Транспозондар мен мобильді генетикалық элементтердің жіктелуі, қозғалыс механизмдері және биологиялық рөлі


Орындаған:Нигматиллаева С. С
Тобы:ЖМҚБ 01-19
Тексерген:Өтебай Д. М
ӨҢТҮСТІК ҚАЗАҚСТАН МЕДИЦИНА АКАДЕМИЯСЫ
ЮЖНО-КАЗАХСТАНСКАЯ МЕДИЦИНСКАЯ АКАДЕМИЯ
ПРЕЗЕНТАЦИЯ

ЖОСПАРЫ:
Кіріспе
Негізгі бөлім:
1. Жіктелуі
2. Транспозондар
3. ДНК-транспозондар
4. Ретротранспозондар
5. Инсерционды элементтер
6. Транспозондарды бұғаттау механизмі
7. Транспозондарды пайдалану
Қорытынды
Пайдаланылған әдебиеттер тізімі


Құрылыс және қозғалыс тәсілі бойынша:
- Транспозондар, мысалы, Tn5;
- Инсерциялық элементтер, мысалы, IS1603;
- ДНК-транспозондар;
- Ретротранспозондар;
- Плазмидтер, мысалы, ішек таяқшасының жыныстық факторы (F-плазмид) ;
- Бактериофагтар, мысалы, кездейсоқ геном учаскелеріне интеграцияланатын;

Транспозон-транспозиция деп аталатын процесс нәтижесінде геномның ішінде қозғалуға қабілетті ДНҚ тізбегі. Транспозондар-геномға қосылып, мутация тудыруы мүмкін, соның ішінде хромосомалық қайта құрулар сияқты маңызды геном элементтерінің мобильді элементтерінің бірі. Олар микроорганизмдер арасында дәрілік тұрақтылықты тасымалдау, рекомбинация және табиғатта (гендердің көлденең ауысуы), сондай-ақ гендік-инженерлік зерттеулер барысында әртүрлі түрлер арасында генетикалық материал алмасу процестерінде маңызды рөл атқарады.

Транспозондарды 1951 жылы Барбара Мак-Клинток ашты, оған 1983 жылы Нобель сыйлығы осы зерттеулері үшін берілді.
Транспозондар әдетте екі тік немесе инвертирленген қайталанатын ДНҚ тізбектерінен тұрады, олардың арасында ақуыз-кодтаушы гендер болады. Кейде транспозондардың орталық бөлігінің құрамында ұялы элементі бар организм үшін селективті артықшылықты қамтамасыз ететін гендер бар.

Барлық МАЭ жылжыту тәсілі бойынша екі негізгі сыныпқа топталуы мүмкін:
1-РНК-делдалдың ("copy and paste") көмегімен қозғалатын ретротранспозондар, және
2-ДНҚ транспозондар (тікелей ДНҚ қозғалады) .
Ретротранспозондар барлық эукариоттарда, саңырауқұлақтардан сүтқоректілерге дейін және бұл МГЭ жыныстық көбеюмен байланысты болады.
ДНК транспозондар-про-және эукариот арасында таралған МГЭ ең көне тобы.
Бір қызығы, бірінші класты МГЭ жалпақ және дөңгелек құрттарда табылған, ал екінші класты МГЭ паразиттік нематод үшін тән болды, бірақ трематод және таспалы құрттар үшін емес. Қазіргі уақытта сүтқоректілердің гельминттерінде ғана МГЭ 30-дан астам түрі сипатталған.

Оба класса делятся последовательно на подклассы, суперсемейства и семейства, согласно механизму их интеграции.
Ретротранспозондар саңырауқұлақтардан сүтқоректілерге дейінгі барлық эукариоттар табылған; олар
- LTR-ретротранспозондар,
- non-LTR ретротранспозондар және
- тирозин-рекомбинантты ретротранспозондар.
LTR ретротранспозондар бес топтарды біріктіреді: Copia, BEL, Gypsy, эндогенді ретровирустар (ERV) және DIRS.
non-LTR ретротранспозондар дербес LINES (дербес қозғалатын) және автоматты емес SINES (дербес МГЭ болуын өз орнын ауыстыру үшін талап ететін) элементтерді қамтиды.

Ретротранспозондар, шамасы, жыныстық көбеюмен байланысты, өйткені жануарлардың 24 түрі өкілдерінің арасында олар тек қана жыныссыз жолмен көбею арқылы коловраткалардан табылған жоқ.
Өзінің экспансиясы үшін ретротранспозондар алдымен қожайынның РНК полимеразасымен транскриптазамен көшіріліп, содан кейін өзінің кері транскриптазасымен көшіріліп отырады. Екі ферменттің де өңдеу қабілеті жоқ болғандықтан, бұл көшірмелер үрдісі бар

ДНК-ТРАНСПОЗОНДАР
ДНК-транспозондар транспозаза деп аталатын ферменттер кешенінің арқасында геном әдісімен "кесу және қою" қозғалады. Транспозаза ақуызының аминқышқыл тізбегі туралы ақпарат транспозон ретімен кодталған. Бұдан басқа, ДНҚ-ның бұл учаскесі, мысалы, гендер немесе олардың бөліктері транспозонмен байланысты басқа да тізбектерді қамтуы мүмкін. ДНҚ-транспозондардың көпшілігі толық тізбегі бар. Мұндай транспозондар дербес болып табылмайды және транспозаздың арқасында геноммен қозғалады, ол басқа, толық, ДНК-транспозонмен кодталған.
ДНК-транспозон учаскелерінің ұштарында транспоззаны танудың ерекше учаскелері болып табылатын инвертирленген қайталаулар орналасқан, осылайша геномның осы бөлігін басқалардан айырады. Транспозаза ДНҚ екі тізбекті кесінділерін жасауға, ДНҚ-нысана транспозонды кесуге және салуға қабілетті

Транспозондардың қозғалыс схемасы
I. ДНК-транспозондар: қозғалыс тәсілі "кесу және қою".
ІІ. ДДП-1-ретротранспозондар: қозғалыс тәсілі "көшіру және қою".

Инсерциялық бірізділік (IS, IS-элемент) - ДНҚ қысқа фрагменті, қарапайым ұялы генетикалық элемент.
Инсерциялық тізбектер екі маңызды сипаттамаға ие - олар басқа мобильді элементтерге (700-2500 нуклеотидтерге жуық) аз ұқсас және транспозиция процесіне тартылған ақуыздарды ғана кодтайды (транспозондарға қарағанда, сондай-ақ кейбір қосалқы гендерді кодтайтын, мысалы, антибиотиктерге резистенттілік гендерін) . Бұл белоктар әдетте IS элементіне ауысуға мүмкіндік беретін ферментативті реакцияны катализдейтін транспозазамен, сондай-ақ транспозиция белсенділігін ынталандыратын немесе тежейтін реттеуші белок бар.
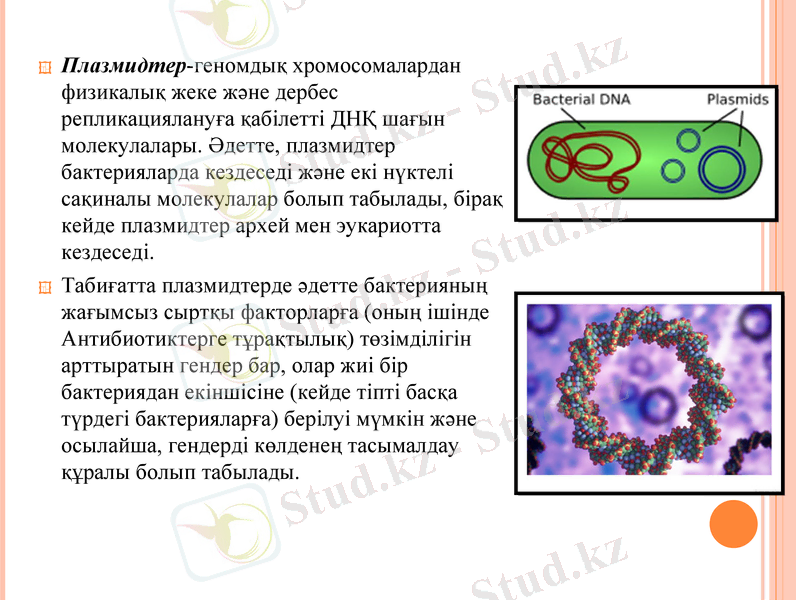
Плазмидтер-геномдық хромосомалардан физикалық жеке және дербес репликациялануға қабілетті ДНҚ шағын молекулалары. Әдетте, плазмидтер бактерияларда кездеседі және екі нүктелі сақиналы молекулалар болып табылады, бірақ кейде плазмидтер архей мен эукариотта кездеседі.
Табиғатта плазмидтерде әдетте бактерияның жағымсыз сыртқы факторларға (оның ішінде Антибиотиктерге тұрақтылық) төзімділігін арттыратын гендер бар, олар жиі бір бактериядан екіншісіне (кейде тіпті басқа түрдегі бактерияларға) берілуі мүмкін және осылайша, гендерді көлденең тасымалдау құралы болып табылады.

Жасушадағы плазмид функциялары өте әртүрлі. Оларға жатады:
конъюгация кезінде генетикалық материалды тасымалдау-F-плазмид;
бактериоциногендігі плазмидтері басқа бактериялар үшін өлім - жітім ақуыздарының синтезін бақылайды-Col-плазмидтер;
гемолизиндердің синтезі-Hly-плазмидтер (конъюгативті болып табылады) ;
ауыр металдарға төзімділік;
Антибиотиктерге тұрақтылық (R-плазмидтер) ;
энтеротоксиндер синтезі-Ent-плазмидтер;
УК-сәулеленуге төзімділік;
адам және жануарлар ағзасындағы жасушалардағы бактериялардың адгезиясын қамтамасыз ететін антигендердің синтезі - отарлау антигендерінің плазмидтері;
рестрикция-түрлендіру жүйесі;
расщепление камфора (плазмида ӨЗІ), ксилол (плазмида XYL), салицилаты (плазмида SAL) (табылған кейбір штаммдар Pseudomonas)
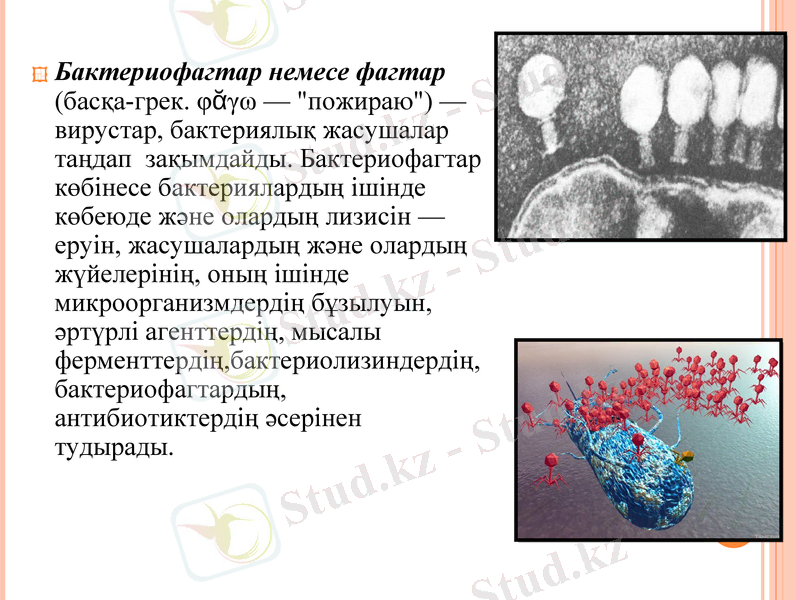
Бактериофагтар немесе фагтар (басқа-грек. φᾰγω - "пожираю") - вирустар, бактериялық жасушалар таңдап зақымдайды. Бактериофагтар көбінесе бактериялардың ішінде көбеюде және олардың лизисін - еруін, жасушалардың және олардың жүйелерінің, оның ішінде микроорганизмдердің бұзылуын, әртүрлі агенттердің, мысалы ферменттердің, бактериолизиндердің, бактериофагтардың, антибиотиктердің әсерінен тудырады.

МОБИЛЬДІ ЭЛЕМЕНТТЕР ТУДЫРАТЫН ӘСЕРЛЕР
- Ген ішіне мобильді элементтерді енгізу генді өшіруге әкеледі.
- Егер мобильді элемент оператор мен цистрон арасында енгізілсе, геннің реттелуі бұзылуы мүмкін (мобильді элементте өзінің промоторы бар) .
- мобильді элементті кірістіру гендер экспрессиясына әкелуі мүмкін, олар қазіргі уақытта жұмыс істемеуі тиіс.
Мобильді элементтердің болуы заңсыз рекомбинацияға ықпал ететін фактор болып табылады.
Заңсыз рекомбинация кезінде бір-біріне қатысы жоқ гендер сорылады.
мобильді элементтер делеция, инверсия, дупликация шақырады. Мұның бәрі-хромосомдық мутацияларға әкеледі.

ТРАНСПОЗОНДАРДЫ БҰҒАТТАУ МЕХАНИЗМДЕРІ
Геномның мобильді элементтері өсімдік және жануарлар геномдарында кеңінен ұсынылған. Олардың жоғары белсенділігі геномның тұрақтылығы үшін тәуекел болып табылады, сондықтан олардың экспрессиясы, әсіресе гаметаларды қалыптастыруға және ұрпақтарға тұқым қуалайтын ақпаратты беруге қатысатын ұлпаларда қатаң реттеледі. Өсімдіктер мен жануарларда геномның мобильді элементтерінің белсенділігін реттеу ДНҚ реттілігі мен ақуыз Комплекстерімен Аргонавт бірге түзілмейтін РНҚ белсенділігінің de novo метилденуі арқылы жүзеге асырылады

пиРНК-транспозондарды индукцияланған басу механизмі мүлдем анықталмаған, бірақ оны схемалық түрде мұндай модельмен ұсынуға болады:
РНК, пиРНК-прекурсорлардың Бір тізбекті молекулаларының бастапқы жиналуы;
пиРНК пісуі және оларды пиви-ақуыздарының көмегімен амплификациялау (пинг-понг циклы) ;
мақсатты транспозонның басылуы, бұл бірнеше жолмен болуы мүмкін: РНК тозуы (ақуыз-аргонавттардың h-ұқсас доменінің Рнказдық белсенділігінің көмегімен), трансляцияның басылуы және хроматин-түрлендіруші жүйелерді тарту (SWI/SNF ақуыздар сияқты) және транспозонның одан әрі эпигенетикалық басылуы.
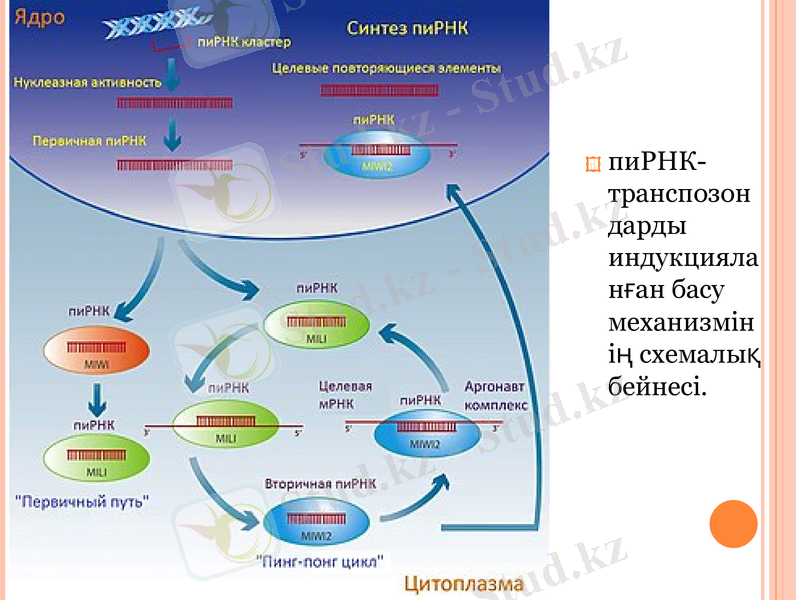
пиРНК-транспозондарды индукцияланған басу механизмінің схемалық бейнесі.

Көбею үшін иесі ағзасын пайдаланатын және оны тастап кетуі мүмкін вирустардан айырмашылығы, мобильді генетикалық элементтер тек қана иесі ағзасында бар. Сондықтан транспозондар өз белсенділігін реттеуге қабілетті. Бұл мысал-AC ДНК-транспозондар-дербес мобильді өсімдіктер элементтері. Ac-элементтер транспозаның белсенділігін оның көшірмелерін көбейту кезінде төмендету қабілетін көрсетеді.
Сондай-ақ, MuDR өсімдік автономды ДНК-транспозондарын басу Muk арқылы болуы мүмкін. Muk mudr нұсқасы болып табылады және ДНҚ бірнеше палиндромдық учаскелері бар. Muk транскрипцияланған кезде, мұндай РНК шпильканы қалыптастырады, содан кейін РНК-интерференция процесінің көмегімен MuDR белсенділігін тежейтін шағын интерферациялаушы РНК (миРНК) ферменттер кешенімен кесіледі

АУРУЛАР
2012 жылғы жағдай бойынша адамның 96 түрлі ауруы құжатталған, оның себебі de novo мобильді генетикалық элементтерді енгізу болып табылады. Alu-қайталаулар жиі хромосомдық аберрация туғызады және 50 түрлі аурулардың себебі болып табылады. Мәселен, I типті нейрофиброматозда 18 ретротранспозон жағдайлары табылды, оның 6-ы 3 арнайы жерде орын алады. Соматикалық тіндердегі ДДП-1 мобильдік элементтерінің белсенділігі өкпе обыры бар емделушілерде тіркелген.
Егер ауру тудыратын транспозиция гаметаларда болса, онда келесі ұрпақ ауруларды зерттейді. Осылайша, гемофилия ДДП-1 ретротранспозонының ДНК учаскесіне қосылуынан пайда болуы мүмкін, ол қанның ұюының VIII факторының генін кодтайды. Тінтуірде онкогенез, дамудың тоқтауы және геномның мобильді элементтерінің кіруіне байланысты стерильдік жағдайлары тіркелді

ТРАНСПОЗОНДАРДЫ ПАЙДАЛАНУ
Гендік инженерия
Геномның мобильді элементтері хроматинаға қосылуға қабілетті болғандықтан, олар гендік инженерияда гендерді немесе ДНҚ учаскелерін арнайы және бақыланатын біріктіру үшін пайдаланылады. Транспозондар мутагенез үшін және зертханаларда геномның реттеуші элементтерін анықтау үшін қолданылады.
In vivo - P енгізілген мутагенездің ең танымал жүйесі-шыбынның d. melanogaster мобильді элементі, оның көмегімен гендердің функцияларын зерттеуге, хромосомдық аберрацияларды жолға қоюға және т. б.

Омыртқалы жануарларда ұзақ уақыт геномның транспозондық модификациясының тиімді әдісі болған жоқ. Қазір tol2 мобильді элемент жүйесі бар, ол жапондық Oryzias latipes балығынан алынған. Сондай-ақ, Minos транспозондар жүйесі табысты болып табылады.
"Ұйқыдағы ару" транспозондар жүйесі (ағылш. Sleeping Beauty) Балықтан ДНК транспозасының реттілігі негізінде құрылды. Бұл жүйені сәтті пайдалану адам ішек обырының онкогеніне кандидаттарды анықтауға мүмкіндік берді.

ФИЛОГЕНЕТИКА
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz