Қаңқа бұлшықетінің биомеханикасы және жиырылуын модельдеу


МАРАТ ОСПАНОВ АТЫНДАҒЫ БАТЫС ҚАЗАҚСТАН МЕМЛЕКЕТТІК МЕДИЦИНА УНИВЕРСИТЕТІ
Тәжірибелік сабақ №5
Тақырыбы: Бұлшық ет биомеханикасы. Бұлшық ет жиырылуын модельдеу.
Орындаған: Султанов Р. А
Тексерген: Турганбаева А. У.

Жоспар
І. Кіріспе
ІІ. Негізгі бөлім
а) Хилл теңдеуі. Бұлшық ет жиырылуын модельдеу.
б) Қаңқа бұлшық етінің биомеханикалық қасиеті
ІІІ. Қорытынды
ІV. Пайдаланылған әдебиеттер
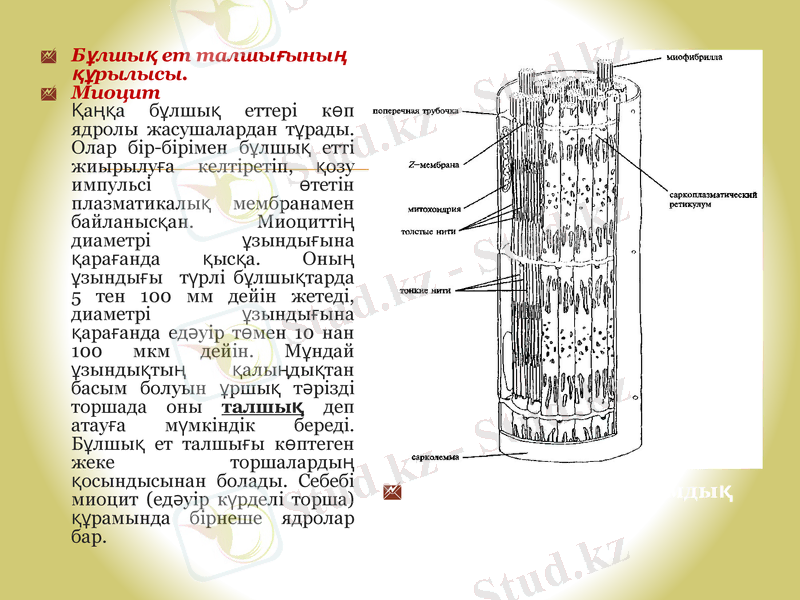
Бұлшық ет талшығының құрылысы.
Миоцит
Қаңқа бұлшық еттері көп ядролы жасушалардан тұрады. Олар бір-бірімен бұлшық етті жиырылуға келтіретіп, қозу импульсі өтетін плазматикалық мембранамен байланысқан. Миоциттің диаметрі ұзындығына қарағанда қысқа. Оның ұзындығы түрлі бұлшықтарда 5 тен 100 мм дейін жетеді, диаметрі ұзындығына қарағанда едәуір төмен 10 нан 100 мкм дейін. Мұндай ұзындықтың қалыңдықтан басым болуын ұршық тәрізді торшада оны талшық деп атауға мүмкіндік береді. Бұлшық ет талшығы көптеген жеке торшалардың қосындысынан болады. Себебі миоцит (едәуір күрделі торша) құрамында бірнеше ядролар бар.
Бұлшықтың құрылымдық схемасы

Саркомер құрылысы Миоцит шекараларының реттілігіне байланысты көрші миофибриллдердің саркомерлері беттеседі. Микроскоптан қарағанда, көлденең жолақты (қаңқалық) бұлшықтың белгілі бір реттілікпен орналасуы олардағы ашық немесе (изотропты-I) және қара (анизотропты-А) жолақтардың бір текті реттілігіне байланысты (сур. 2) . Анизотропты дисктер екілік сәуле сынумен қабілетті: қарапайым жарықта қараңғы болады, ал поляризацияланған жарықта-көлденеңінен мөлдір (ашық түсті), ұзынынан мөлдір емес.
Миофибриллалар
Миоцит құрамында (көлденең қимасында) 103 миофибрилл бар. Бұлшық ет жасушасы көптеген қысқару талшықтарынан тұрады. Олар - бір-біріне параллельді орналасқан миофибриллдер деп аталады (Сурет 1) . Қысқаруға бейім миофибриллдер диаметрі жуықтағанда 1мкм-ді құрайды. Миофибриллдердің бойымен саны көп саркомерлер орналасады ( грек. сөзінен sarx - ет, meros - бөлік ), бір-бірімен тізбек тәрізді жалғанады. Көрші саркомерлер бір-бірімен десмин ақуызы арқылы «желімденеді», өз кезегінде десмин актин жіпшелерімен актин-десминді торлы байланыс түзеді.

Саркомердің құрылысы: а-тігінен қимасы, б-жуан және жіңішке жіпшелердің қиылысындағы көлденең қимасы, в-жуан және жіңішке жіпшелердің қозғалысы нәтижесіндегі саркомер ұзындығының өзгеруі.
Екі F-актин жіпшелерінің арасындағы бұралған белсенді орталықтар миозинді бастармен байланысқа түсіп, бұлшық ет жиырылады. Актин, миозин және тропомиозин миоценттерінен басқа көлденең жолақты бұлшық еттің маңызды бағыттаушы ақуыз болып тропонин табылады. Ол актинді филамент пен тропомиозинді жіпшемен байланысады. Оның үш суббірлігі қосындыда молекулярлы массасы 76 кДа тең. Бір суббірлік 4 ион Са2+ байланысуы мүмкін мұндай әрекеттесу пайда болғанда, тропонин тропомиозинге әсер етіп, тропомиозин актинді филаментте миозинге белсенді орталықты босатады. Осылайша, олардың өзара әрекеттеріне кедергі жойылады

Бұлшық еттің жиырылуы.
Кальций иондары F- белсенді актинді орталықтарынан блокаданы бұзғаннан соң миозинді бастар олармен әрекеттеседі - олар саркомер орталығына қарай жылжиды. Миозин актинді жіпшеге бекітіліп, өз талшығының бойына иіледі (сур 4), актинді жіпше бойында миозинді жіпше бір қадамға ығысады (шамамен 10н. м-ге) . Актин өз кезегінде АТФаза ретінде миозиннің АТФ гидролиздеу қызметін белсендіреді. Демек, миозин молекуласының бастары жуан жіпшелер мен жіңішке жіпшелерді байланыстырып қоймай, АТФ гидролизіне қатысады. Актин жіпшесінің бойындағы миозин бастарының соңында миозин молекуласына жаңа АТФ молекуласы қосылады. Келесі АТФ гидролизінде миозин молекуласының алғашқы конформациясына қайтуы байқалады, бұл оның актин жіпшесіне қайта қосылуына мүмкіндік береді, бірақ алғашқы әрекеттесуге қарағанда саркомер орталыққа жақын орналасады.
Әрбір жуан филаментте 500 миозин бастарына жуық филамент құралады.
Бұлшық ет жиырылуы кезінде 1 секунд ішінде 5 циклдік секіріс жасайды (көлденең көпірлердің байлануы және босауы) . Көпірлер бірдей бір мезетте жұмыс істемейді: біреулері актин жіпшелерінде байланады, басқалары бұл мезгілде олардан бөлектенеді. М. В. Волькенштейн айтуы бойынша миозинді көпір 5*10-12 шартты қалыптастыруы керек. Мысалы: адам бицепсіне келетін бір аудан көлеміндегі барлық миозинді көпірлерлер 2*105 Па кернеуге сәйкес келеді, бұл бұлшық еттің (1, 8*105 Па) кернеуіне жақын келеді.
Бір ретті миозин басының актин филаментіне байлануы саркомердің алғашқы ұзындығын 1℅ қысқартады және шамамен 20℅ ПЭК-тің 3-5 пиконьютон күш жетілдіреді. Табиғи қалыпта бұлшық ет жиырылуы кезінде болатын көп ретті актин-миозин әсерлесулер әрбір саркомер барлық бұлшықтың қысқаруына пропорционалді қысқарады. Бұлшық еттің жиырылуы жуан және жіңішке жіпшелердің өздік ұзындықтарының өзгерісіне әкелмейді. Олар тек өз өлшемдері мен оларды құрайтын суббірліктердің сипаттамасын сақтап, бір-біріне қатысты бағыттарда жылжиды.
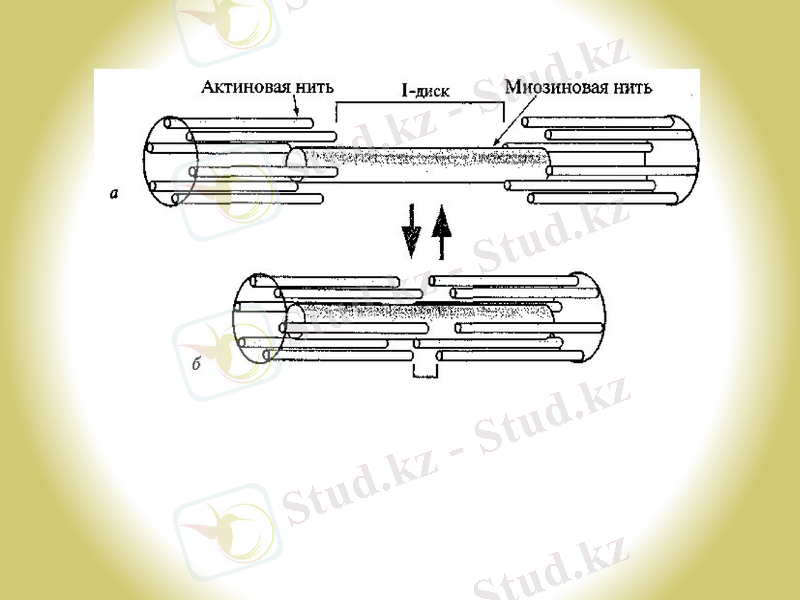
Актин жіпшесінің миозин жіпшелерімен қоршалуына байланысты жылжуы : а- босаңсу; 6 - жиырылу.

1950-жылдың басында Эндрю және Хью Хаксли, Р. Нидергерк пен Ж. Хэнсон, бұлшық етті рентгенқұрылымды талдау, оптикалық және электронды микроскоппен зерттеулер жүргізіп, бір-бірінен тәуелсіз «Жылжыған жіптер моделін ұсынды».
Жарты ғасыр ішінде бұлшық ет жиырылуының механизмдері туралы түсінік өзгеріске ұшыраған жоқ: бұлшық еттің максимальді жиырылу күші жуан және жіңішке жіпшелердің өзара жабу дәрежесіне пропорционалды болып келеді. Саркомер ұзындығы қысқарғанда тек I-дисктер ғана қысқарады, ал А-дисктері болса өз өлшемдерін өзгертпейді.
Саркомердің максимальді жиырылуы үшін миозин көпірлері шамамен 50 қағыс жасауы тиіс. Жылжу процессінің дискреттілігі жиырылу дәрежесінің мөлшерлігін қамтамасыз етеді. Жылжу процессінің дискреттілігі бір мезетте жұмылдырылған көлденең көпірлерге байланысты, ал жылдамдығы жұмылдандырылу жиілігіне тең. Қаңқа бұлшық етінің миоциттері үшін қысқару жылдамдығы 10-20 мкм*С-1-ге тең, ал кардиомиоциттер мәні бір реттілікке төмен. Айтылып кеткендей саркомер 3, 6-3, 8 мкм-ге дейін созылуы мүмкін. Сондықтан бұлшық етте ешқандай кернеу туындамайды. Бұлшық еттің қалыпты жиырылуы кезінде саркомер ұзындығы 2, 5 мкм жуық шамаға тең болады. Саркомер 2, 0-2, 2 мкм-ге қысқарғанда бұлшық ет максимальді күш жетілдіреді. Алайда оның ұзындығы 1, 5-1, 6 мкм дейін төмендеуі мүмкін. Бұл жағдайда бұлшық ет кернеуі бәсеңдейді, себебі миозинді филаменттер ұштары Z-жолаққа тіреледі (актин-десминді тор), сондықтан, миозин көпірлерінің қалпы актин филаменттеріне қатысты өзгеріп, күштің әлсіреуіне әкеледі.
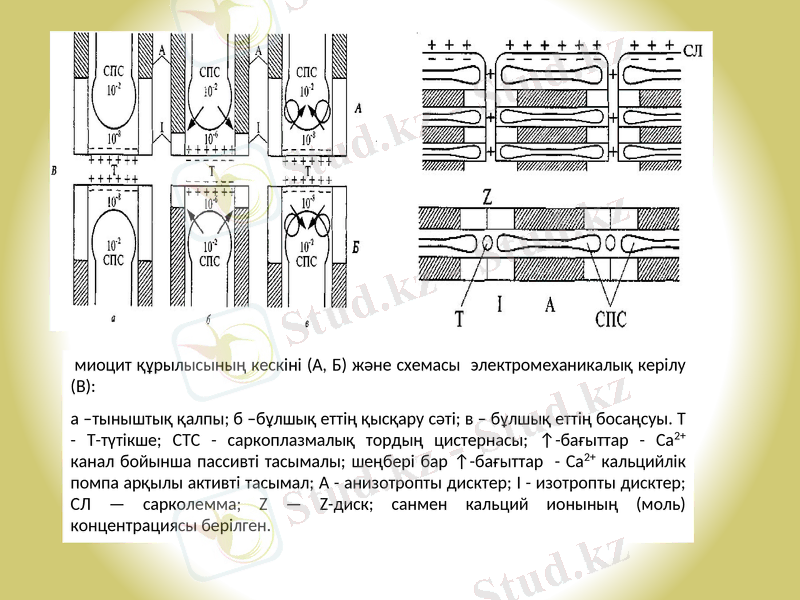
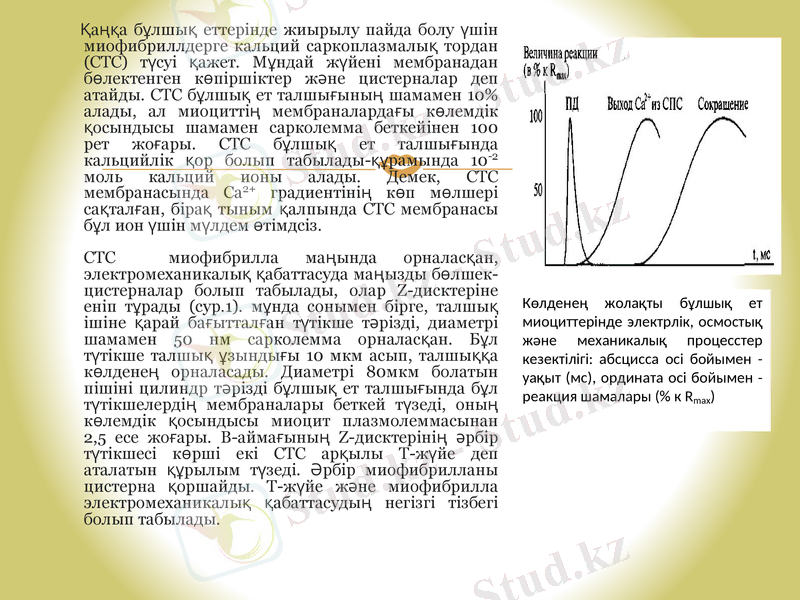
Қаңқа бұлшық еттерінде жиырылу пайда болу үшін миофибриллдерге кальций саркоплазмалық тордан (СТС) түсуі қажет. Мұндай жүйені мембранадан бөлектенген көпіршіктер және цистерналар деп атайды. СТС бұлшық ет талшығының шамамен 10% алады, ал миоциттің мембраналардағы көлемдік қосындысы шамамен сарколемма беткейінен 100 рет жоғары. СТС бұлшық ет талшығында кальцийлік қор болып табылады-құрамында 10-2 моль кальций ионы алады. Демек, СТС мембранасында Са2+ градиентінің көп мөлшері сақталған, бірақ тыным қалпында СТС мембранасы бұл ион үшін мүлдем өтімдсіз.
СТС миофибрилла маңында орналасқан, электромеханикалық қабаттасуда маңызды бөлшек-цистерналар болып табылады, олар Z-дисктеріне еніп тұрады (сур. 1) . мұнда сонымен бірге, талшық ішіне қарай бағытталған түтікше тәрізді, диаметрі шамамен 50 нм сарколемма орналасқан. Бұл түтікше талшық ұзындығы 10 мкм асып, талшыққа көлденең орналасады. Диаметрі 80мкм болатын пішіні цилиндр тәрізді бұлшық ет талшығында бұл түтікшелердің мембраналары беткей түзеді, оның көлемдік қосындысы миоцит плазмолеммасынан 2, 5 есе жоғары. В-аймағының Z-дисктерінің әрбір түтікшесі көрші екі СТС арқылы Т-жүйе деп аталатын құрылым түзеді. Әрбір миофибрилланы цистерна қоршайды. Т-жүйе және миофибрилла электромеханикалық қабаттасудың негізгі тізбегі болып табылады.

Электромеханикалық қабаттасу келесі процесстер кезектерінен тұрады: сарколемма бойымен ӘП таралуы → Т-жүйенің мембрана түтікшелерінің деполяризацияс → СТС мембранасында кальцийлік каналдардың ашылуы → СТС цистерналарынан Са2+ -тің бұлшық ет талшығының цитозоліне шығуы → миозинді көпірлердің тұйықталуы. Бұлшық еттің босаңсуы сарколемма реполяризациясы және СТС мембранасында жиналған кальцийлік түтіктің белсенген жұмысымен байланысты.
Электрлік, осмостық және механикалық процесстердің қаңқа бұлшық ет миоцитіндегі кезектігі (сур. 2) берілген.
Cинаптық қозудың жүйке талшығынан субсинаптық мембранаға берілгенде пластинка ұшының потенциалы (ПҰП) пайда болады, олар сарколеммадағы әрекет потенциалын генерациялайды. Оның амплитудасы 120-130 мВ, ал әрбір сарколемма нүктесінде 3 мс құрайды, бұл көрсеткіш сарколемма бойымен таралып Т-түтікше мембранасында мәні өзгеріп 3-5 м*с-1 жылдамдықты құрайды. Әрекет потенциалы орнағаннан соң шамамен 20 мс кейін СТС цистерналарынан максимальді Са2+ шығады, миозинді филаменттердің актин бойымен жылжуы нәтижесінен миоцит жиырылады. Бір ретті бұлшық ет талшығының қысқару уақыты 15 тен 50 мс дейін созылады содан соң миоцит босаңсиды. Бұлшық ет жиырылғанда сіңір аз қашықтыққа ауысады, бірақ едәуір қысыш күші дамиды.

Қаңқа бұлшық етінің биомеханикалық қасиеті.
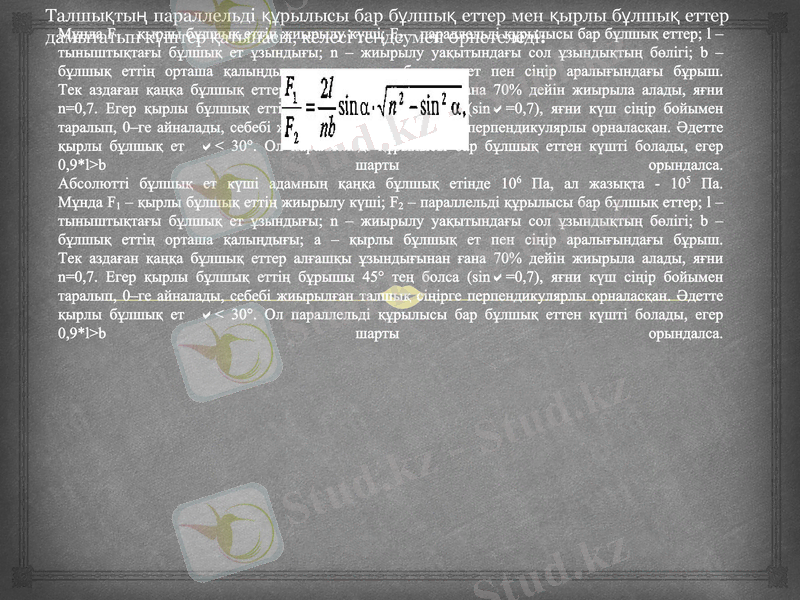
Талшықтың параллельді құрылысы бар бұлшық еттер мен қырлы бұлшық еттер дамытатын күштер қатынасы, келесі теңдеумен өрнетеледі:
Қаңқа бұлшық етінің биомеханикалық қасиеті.
Талшықтың параллельді құрылысы бар бұлшық еттер мен қырлы бұлшық еттер дамытатын күштер қатынасы, келесі теңдеумен өрнетеледі:
Мұнда F1 - қырлы бұлшық еттің жиырылу күші; F2 - параллельді құрылысы бар бұлшық еттер; l - тыныштықтағы бұлшық ет ұзындығы; n - жиырылу уақытындағы сол ұзындықтың бөлігі; b - бұлшық еттің орташа қалыңдығы; а - қырлы бұлшық ет пен сіңір аралығындағы бұрыш.
Тек аздаған қаңқа бұлшық еттер алғашқы ұзындығынан ғана 70% дейін жиырыла алады, яғни n=0, 7. Егер қырлы бұлшық еттің бұрышы 45° тең болса (sin=0, 7), яғни күш сіңір бойымен таралып, 0-ге айналады, себебі жиырылған талшық сіңірге перпендикулярлы орналасқан. Әдетте қырлы бұлшық ет < 30°. Ол параллельді құрылысы бар бұлшық еттен күшті болады, егер 0, 9*l>b шарты орындалса.
Абсолютті бұлшық ет күші адамның қаңқа бұлшық етінде 106 Па, ал жазықта - 105 Па.
Мұнда F1 - қырлы бұлшық еттің жиырылу күші; F2 - параллельді құрылысы бар бұлшық еттер; l - тыныштықтағы бұлшық ет ұзындығы; n - жиырылу уақытындағы сол ұзындықтың бөлігі; b - бұлшық еттің орташа қалыңдығы; а - қырлы бұлшық ет пен сіңір аралығындағы бұрыш.
Тек аздаған қаңқа бұлшық еттер алғашқы ұзындығынан ғана 70% дейін жиырыла алады, яғни n=0, 7. Егер қырлы бұлшық еттің бұрышы 45° тең болса (sin=0, 7), яғни күш сіңір бойымен таралып, 0-ге айналады, себебі жиырылған талшық сіңірге перпендикулярлы орналасқан. Әдетте қырлы бұлшық ет < 30°. Ол параллельді құрылысы бар бұлшық еттен күшті болады, егер 0, 9*l>b шарты орындалса.
Абсолютті бұлшық ет күші адамның қаңқа бұлшық етінде 106 Па, ал жазықта - 105 Па.
Мұнда F1 - қырлы бұлшық еттің жиырылу күші; F2 - параллельді құрылысы бар бұлшық еттер; l - тыныштықтағы бұлшық ет ұзындығы; n - жиырылу уақытындағы сол ұзындықтың бөлігі; b - бұлшық еттің орташа қалыңдығы; а - қырлы бұлшық ет пен сіңір аралығындағы бұрыш. Тек аздаған қаңқа бұлшық еттер алғашқы ұзындығынан ғана 70% дейін жиырыла алады, яғни n=0, 7. Егер қырлы бұлшық еттің бұрышы 45° тең болса (sin=0, 7), яғни күш сіңір бойымен таралып, 0-ге айналады, себебі жиырылған талшық сіңірге перпендикулярлы орналасқан. Әдетте қырлы бұлшық ет < 30°. Ол параллельді құрылысы бар бұлшық еттен күшті болады, егер 0, 9*l>b шарты орындалса. Абсолютті бұлшық ет күші адамның қаңқа бұлшық етінде 106 Па, ал жазықта - 105 Па. Мұнда F1 - қырлы бұлшық еттің жиырылу күші; F2 - параллельді құрылысы бар бұлшық еттер; l - тыныштықтағы бұлшық ет ұзындығы; n - жиырылу уақытындағы сол ұзындықтың бөлігі; b - бұлшық еттің орташа қалыңдығы; а - қырлы бұлшық ет пен сіңір аралығындағы бұрыш. Тек аздаған қаңқа бұлшық еттер алғашқы ұзындығынан ғана 70% дейін жиырыла алады, яғни n=0, 7. Егер қырлы бұлшық еттің бұрышы 45° тең болса (sin=0, 7), яғни күш сіңір бойымен таралып, 0-ге айналады, себебі жиырылған талшық сіңірге перпендикулярлы орналасқан. Әдетте қырлы бұлшық ет < 30°. Ол параллельді құрылысы бар бұлшық еттен күшті болады, егер 0, 9*l>b шарты орындалса.
Талшықтың параллельді құрылысы бар бұлшық еттер мен қырлы бұлшық еттер дамытатын күштер қатынасы, келесі теңдеумен өрнетеледі:

- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz