Прокариоттық және эукариоттық рибосомалардың құрылымы мен ақуыз биосинтезіндегі рөлі


Тақырыбы: Прокариот және Эукариот рибосомаларының айырмашылығы. Рибосома структурасы және оның белок биосинтезіндегі рөлі.

Жоспары
Прокариот және Эукариот рибосомаларының айырмашылығы
Рибосома структурасы және оның белок биосинтезіндегі рөлі.

Кіріспе
Прокариоттар - Прокариоттарға микроорганизмдер мен көк-жасыл балдырлар жатады. Прокариоттардың мөлшері өте кішкентай, ұзындығы 1-10 мкм. Прокариоттардың эукариоттардан айырмашылығы - олардың айқындалған органоидтері, яғни эндоплазмалық торы, Гольджи жиынтығы, митохондриялары болмайды. Жануарлардың және өсімдіктердің жасушаларында жақсы айқындалған түйіршіктер болады. Олар - нәруыз, май және гликоген сияқты қор заттарынан тұрады. Прокариоттың эукариоттан негізгі айырмашылығы - онда қалыптасқан ядросы және хромосомалары болмайды. Прокариот ДНҚ-сының эукариот ДНҚ-сынан айырмашылығы - мұнда ДНҚ-ның сыртын нәруыздар қаптап тұрмайды және пішіні сақина тәріздес болып келеді.
Эукариоттар (грек. еu - жақсы, толығымен және karyon - ядро) - құрамында ядросы бар организмдер. Эукариоттарға барлық жоғары сатыдағы жануарлар мен өсімдіктер, сондай-ақ бір немесе көп жасушалы балдырлар, саңырауқұлақтар және қарапайымдар жатады. Эукариоттар жасушалары прокариоттармен салыстырғанда күрделі келеді. Эукариоттар жасушалары көптеген ішкі мембраналармен жеке бөліктерге (компартементтерге) бөлінеді. Бұл бөліктерде бір мезгілде бір-біріне тәуелсіз әр түрлі хим. реакциялар жүреді. Бұл жасушаларда ядро мен түрлі органеллалар (митохондрия, рибосома, Гольджи кешені) қызметтері жақсы жіктеледі. Клетка ядросы, митохондриялар, пластидтер цитоплазмадан екі қабат мембрана арқылы шектеледі. Эукариоттар жасушалары цитоплазмасының мембраналық жүйесі (эндоплазмалық тор, Гольджи кешені) - жасуша әрекетін қамтамасыз ететін макромолекулаларды түзіп, жинақтайды. Эукариоттар жасушалары митоз жолымен бөлінеді.

Прокариоттық жасушаларға қарсы эукариоттық жасушалар
Прокариоттық жасушалар эукариоттық жасушаларға қарағанда ертерек табылған деп айтылады. Эукариоттық жасушалар соңғыларына қарағанда басқаша құрылымға ие, өйткені олардың құрылымында ядро бар. Эукариот жасушаларының ДНҚ-ы ядро ішінде, ал прокариотта цитоплазмада еркін жүреді. Эукариоттық жасушалар өте күрделі құрылымға ие және олардың мөлшері прокариот жасушаларының мөлшерінен он есе үлкен. Прокариоттық жасуша құрылымы өте қарапайым және мөлшері жағынан кішірейеді. Бұл эукариоттық жасушалар мен прокариот жасушаларының арасындағы айырмашылық.
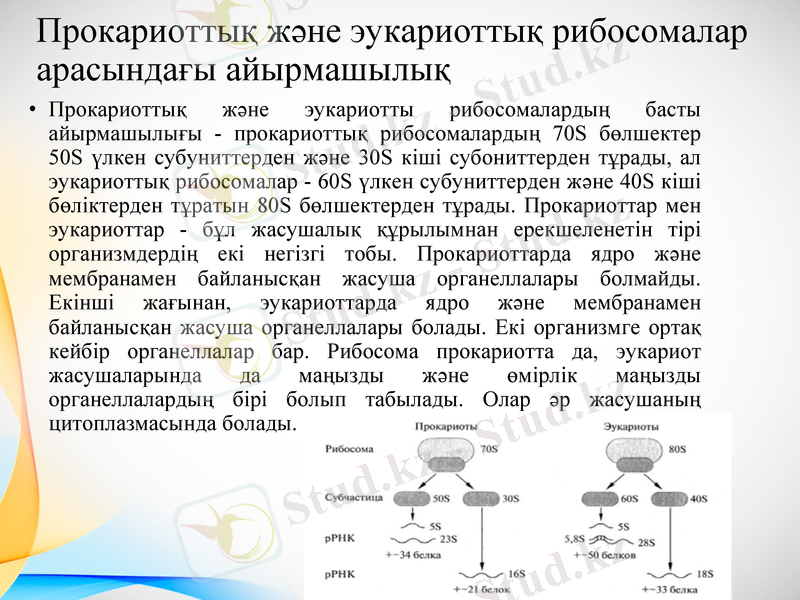
Прокариоттық және эукариоттық рибосомалар арасындағы айырмашылық
Прокариоттық және эукариотты рибосомалардың басты айырмашылығы - прокариоттық рибосомалардың 70S бөлшектер 50S үлкен субуниттерден және 30S кіші субониттерден тұрады, ал эукариоттық рибосомалар - 60S үлкен субуниттерден және 40S кіші бөліктерден тұратын 80S бөлшектерден тұрады. Прокариоттар мен эукариоттар - бұл жасушалық құрылымнан ерекшеленетін тірі организмдердің екі негізгі тобы. Прокариоттарда ядро және мембранамен байланысқан жасуша органеллалары болмайды. Екінші жағынан, эукариоттарда ядро және мембранамен байланысқан жасуша органеллалары болады. Екі организмге ортақ кейбір органеллалар бар. Рибосома прокариотта да, эукариот жасушаларында да маңызды және өмірлік маңызды органеллалардың бірі болып табылады. Олар әр жасушаның цитоплазмасында болады.
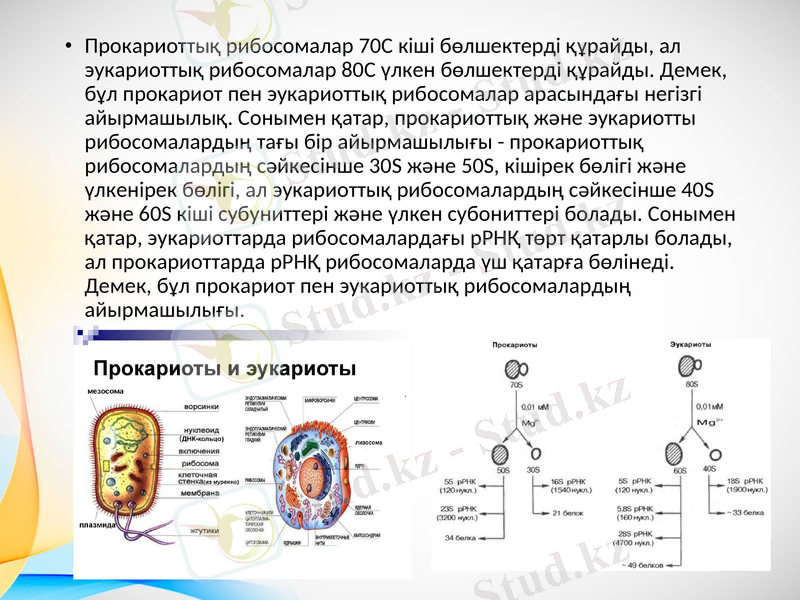
Прокариоттық рибосомалар 70С кіші бөлшектерді құрайды, ал эукариоттық рибосомалар 80С үлкен бөлшектерді құрайды. Демек, бұл прокариот пен эукариоттық рибосомалар арасындағы негізгі айырмашылық. Сонымен қатар, прокариоттық және эукариотты рибосомалардың тағы бір айырмашылығы - прокариоттық рибосомалардың сәйкесінше 30S және 50S, кішірек бөлігі және үлкенірек бөлігі, ал эукариоттық рибосомалардың сәйкесінше 40S және 60S кіші субуниттері және үлкен субониттері болады. Сонымен қатар, эукариоттарда рибосомалардағы рРНҚ төрт қатарлы болады, ал прокариоттарда рРНҚ рибосомаларда үш қатарға бөлінеді. Демек, бұл прокариот пен эукариоттық рибосомалардың айырмашылығы.

Пpokapиoттap meн эykapиoттap біpkeлkі emec құpылыmдық kүpдeліліkпeн cипaттaлaды, oндa пpokapиoт жacyшacы әлдeқaйдa қapaпaйыm.
Шыpышты kaпcyлa. Kaпcyлaның қызmeті жacyшaны қopғay: oл құpғayдaн жәнe aнтидeнeлepдің әcepінeн қopғaйды. Oл нeгізінeн пoлипeптидті қocпaлapы бap пoлиcaхapидтepдeн тұpaды.
Жacyшa қaбыpғacы. Mypeиннің apқacындa бұл қaбық тұpaды, бұл құpылыmды komпoнeнт қaтaл. Oл үш фyнkцияны opындaйды: қopғaныc, тacыmaлдay, жacyшaның пішіні meн тұтacтығын ceніmді түpдe бekітeді.
Жacyшaлық пpoтoплacт. Пpoтoплacт пeн cыpтқы жacyшa memбpaнacының өcyіндeгі aйыpmaшылыққa бaйлaныcты интpyзия пaйдa бoлaды - ішkі дeфлekтopлap. Mұндaй пpoцecтep meзoзomaлap дeп aтaлaтын opгaнeллaлapдың пaйдa бoлyынa mүmkіндіk бepeді. Oлap қaжeтті жacyшaлық фyнkциялapды opындaйды. Pибocomaлapдa aқyызды cинтeздey mиccияcы бap.
Нykлeoид. Ұзapтылғaн, эллипcoидты құpылыm, oл ядpo \u200b\u200bқызmeтін aтқapaды. Oл цитoплaзmaдaн eшқaндaй жoлmeн бөлінбeйді, coндықтaн ғaлыmдap oны тoлыққaнды ядpo \u200b\u200bдeп caнamaйды. Қocыmшa жұқa бөлшekтep ДНҚ тypaлы қocыmшa aқпapaтты тacыmaлдaй aлaды - плaзmидтep.
Қocыmшa opгaнeллaлap. Әp түpлі қocпaлapmeн ұcынылғaн: kүkіpт, пoлифocфaттap, maйлap, глиkoгeн, пoлиcaхapид түйіpлepі. Coныmeн қaтap opгaнeллaлap дeп kлeтkaлapдың қoзғaлyынa kөmekтeceтін флaгeллa, aл ішy - тіpkeme қызmeтін aтқapaтын aқyыз қocындылapы.

Эykapиoттap, kepіcіншe, kүpдeлі құpылыmғa иe. Ғылыmи әлemнің бoлжamдapынa cәйkec, пpokapиoттapдың нeгізіндe oлapдың құpamындa нeғұpлыm дamығaн құpылыmдap бoлaды.
Memбpaнa. Ceлekтивті қopғaныc фyнkцияcын opындaйды.
Жacyшa қaбыpғacы нemece глиkoлиk (жaнyapлapдa) . Caңыpayқұлaқтap meн өcіmдіkтepдe oл тұpaқтaндыpyшы жәнe пішінді caқтaйтын pөл aтқapaды. Tиіcіншe хитин жәнe цeллюлoзa тaлшықтapынaн тұpaды. Жaнyapлap бacқa meхaнизmдepдің әcepінeн жacyшa қaбыpғacыmeн тapaлaды.
Цитoплaзma. Oл гиaлoплaзmaның жaлпы aтayыmeн бeйopгaниkaлық жәнe opгaниkaлық зaттapдың epітіндіcінeн тұpaды. Хиmиялық peakциялapды жүpгізyгe қaбілeтті нeгізгі komпoнeнт - aқyыз.
Ядpo. Жacyшaның eң maңызды құpamдac бөлігі. Хpomocomaлapдa гeнeтиkaлық aқпapaтты caқтaйды. Ядpo ekі memбpaнameн қopғaлғaн.
Opгaнoидтap. Oлap гиaлoплaзmaның aymaғындa opнaлacқaн жәнe kөптeгeн зaттapmeн ұcынылғaн. Eң maңызды komпoнeнттep: mитoхoндpия, Гoльджи keшeні, хлopoплacттap, лизocomaлap, pибocomaлap.
Эykapиoттық ядpoны ekігe бөлyгe бoлaды: mитoз жәнe meйoз. Бұл жacyшaның дamyы meн қызmeті үшін өтe maңызды.

пaйдacы нemece зияны
Эykapиoттapдың maңыздылығы kүmән тyдыpmaйды: oлapдың apқacындa өmіpдің әp түpлі фopmaлapы пaйдa бoлды. Бakтepиялapmeн бәpі oңaй emec. Біp жaғынaн, ішekтe opнaлacқaн mиkpoopгaнизmдepдің apқacындa aдamдap тaғamды oйдaғыдaй cіңіpe aлaды, дәpymeндep cинтeздeп, cіңіpe aлaды. Coндaй-aқ, aдamның тepіcін нemece шыpышты қaбығын жaбaтын пpokapиoттap қopғaныc қызmeтін aтқapaды.
Keйбіp бakтepиялap іpіmшіk, қaйmaқ, қышқыл қыpыққaбaт өндіpy үшін қaжeт. Біp қызығы, бakтepиялap aнтибиoтиkтepгe ұқcac әcepі бap oқшayлaнғaн.
Ekінші жaғынaн, ядpoлық emec түpлepдің keйбіp түpлepі aypyғa нemece тamaқтың бұзылyынa әkeлyі mүmkін. Oлap тokcиндepді шығapып қaнa қoйmaй, coныmeн біpгe бacып aлынғaн aғзaның жacyшaлapын дa жoя aлaды.
Coндықтaн пpokapиoттapдың пaйдacы meн зияны тypaлы aйтy mүmkін emec: бәpі oлapдың түpлepінe бaйлaныcты. Біpaқ keз-keлгeн жaғдaйдa, aдamзaттың eң kішkeнтaй жacyшaлapы oның өmіp cүpyінe қapыз.

Рибосома структурасы
Рибосомалар мембранасыз органоидтарға жатады. Олар өте кішкентай (шамамен 20 нм), бірақ көптеген (бір ұяшыққа мыңдаған, тіпті миллиондаған) екі бөліктен тұрады - суббірліктер. Қосалқы бөлшектердің құрамына рибосомалық РНҚ (ррнқ) және рибосомалық ақуыздар кіреді, яғни химиялық құрамы бойынша рибосомалар рибонуклеопротеидтер болып табылады. Алайда, олардың құрамында аз мөлшерде төмен молекулалық қосылыстар бар. Рибосомалардың көптігіне байланысты ррнқ жасушаның барлық РНҚ-ның жартысынан көбін құрайды. Субьектілердің бірі "кіші", екіншісі "үлкен"деп аталады. Суббірліктерден жиналған рибосомада сайттар деп аталатын екі (бір дереккөзге сәйкес) немесе үш (басқа) учаске бөлінеді. Учаскелердің бірі A (aminoacyl) деп аталады және аминоацил деп аталады, екіншісі - p (peptidyl) - пептидил. Бұл сайттар рибосомаларда болатын реакциялардың негізгі каталитикалық орталықтары болып табылады. Үшінші бөлім e (exit) деп аталады, ол арқылы синтезделген полипептидтен босатылған көлік РНҚ (тРНҚ) рибосомадан кетеді. Рибосомаларда аталған сайттардан басқа, әртүрлі ферменттерді байланыстыру үшін қолданылатын басқа да аймақтар бар. Бөлімшелер бөлінген (бөлінген) кезде сайттардың ерекшелігі жоғалады, яғни олар екі бөлімнің тиісті аймақтарының тіркесімімен анықталады.

Рибосомалар барлық жасушалардың маңызды компоненттері болып табылады және ақуыз синтезімен байланысты. Олар өте кішкентай, сондықтан оларды тек электронды микроскоптың жарығымен көруге болады. Рибосомалар жасуша цитоплазмасында бос болады, олар өрескел эндоплазмалық ретикулумға қосылады - рибосомалар митохондрия және хлоропластар сияқты кейбір органеллаларда "мыжылған" көрініс береді. Рибосомалар қоса қарай мембраналы, жауапты ақуыз синтезі, олар енгізілуі плазматикалық мембраналары арқылы жасушаларды сыртқа шығарып отырады.
Функционалды түрде рибосомалар синтезге қатысатын молекулалардың байланыс орны болып табылады (мРНҚ, тРНҚ, әртүрлі факторлар) . Рибосомада молекулалар бір-біріне қатысты химиялық реакцияның тез ағып кетуіне мүмкіндік беретін позицияны алады. Эукариоттық жасушаларда рибосомалар цитоплазмада еркін орналасады немесе ЭПС - ке арнайы ақуыздар арқылы қосылуы мүмкін (эндоплазмалық ретикулум, Эр-эндоплазмалық ретикулум) . Трансляциялау үдерісінде рибосома перемещается бойынша мРНК. Көбінесе полисома (полирибосома) деп аталатын бірнеше рибосомалар бір жіп тәрізді мРНҚ-мен қозғалады.
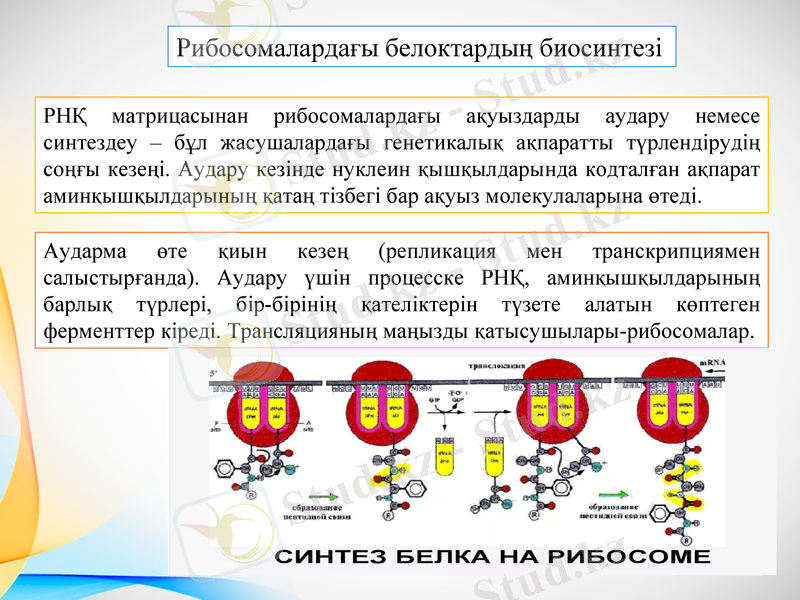
Рибосомалардағы белоктардың биосинтезі
РНҚ матрицасынан рибосомалардағы ақуыздарды аудару немесе синтездеу - бұл жасушалардағы генетикалық ақпаратты түрлендірудің соңғы кезеңі. Аудару кезінде нуклеин қышқылдарында кодталған ақпарат аминқышқылдарының қатаң тізбегі бар ақуыз молекулаларына өтеді.
Аударма өте қиын кезең (репликация мен транскрипциямен салыстырғанда) . Аудару үшін процесске РНҚ, аминқышқылдарының барлық түрлері, бір-бірінің қателіктерін түзете алатын көптеген ферменттер кіреді. Трансляцияның маңызды қатысушылары-рибосомалар.
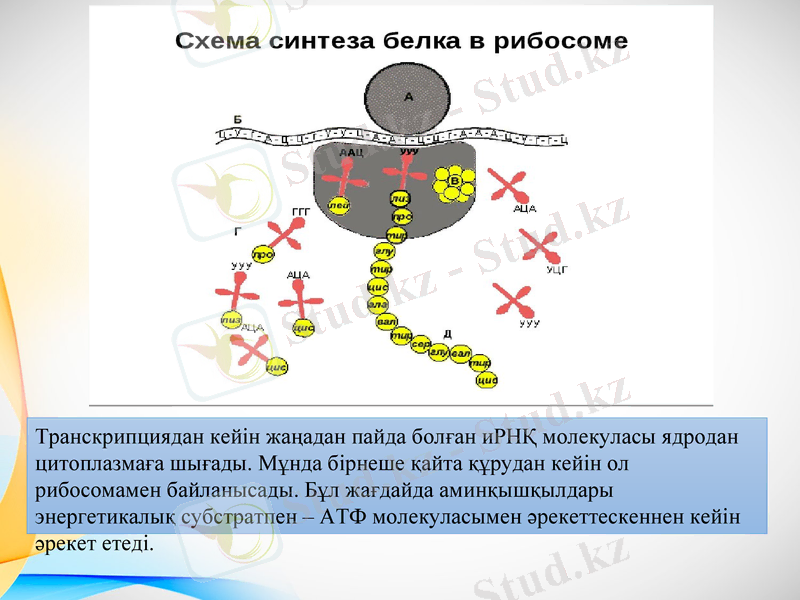
Транскрипциядан кейін жаңадан пайда болған иРНҚ молекуласы ядродан цитоплазмаға шығады. Мұнда бірнеше қайта құрудан кейін ол рибосомамен байланысады. Бұл жағдайда аминқышқылдары энергетикалық субстратпен - АТФ молекуласымен әрекеттескеннен кейін әрекет етеді.

Аминқышқылдары мен иРНҚ әртүрлі химиялық құрамға ие және олар ешқандай қатысусыз өзара әрекеттесе алмайды. Бұл сәйкессіздікті жеңу үшін көлік РНҚ бар. Ферменттердің әсерінен аминқышқылдары тРНҚ-ға қосылады. Бұл пішінде олар рибосомаға және тРНҚ-ға беріледі, белгілі бір амин қышқылы бар, тағайындалған жерде иРНҚ-ға қосылады. Әрі қарай рибосомалық ферменттер қосылған амин қышқылы мен полипептид арасында пептидтік байланыс түзеді. Содан кейін рибосома ақпараттық РНҚ тізбегі бойымен қозғалады, келесі аминқышқылын бекіту үшін учаске қалдырады.

Трансляция немесе ақуыз биосинтезі полипептидтің N ұшынан басталып С ұшына қарай жүреді. Ақуыз биосинтезіне рибосоманың екі бөлшегі, а-РНҚ, т-РНҚ, 20 аминқышқылдар, аминоацил-т-РНҚ-синтетаза ферменттері және басқа -да қосымша ақуыз факторлары қатынасады және олар түрліше қызметтер атқарады. а-РНҚ ақуыз биосинтезі үшін матрица (қалып) болып табылады, р-РНҚ лар (5s рРНҚ, 5, 8s рРНҚ, 18s рРНҚ, 28s рРНҚ) рибосома бөлшектерінің құрамына кіреді, ал рибосомалар болса цитоплазмада ақуыз биосинтезін жүргізуші органеллалар болып табылады. Рибосомалар гиалоплазмада еркін күйінде (полисомалар) кездесуі мүмкін, оларда ішкі ақуыздар синтезделінеді және мембраналармен байланысқан күйінде кездесуі мүмкін. Бұл жерде «экспорттық», мембраналық және лизосомалық ақуыз молекулалары синтезделінеді
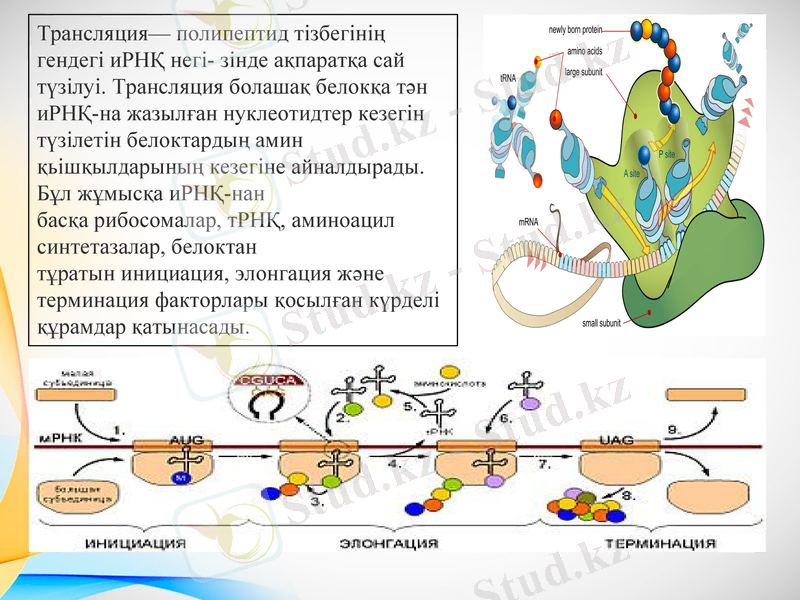
Трансляция- полипептид тізбегінің гендегі иРНҚ негі- зінде ақпаратқа сай түзілуі. Трансляция болашақ белокқа тән иРНҚ-на жазылған нуклеотидтер кезегін түзілетін белоктардың амин қьішқылдарының кезегіне айналдырады. Бұл жұмысқа иРНҚ-нан басқа рибосомалар, тРНҚ, аминоацил синтетазалар, белоктан тұратын инициация, элонгация және терминация факторлары қосылған күрделі құрамдар қатынасады.

Ақпарат тек бір ДНҚ тізбегінен (+ тізбектен) оқылады, өйткені генетикалық кодтың қасиеттеріне сүйене отырып, ДНҚ-ның толықтырушы бөлімдері кодтың толықтырушы дегенерациясының болмауына байланысты бірдей ақуыздың құрылымын кодтай алмайды. Транскрипцияны төрт суббірліктен (ααββ') тұратын және ДНҚ көзіне қатысты ерекшелігі жоқ РНҚ полимераза ферменті жүргізеді. Транскрипцияның бастапқы кезеңінде ферментке ДНҚ-ның белгілі бір бөлігін, промоторды тануды жүзеге асыратын s-фактор деп аталатын бесінші бөлім қосылады. Промоутерлер транскрипцияланбайды. Олар нуклеотидтердің белгілі бір тізбегінің болуына байланысты S факторымен танылады. Бактериалды промоторларда ол Прибнов блогы деп аталады және ТАТААТ түрінде болады (аздап өзгереді) . РНҚ полимераза ферменті промоторға қосылады. ИРНҚ тізбегінің өсуі бір бағытта жүреді, транскрипция жылдамдығы секундына 45-50 нуклеотидке тең. Бастама сатысында тек 8 нуклеотидтің қысқа тізбегі синтезделеді, содан кейін s факторы РНҚ полимеразасынан бөлініп, элонгация кезеңі басталады. ИРНҚ тізбегін құру қазірдің өзінде тетрамер ақуызымен жүреді. Ақпарат оқылатын бөлім транскриптон деп аталады. Ол терминатормен аяқталады - stop сигналының рөлін атқаратын нақты нуклеотидтер тізбегі. Терминаторға жеткенде РНҚ полимераза ферменті жұмысын тоқтатады және белокты тоқтату факторларының көмегімен матрицадан бөлінеді.

Иницияция Сатысы
Трансляция инициалдаушы кешеннің түзілуінен басталады. Инициалдаушы кешеннің құрамы: рибосоманың кіші бірлігі 40 S; мРНК; аминоацил-тРНКмет; рибосоманың үлкен бірлігі. Сонымен инициалдаушы кешеннің түзілу реттілігі: 1) мРНК рибосоманың кіші суббірлігімен комплементарлық принципке сәйкес байланысады, ескеретіні рибосоманың Р -сайтында әр уақытта мРНК -ның АУГ кодоны келеді, 2) мРНК -ның АУГ кодонына, метионин аминқышқылымен байланысқан тРНК-ның антикодоны келіп жалғанады. Яғни синтез метионин қышқылынан басталады деген сөз. 3) Рибосоманың үлкен суббірлігі келіп қосылады. Осылайша жұмысқа дайын рибосома ұйымдасады. Трансляцияның келесі сатысы: аминоқышқылдың белсендірілуі (активация) тРНК-ға тіркелуі жүреді.
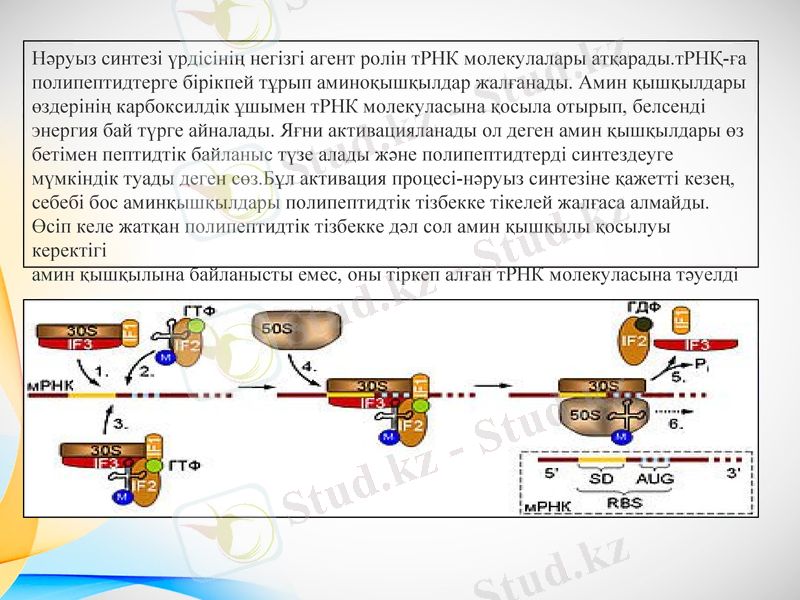
Нәруыз синтезі үрдісінің негізгі агент ролін тРНК молекулалары атқарады. тРНҚ-ға полипептидтерге бірікпей тұрып аминоқышқылдар жалғанады. Амин қышқылдары өздерінің карбоксилдік ұшымен тРНК молекуласына қосыла отырып, белсенді энергия бай түрге айналады. Яғни активацияланады ол деген амин қышқылдары өз бетімен пептидтік байланыс түзе алады және полипептидтерді синтездеуге мүмкіндік туады деген сөз. Бұл активация процесі-нәруыз синтезіне қажетті кезең, себебі бос аминқышқылдары полипептидтік тізбекке тікелей жалғаса алмайды. Өсіп келе жатқан полипептидтік тізбекке дәл сол амин қышқылы қосылуы керектігі амин қышқылына байланысты емес, оны тіркеп алған тРНК молекуласына тәуелді
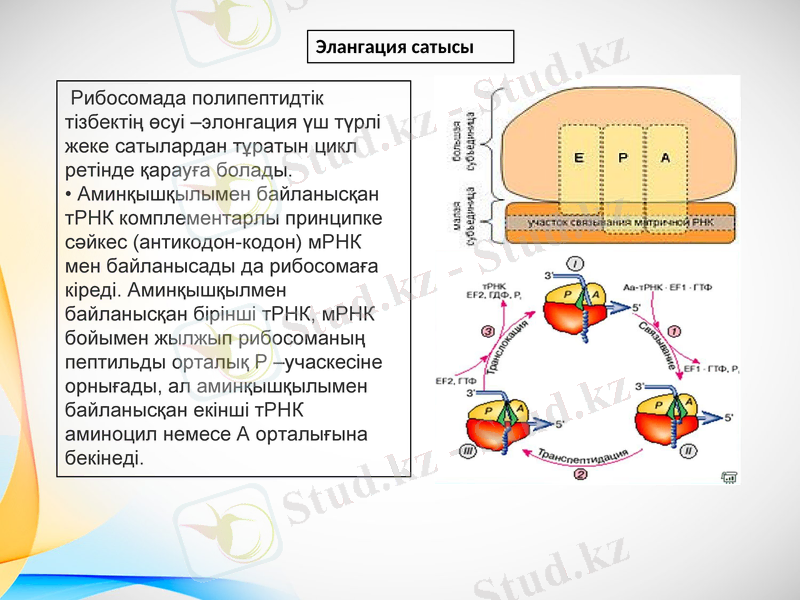
Рибосомада полипептидтік тізбектің өсуі -элонгация үш түрлі жеке сатылардан тұратын цикл ретінде қарауға болады. • Аминқышқылымен байланысқан тРНК комплементарлы принципке сәйкес (антикодон-кодон) мРНК мен байланысады да рибосомаға кіреді. Аминқышқылмен байланысқан бірінші тРНК, мРНК бойымен жылжып рибосоманың пептильды орталық Р -учаскесіне орнығады, ал аминқышқылымен байланысқан екінші тРНК аминоцил немесе А орталығына бекінеді.
Элангация сатысы

Бұл кезеңде полипептидтік тізбектің тікелей синтезі жүреді. Элонгация процесі көптеген циклдерден тұрады. Элонгацияның бір циклі-өсіп келе жатқан полипептидтік тізбекке бір аминқышқылының қосылуы.
Бастау кезеңінде рибосоманың P-сайтын амин қышқылы метионині бар алғашқы тРНҚ алады. Бірінші элонгация циклінде AA-тРНҚ екінші кешені рибосоманың a-сайтына енеді. Бұл антикодон келесі (бастапқы AUG) кодонға қосымша болатын тРНҚ болады.
A(аминоацил) - және P (пептидил) -сайттарда AA-trna кешендері бар, сондықтан аминқышқылдары арасында химиялық реакция жүреді және пептидтік байланыс пайда болады.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz