомадан тыс экспрессиялаушы векторлары: құрылымы, селективті маркерлері және қолданылуы



Сүткоректілердің хромосомадан тыс экспрессиялаушы векторлары сүткоректілердің геномдарының функцияларын және реттелуін зерттеу үшін қолданылады. Сонымен қатар олардың көмегімен адамдардың кейбір сырқаттарын емдеу үшін қолданылатын аутентикалық белоктар алынуы мүмкін.
Сурет 7. 13. Сүткоректілердің экспрессиялаушы векторының жалпылама схемасы. Полилинкер (ПЛ) және селективті маркер эукариоттық промотор (р) және полиаденирлену сигналы (ро) . Бақылауында болады. Е. coli және сүткоректілер клеткасындағы векторлар репликациясы репликация репликациясы сайттарымен oriE и orieuk қамтамасыз етіледі. Трансформирленген E. coli клеткаларын таңдау үшін ампицилинге (Ampr) тұрақты ген қолданылады.

7. 13 суретте келтірілген жануар вирусының иниация репликация сайты. Клонданған және селективті маркерлі гендерлдің промоторлары, сонымен қатар олардың транскрипция терминациясының сигналдпары эукариот клеткаларында өтуі керек; көбіне жануар вирустарының ДНҚ - ң реттегіш кезектілігін қолданады. Мұнда күшті промоторлар және ұтымды полиаденирлену сигналдарын қолданған жөн.

Сүткоректілердің транфицирленген жасушаларын таңдау үшін, көбіне неомицинфосфотазаны кодтайтын Neor бактериальды генді қолданады. Бұл жүйеде генетицин деп аталатын токсинді қосылыс қолданылады, ол сүткоректілердің трансфицирленбеген клеткаларындағы трансляцияны шектейді. Сонымен қатар, трансфицирленген клеткаларда G - 418 фосфорланады және инактивтеледі. Сәйкесінше, Neor ген өнімін синтездейтін клеткалар тірі қалады және маманданады.
Сүткоректілердің трансфицерленген клеткаларын таңдаудың басқа жүйесі дигидрофолатредуктаза ферментін кодтайтын генді қолдануға негізделген. Бұл жүйеде DHFR дефекті гені бар клетканы қолданады, бұл клеткаларда DHFR функциясы синтезделмейді. Трансфекциядан кейін ортаға метотрексат қосады. Оның қатысында трансфицирленбеген клеткалар өспейді, ал синтездейтін клеткалар тірі қалады. DHFR-генді клеткаларды шамамен таңдап алғаннан кейін, метотрексат концентрациясын жоғарлатады және көп мөлшерде рекомбинантты белок синтездейтін вектор көшірмелерінің саны көп клеткаларды таңдап алады.
Селективті маркерлі гендер

Доминантты маркермен таңдап алудың басқада схемалары жасалған, мысалы, глутаминсинтетаза ферментін қолданумен, ол цитотоксинді әсеріне тұрақтылықты қамтамасыз етеді. Бұл жүйеде GS - генін тасымалдайтын вектор қолданылады. Оны сүткоректі клетка культурасына енгізеді және көп мөлшерде вектор көмірмесін тасымалдайтын клеткаларды даңдайды, ортада концентрациясын жоғарлатады. Мұнда қожайын клеткаларда GS болуы керек, себебі тек GS - генінің көп көшірмесінің болуы тұрақтылықты қамтамасыз етеді. Мұндай схеманың белгілі артықшылықтары бар.
Сүткоректілердің экспрессияланатын векторларына түрлі белоктардың гендері енгізілген және олардың қожайын клеткаларында экспрессиясы іске асқан. Кейде промотор және клонданған ген арасына интрон орналастырса, өнімнің шығуы жоғарлайды. Бұл феноминнің механизмі белгісіз.

Клонданған геннің экспрессиясын жоғарлату селективті маркерлі геннің экспрессиясын координациялау арқылы қол жеткізді. Бұл үшін DHFR генін клонданған генге жақын орналастырды, себебі екі генде бір промотордың бақылауында болуы қажет және полиоденилирленудің ортақ сигналы болуы тиіс. Ал DHFR гені интрон сплайсинг сайттарымен фланкирленген. DHFR және рекомбинантты белок біріншілік транскриптен трансляцияланды және сплайсирленген м РНҚ сәйкесінше (сур. 7. 14)

Сүткоректінің бір клеткасында клонданған екі ген экспрессиясы
Сурет. 7. 14 және рекомбинантты белоктың гендерінің координацияланған экспрессиясы. DHFR гені интронның донорлы және акцепторлы сплайсинг сайттарының арасына орналастырған, нысана геннің алдына (ген α) . DHFR және клонданған ген бір эукариоттық промотордың бақылауында және полиаденирленудің ортақ сигналы бар. DHFR сплайсирленбеген біріншілік транскриппен трансляцияланады, ал гетерологинді белок (белок α) сплайсингке түскен транскриптен. )
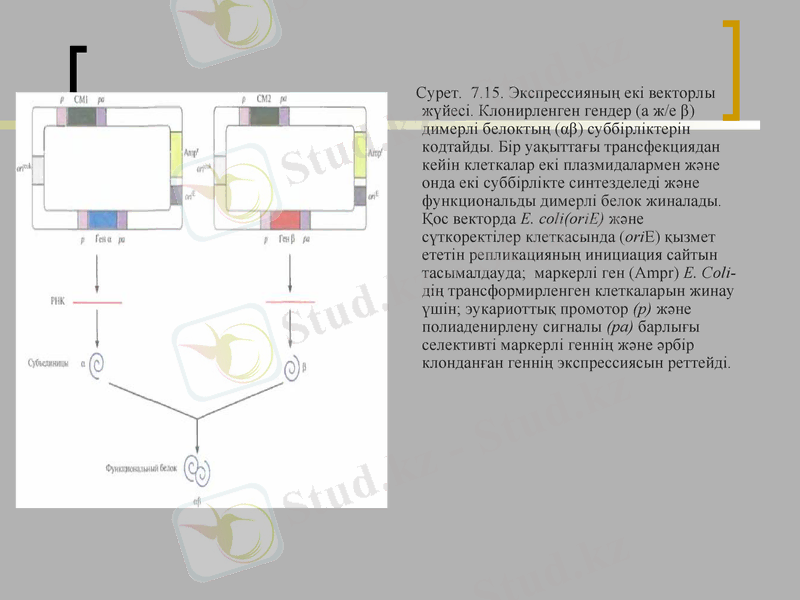
Сурет. 7. 15. Экспрессияның екі векторлы жүйесі. Клонирленген гендер (а ж/е β) димерлі белоктың (αβ) суббірліктерін кодтайды. Бір уақыттағы трансфекциядан кейін клеткалар екі плазмидалармен және онда екі суббірлікте синтезделеді және функциональды димерлі белок жиналады. Қос векторда E. coli(oriE) және сүткоректілер клеткасында (oriE) қызмет ететін репликацияның инициация сайтын тасымалдауда; маркерлі ген (Ampr) Е. Coli- дің трансформирленген клеткаларын жинау үшін; эукариоттық промотор (р) және полиаденирлену сигналы (ра) барлығы селективті маркерлі геннің және әрбір клонданған геннің экспрессиясын реттейді.

Димерлі және тетрамерлі белоктарды in vivo құрастыру ұтымды өтеді. Сондықтан бір клеткада екі түрлі рекомбинантты белокты синтездеу стратегиясы жасалды. Бұл үшін қожайын клетканы бір уақытта сүткоректінің екі экспрессирлеуші векторымен трансфицирледі, олардың әрқайсысы ген немесе к ДНК буббірліктің біріне тасымалдады және селективті маркерлердің гені (сур. 7. 15) . Трансфицерленген клеткаларды екі рет таңдауға ұшыратты, сәйкесінше тірі қалған клеткалар екі векторды да тасымалдады. Екі векторлы жүйелер аутентті димерлі және тетрамерлі рекомбинантты белоктарды синтездеу үшін қолданылады. Кейде екі рет трансфирленген клеткалар екі векторларының бірінен айырылады. Сонымен қатар, әрбір вектордың көшірмелерінің саны әр уақытта бірдей емес, сондықтан бір суббірлік екіншісімен салыстырғанда көп мөлшерде синтезделуі мүмкін және соңғы өнімнің шығымы азаюы мүмкін. Осы мәселені шешу үшін құрамында екі клонданған гені бар вектор құрастырылды. Рекомбинантты белоктордың бірдей мөлшердегі синтезіне қол жеткізу үшін екі ыдысты векторлар жасалды, онда клонданған гендер ДНқ сенменттеріне бөлінді, онда рибосомамен байланысудың ішкі сайты болды.

. Мұндай сайттар сүткоректілер вирустарының геномдарында табылған; Олар бір уақытта полицистронды мРНҚ - сы бар түрлі белоктардың бір уақыттағы трансляциясын қамтамасыз етеді. Конструкция транскрипциясы «ген - рибосомаларды байланыстырудың ішкі сайты - ген», бір промотормен және полиаденирлену сигналымен басқарылады. Бір транскрипті екі ген синтезделеді. Трансляция мРНҚ 5 ′ - соңынан және ішкі сайтттан басталады, нәтижесінде α және β димерлі белогінің суббірліктері синтезделеді. ( сур. 7. 17) .
Жинақтай келе, зерттеу және медициналық мақсаттарда аутентикалық рекомбинатты белоктар алу үшін, сүткоректілердің экспрессиялаушы векторлары әмбебап және ұтымды деп айта аламыз. Бірақта сүткоректілердің модификацияланған клеткаларын қолдану арқылы синтезделіп алынған рекомбинатты белоктар өте қымбат.
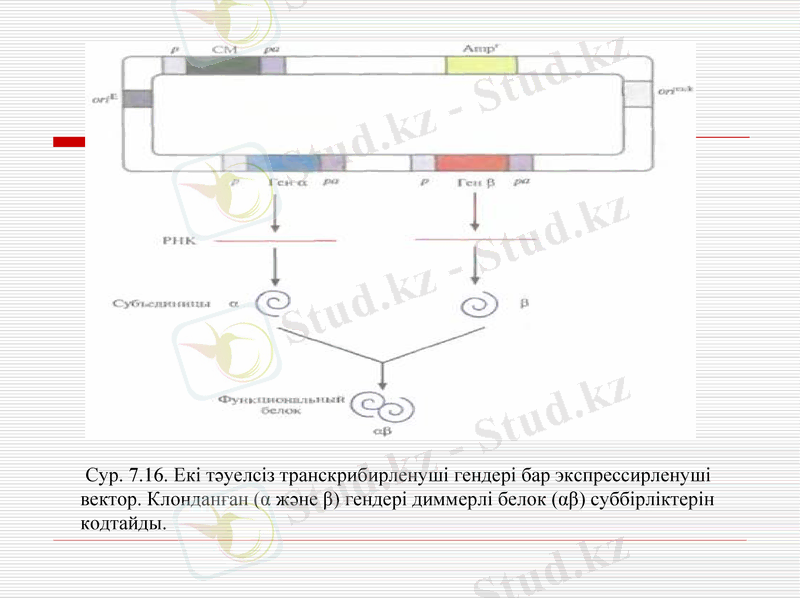
Сур. 7. 16. Екі тәуелсіз транскрибирленуші гендері бар экспрессирленуші вектор. Клонданған (α және β) гендері диммерлі белок (αβ) суббірліктерін кодтайды.
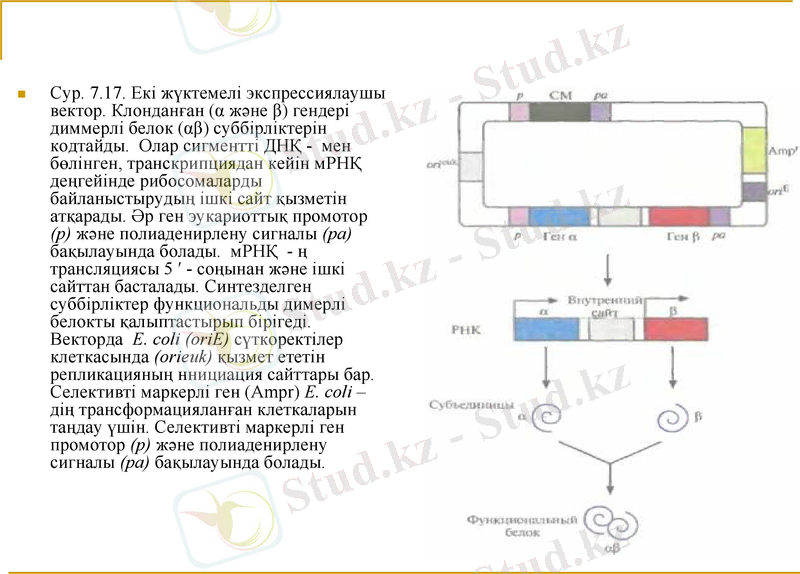
Сур. 7. 17. Екі жүктемелі экспрессиялаушы вектор. Клонданған (α және β) гендері диммерлі белок (αβ) суббірліктерін кодтайды. Олар сигментті ДНҚ - мен бөлінген, транскрипциядан кейін мРНҚ деңгейінде рибосомаларды байланыстырудың ішкі сайт қызметін атқарады. Әр ген эукариоттық промотор (р) және полиаденирлену сигналы (ра) бақылауында болады. мРНҚ - ң трансляциясы 5 ′ - соңынан және ішкі сайттан басталады. Синтезделген суббірліктер функциональды димерлі белокты қалыптастырып бірігеді. Векторда E. coli (oriE) сүткоректілер клеткасында (orieuk) қызмет ететін репликацияның ннициация сайттары бар. Селективті маркерлі ген (Ampr) Е. соli - дің трансформацияланған клеткаларын таңдау үшін. Селективті маркерлі ген промотор (р) және полиаденирлену сигналы (ра) бақылауында болады.

Прокариоттық экспрессия жүйелері көптеген белоктардың синтезі үшін жемісті қолданылады. Бірақта кейбір белоктар активті формаға өту үшін кейбір арнайыв посттрансляционды модификациядан - гликолиздену, фосфорирлену немесе ацетилденуден өтуі керек, ал бактериялар мұндайға қабілетсіз. Сондықтан клонданған гендерді эукариот клеткалардың көмегімен арнайы жасалған эукариоттты экспрессиялаушы векторлармен экспрессилауға бел буды.
Клонданған гендермен кодталатын түрлі белоктардың синтезі үшін S. Cerevisiae ашытқылары қолданылды. Олардың генетикасы жақсы зерттелген, сонымен қатар оларды оларды үлкен ферментерларда өсіруге болады. Белоктарды тазартуды жеңілдету үшін, олардың секрециясын қамтамасыз ететін векторлар құрастырылды. S. Cerevisiae ашытқылары көмегімен көптеген түрлі аутентикалық белоктар алынды. Бірақта бұл жүйедегі көптеген рекомбинатты белоктар посттрансляционды модификацияға ұшырамады, олардың шығымы жеткілікті жоғары болмады. Сондықтан рекомбинантты белоктардың синтезіне басқа ашытқы жүйелерді қолдану ұсынылды.

- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz