Сүйек тінінің көмірсулары: құрамы, метаболизмі және биологиялық қызметі


Сүйек тінінің көмірсулары
Орындаған: Аманбай Аида
Факультет:Жалпы медицина
Қабылдаған:Тулеева Г. Т.

Жоспар
Сүйек тінінің көмірсулары және биологиялық ролі.
Көмірсулар метаболизмі.
Көмірсулар алмасуы.
Гликогеннің синтезделуі туралы жалпы түсінік қалыптастыру.
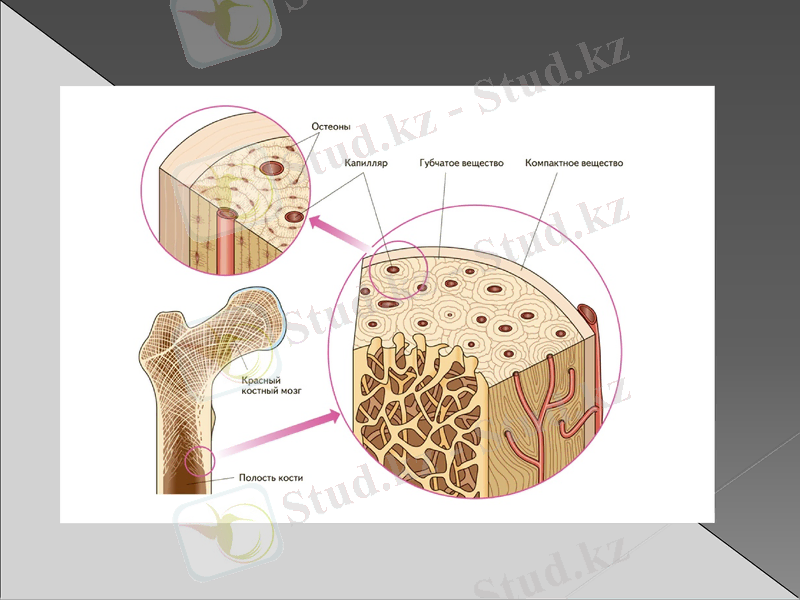

Сүйек тінінің құрамының ерекшеліктері
Жасушалардың саны өте аз.
Коллагеннің мөлшері өте көп (органикалық заттардың 95% құрайды) .
Коллагеннің талшықтары өте тығыз, жоғары дәрежедегі тәртіппен орналасады.
Коллагеннің құрамында оксипролин, лизин мен оксилизин көп, оларға фосфаттар қосылып, кристаллизация орталығын түзеді, ізінше гидроксилапатиттер түзіледі.
Коллагеннің құрамында фосфаттың мөлшері көп, олар серинмен байланысқан, осындай фосфаттарға Са2+ иондары қосылып, кристаллизация орталығын түзеді, ізінше гидроксилапатиттер түзіледі.
Минералды тұздар (әсіресе Са-дің) өте көп.
Сүйек тінінің жасушалары
Остеобласттар- коллаген, ГАГ, ферменттерді түзеді.
Остеоциттер- остеобласттардан түзіледі, қызметі ұқсас.
Остеокласстар- СТ-нің жасушааралық затын ыдыратады (митохондриялар, лизосомалар көп) .

Сүйек тіні көмірсулары
Сүйек тініндегі көмірсулар жасуша ішінде және жасушадан тыс локализацияланған. Жасушаішілік көмірсулар гликоген, ал жасушадан тыс - гликозаминогликандар. Гликоген және глюкоза негізінен энергетикалық функцияны орындайды және глюкозаның ыдырауы АТФ, минералдану үшін қолданылады. Остеобласттар адам жасының ұлғаюымен сүйек тініндегі гликогеннің құрамы бір грамм тіндерге 15-20 мг-ден 5-10 мг-ға дейін төмендейді. жас жасушаларда гликолиз арқылы 60% АТФ, ал ескілерінде - 85% қалыптасады. Остеоциттерде гликоген жоқ (немесе іздер) және бәрі АТФ гликолизге байланысты алынады.

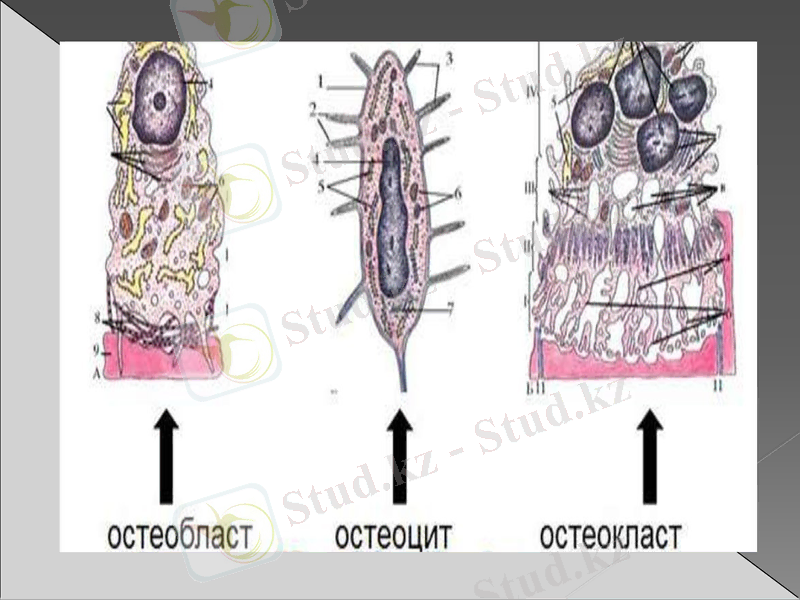
Остеоцит
Остеоциттер -сүйек жасушалары. Бұлар санына қарай сүйек ұлпасының дефинитивті клеткалары, бөлінуге деген қасиетінен айрылған.
Остеоциттердің компакті үлкен ядро және әлcіз базафильді цитаплазмасы бар. Органеллалары әлсіз дамыған жасуша центриом жасушада табылмаған. Сүйек клеткалары сүйек беткейлерінде немесе лакуналарда жатады және де остециттің контурын қайталайды. Беткейлердің ұзындығы 22-55 мкм, ені 6-14 мкм дейін өзгеріп отырады. Сүйек беткейлерінің каналдары ұлпалық сұйықтықпен толтырылған. Остеоциттер мен қан арнасында зат алмасу ұлпалық сұйықтық арқылы жүзеге асады.

Остеобласт
Бұлар жас жасушалар, сүйек ұлпасын құрайды. Қалыптасқан сүйектер сүйекүстінің (надкостница) терең қатпарларында және травмадан кейін регенерация жүрген сүйек ұлпаларында кездеседі. Қалыптасып жатқан сүйектің бүкіл беткейін үздіксіз қабатпен қаптайды. Остеобласттар әр түрлі формада болады: кубтәрізді, пирамидалық, бұрышты (углаватый) . Оның денесінің пішіні шамамен 15-20 мкм. Ядролары домалақ немесе овальды формада, бір немесе бірнеше ядрошықтан тұрады. Остеобласттардың цитоплазмасында гранулалық эндоплазмалық тор, митохондриялар және Гольджий жиынтығы жақсы дамыған. Остеобласттар, остеоциттер сияқты бөлінбейтін клеткаларға айналады.

Остеокласт
Бұл клеткалардың табиғаты гематогенді, шеміршек пен сүйекті бұзу қабілеті бар. Олардың диаметрі 90 мкм және одан үлкен дәрежеге дейін жетеді. Оларда үш бірнеше ядро болады. Цитоплазмасы әлсіз базафильді, кейде оксифильді остеокласттар өзінің қоршаған ортасына СО2 бөледі, ал карбоангидраза ферменті қышқылдардың (Н2 СО3) қалыптасуына әкеледі. Ол сүйектің органикалық матрицасын және кальций тұздарын ерітеді. Остеокласт митохондрия мен лизосомаларға бай. Грануналық эндоплазмалық тор нашар дамыған. Остеокласт сүйек затымен жанасқан жерде лакуна түзіледі.

Сүйек тінінің функциялары
құрылымдық-тірек
механикалық қорғау
көптеген макро-және микроэлементтерге арналған депо
ішкі ортаның қышқылдық-негіздік тепе-теңдігін сақтау.

Сүйек құрамындағы көмірсу
Сүйек протеогдикандарының простетикалық топтарының құрамы әртүрлі кезеңдерде дамып өзгереді. Жас сүйекте гиалуронат, жетілген сүйекте сульфат гликозоаминогликандар (ГАГ) (хондроитин және кератансульфаттар) көбірек. Полианионды сульфат топтары Са2+ - ны белсенді түрде байланыстырады, оның депосын жасайды. ГАГ-тың жойылуы Ca2 + байланыстыруды азайту үшін, ал синтезді белсендіру Ca2 + және сүйектің дамуының белгілі бір кезеңдері минералдануға ықпал етеді. ішінде ең көбі хондроитин сульфат, олардың ішінде хондроитин -4-сульфат және . Хондроитин сульфаттары (ХС) және кератанульфаттар (КС) простетикалық топтар болып табылады, жасушааралық матрицаның протеогликандары, олардың КЛ1-мен байланысын қамтамасыз етеді.

Жас сүйек тінінде ГК мөлшері ХС-нан басым болады, бірақ екі гликозаминогликанның синтезі протеогликандарды KL1-ге қосу және дұрыс қалыптастыру үшін қажет коллаген талшықтары. Осылайша, байланысты формалары KL1 талшықты құрылымын тұрақтандыру және цементтеуді қамтамасыз етеді. Еркін ГК полианион қасиеттеріне байланысты катиондар мен суды белсенді сіңіреді, сонымен қатар су мен электролиттердің алмасуын реттейді.

Көмірсулар алмасуы
Көмірсулардың аралық алмасуы ағзада көмірсулар түрлі
өзгерістерге ұшырайды, нәтижесінде энергия және
ыдыраудың соңғы өнімдері - сүт қышқылы, СО2 және Н2О
бөлінеді. • Жасушадағы көмірсулардың өзгеру жолдары аэробты және анаэробты тотығу жолдарымен жүреді. • Анаэробты тотығу глюкоза ыдырауынан (гликолиз) да гликогеннің ыдырауынан да (гликогенолиз) басталуы мүмкін. • Анаэробты гликолиз(гликогенолиз) үрдісін екі сатыға бөлуге болады: 1. Дайындық кезеңі: глюкозадан екі молекула триоздың, яғни фосфорлы эфирлердің түзілуі. 2. Гликолиттік тотығу-тотықсыздану реакциясы жүзеге асады да, АТФ түзіледі.

Көмірсулар анаболизмі
Жасыл өсімдіктер, балдырлар, цианобактериялар су мен көмір қышқыл газынан, күннің қуатының арқасында көмірсулар түзеді. Ол процесс фотосинтез деп аталады.
6СО2 + 6Н2 О С6 Н12О6 + 6О2 фотосинтез
Фотосинтез :
Энергиясы төмен көміртек қосылысының тотыққан түрі күн сəулесінің квант энергиясының əсерінен жоғары энергиялы көміртектің тотықсызданған қосылысына айналады. Мұндай қосылыстарды адам мен жануарлар (гетеротрофты ағзалар) көміртек жəне энергия көзі ретінде пайдаланады.
Оттек молекуласы түзіледі, бұл процесс ауадағы бос оттектің жалғыз көзі. Фотосинтездің екі фазасы болады: жарықта жүретін фаза жəне қараңғыда жүретін фаза. Жарықта жүретін фаза үш процестен тұрады:
а) күн сəулесінің квант энергиясының əсерінен су молекуласының
ыдырауы: 224 n h v _ 2Н2 О 4Н+ + 4е + О2
ə) түзілген электрондар энергиясы өте жоғары, ол энергия АДФ-тың
фосфорилденіп АТФ-ке айналу реакциясына жұмсалады.
б) электрондар энергиясының бір жартысы НАДФ+ -ты
тотықсыздандыру реакциясына жұмсалады.
Қараңғыда жүретін процесс көмір қышқыл газының (СО2 ) көмірсуларға
айналуы:
12НАДФН·Н+ 6СО2 С6 Н12О6 18АТФ

Гликоген
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz