Саңырауқұлақтардың митохондриялық геномы мен динамикасы: тұқымқуалау, бірігу/бөліну және митофагия


Саңырауқұлақтардың митохондриялық геномы

01
Саңырауқұлақтардың митохондрияларын анықтау жолдары
Жоспар:
03
Митохондриялардың бөлінуі және қосылуы
02
Митохондриялардың тұқымқуалауы
04
Митофагия
Митохондрия
Митохондрия (грек тілінен. μίτος - жіп және βόνδρος-дән, дән) - өз геномы бар жасушаның өздігінен репликацияланатын жартылай автономды екі мембраналы органоидтары. Митохондриялық геном, ядролық геномнан айырмашылығы, бір немесе бірнеше сақиналы, сирек сызықты, ДНҚ молекулалары (MTDNA) .
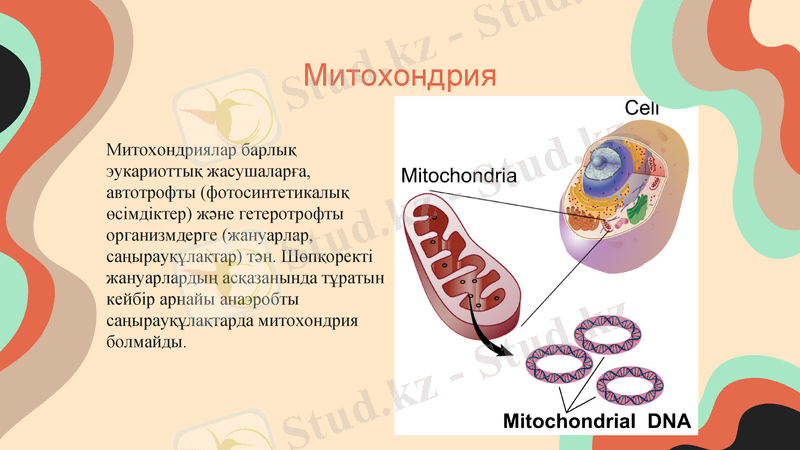
Митохондрия
Митохондриялар барлық эукариоттық жасушаларға, автотрофты (фотосинтетикалық өсімдіктер) және гетеротрофты организмдерге (жануарлар, саңырауқұлақтар) тән. Шөпқоректі жануарлардың асқазанында тұратын кейбір арнайы анаэробты саңырауқұлақтарда митохондрия болмайды.
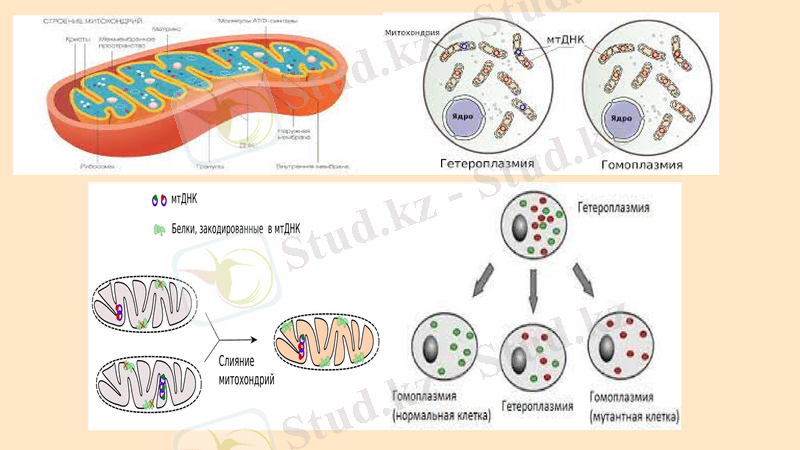

Митохондрия ДНК
Митохондриядағы ДНҚ гистон ақуыздарымен байланыс жасамайтын циклдік молекулалармен ұсынылған. Митохондриялық ДНҚ өте біртекті, айырмашылығы интрондардың/транскрипцияланбаған аймақтардың шамасында. Митохондриялық ДНҚ кластерлерге жиналған бірнеше көшірмелермен ұсынылған. Митохондрияларда гендердің стандартты жиынтығы бар: тотығу фосфорлану процестеріне қатысатын тыныс алу тізбегі ферменттерінің гендері, р-РНҚ гендері, тРНҚ және АТФаза гендері. Митохондриялық ДНҚ жеке аймаққа - нуклеоидқа жиналады. Саңырауқұлақ митохондриясында 1-ден 10-ға дейін нуклеоидтар болуы мүмкін. Митохондриялық ДНҚ синтезі ядродағы ДНҚ синтезімен байланысты емес. S. cerevisiae ашытқы жасушаларында стационарлық фазада төрт геномы бар 22 немесе одан да көп митохондрия бар.
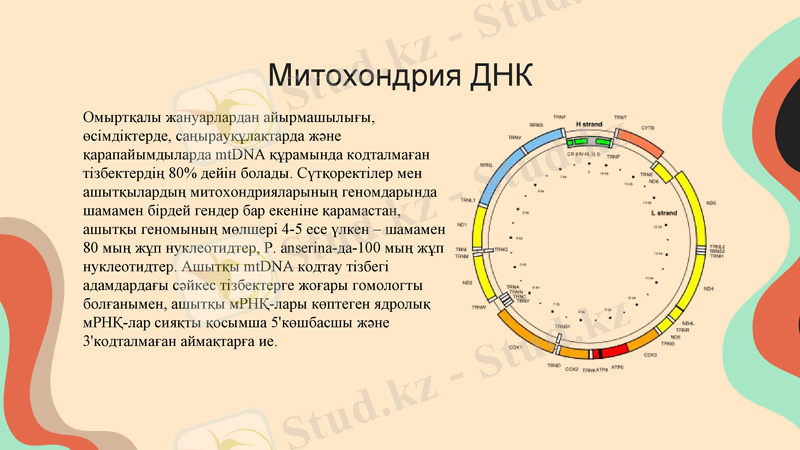
Митохондрия ДНК
Омыртқалы жануарлардан айырмашылығы, өсімдіктерде, саңырауқұлақтарда және қарапайымдыларда mtDNA құрамында кодталмаған тізбектердің 80% дейін болады. Сүтқоректілер мен ашытқылардың митохондрияларының геномдарында шамамен бірдей гендер бар екеніне қарамастан, ашытқы геномының мөлшері 4-5 есе үлкен - шамамен 80 мың жұп нуклеотидтер, P. anserina-да-100 мың жұп нуклеотидтер. Ашытқы mtDNA кодтау тізбегі адамдардағы сәйкес тізбектерге жоғары гомологты болғанымен, ашытқы мРНҚ-лары көптеген ядролық мРНҚ-лар сияқты қосымша 5'көшбасшы және 3'кодталмаған аймақтарға ие.

Саңырауқұлақтардағы митохондриялық тұқымқуалау
Тұқым қуалау процесінде митохондриялық геномның тұтастығын сақтау тыныс алу қызметі үшін өте маңызды. Саңырауқұлақтардағы митохондриялық геномның бөлінуін бақылайтын факторлар аз зерттелген. Жоғары эукариотты организмдердің көпшілігінде . Ascomycota Neurospora tetrasperma және N. crassa класындағы мицелий саңырауқұлақтарында митохондриялардың біртектес тұқым қуалаушылығы көрсетілген. Мамандандырылған жыныс жасушаларының (трихогин мен конидия) бірігуі жағдайында да, әртүрлі mat-локустары бар ядроларда ерекшеленетін вегетативті мицелийдің гомокариоттық жасушаларының бірігуі кезінде де ядро акцепторы жасушаларының митохондриялары сақталады. Бүршіктенетін ашытқы екі ата-аналық мұрамен сипатталады.

Саңырауқұлақтардағы митохондрия қозғалысы
Saccharomyces cerevisiae жасушаларында, сондай-ақ Aspergillus nidulans жасушаларында актин микрофиламенттері митохондриялық тасымалдауда үлкен рөл атқарады. Ашытқыда, Schizosaccharomyces pombe, және Neurospora crassa жасушаларында, керісінше, митохондриялардың таралуына негізінен микротүтікшелер қатысады. Зерттеушілер органеллалардың қозғалысын гифаларда, протопластта, жасуша фрагменттерінде және жасуша қабырғасы жоқ мутанттарда 1. 4 мкм/с жылдамдықпен байқады. Микротүтікшелердің нокодазолмен жойылуы митохондриялардың қозғалғыштығын төмендетті, ал цитохалазин d микрофиламенттерінің жойылуы мұндай әсер етпеді. Митохондриялардың микротүтікшелермен байланысуын осы органеллалардың перифериялық ақуыздары жүзеге асыратыны анықталды. Мұндай ақуыздарға кинезиндер тұқымдасынан немесе динеинге байланысты ақуыздар жатады.

Саңырауқұлақтардағы митохондрия қосылысы англ. fusion мен бөлінуі англ. fission или division
Митохондриялардың динамикалық түрде жүретін бірігу және бөліну процестері (динамикасы) хондриоманың тұрақсыздығын және жасушаның АТФ қажеттіліктеріне сәйкес митохондриялардың құрылымы мен қызметін генетикалық бақылау механизмдері арқылы органеллалардың қызметін реттеуді қамтамасыз етеді.
Митохондриялық Морфология және геном көшірмелерінің саны синтез бен бөліну белсенділігінің тепе-теңдігіне байланысты. Біріктіру бағытындағы өзгеріс жасушаға ұзартылған өзара байланысты митохондриялық желілерді құруға мүмкіндік береді, сонымен бірге бөліну бағытындағы сдысу көптеген морфологиялық және функционалды түрде әртүрлі кішкентай сфералық органеллаларды тудырады.
Митохондриялық бөлімнің жасушалық қажеттіліктерге бейімделуі көптеген маңызды процестердің кілті болып табылады

Саңырауқұлақтардағы митохондрия қосылысы англ. fusion мен бөлінуі англ. fission или division
Митохондриялардың бірігуі мен бөлінуінің негізгі компоненттері мембраналық ақуыздар болып табылады: M1 (Э ақуызы), M2, Mm 10, және mdm12, мембраналық митохор құрылымын құрайды. Митохор митохондриялар мен мднқ-ны цитоскелетпен байланыстырады (ашытқыда актин жіпшелері бар), бұл органеллалар мен олардың ДНҚ-сының жасушада бақыланатын қозғалысын қамтамасыз етеді. Түтікшелі митохондриялық құрылымдарды қалыптастыру және mtdna тұқым қуалауы үшін ммм1р арқылы мембраналық митохор кешенімен әрекеттесетін митохондрияның ішкі мембраналық ақуыздары - mdm31 және Mdm32 қажет. MDM 10, DM12, M1 және M2 жойылуы MDM31 және MDM 32 гендерінің жойылуы аясында өлімге әкеледі. Mmm1, mdm10, mdm12 немесе mmm2 гендерінің жойылуы органеллалар морфологиясының тән өзгеруіне әкеледі-алып дөңгелек деполяризацияланған митохондриялардың түзілуі.

Westermann, 2008

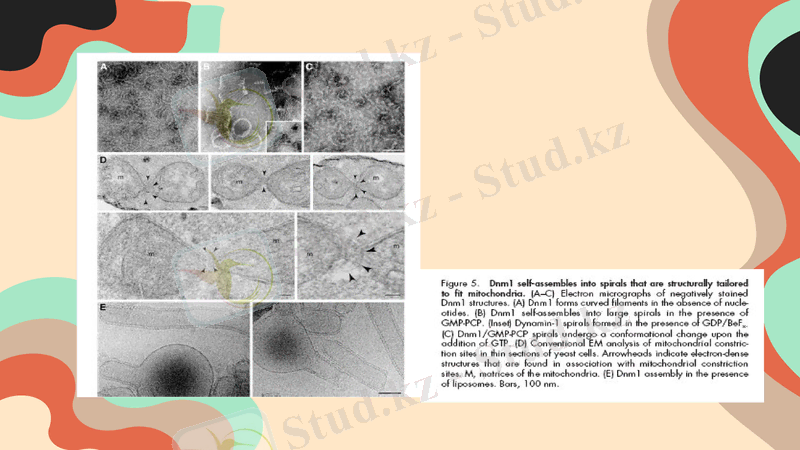
Dnm1, Fis1, Mdv1, Caf4 және Mdm33 ақуыздары қатысатын митохондриялардың бөлінуін түсіндіретін бірнеше модельдер ұсынылған

Ашытқыдағы митохондриялық бөлінудің молекулалық аппаратының моделі
Mdv 1 және Caf 4 домендік ұйымға ие және Dnm 1-ді Fis1-мен байланыстыратын адаптерлер қызметін атқарады. Олардың спиральды бұралған аймағы гомоолигомерлердің өзара әрекеттесуін жүзеге асырады. Dnm1 митохондрияға митохондрияны қоршап тұрған спиральдарды құрайтын динамикалық олигомерлер ретінде жиналады. Dom1 доменінің Mdv 1 немесе CAF 4 ақуызының WD40 қайталанатын аймағымен әрекеттесетіні белгісіз. Mdm33 матрицада бірнеше болжамды спиральды бұралған аймақтарды қамтиды. Әрбір полипептидтің N-ұштары белгіленген; αA және αB, α спираль N-терминал соңында Mdv1 және Caf4; B, кірістіру B; cc, спиральды бұралған; GED, Gtpase эффекторлық домен; IM, ішкі мембрана; IMS, мембрана аралық кеңістік; MD, орта домен; OM, сыртқы мембрана; TPR, кездейсоқ төрт пептидті қайталаулар.
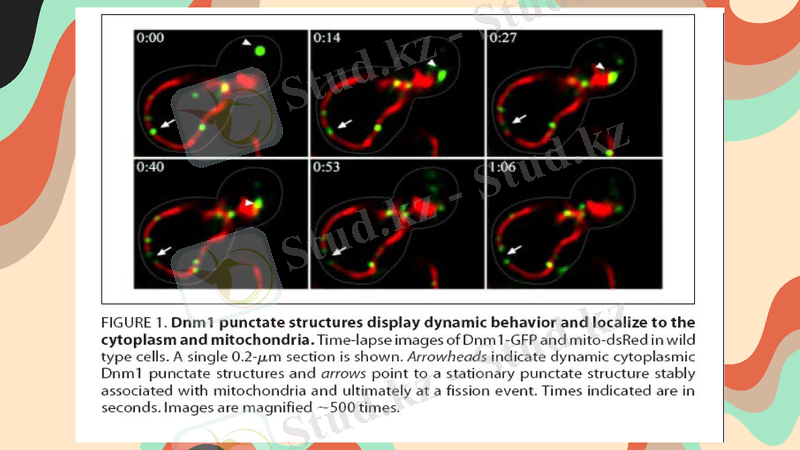

Митохондриялар бірігуі
Теориялық тұрғыдан митохондриялардың бірігуінің екі жолы мүмкін: ішкі және сыртқы мембраналардың бір мезгілде және бөлек әсерімен. Практикалық тұрғыдан алғанда, бұл мәселе қосымша зерттеулерді қажет етеді. Митохондрияның ішкі мембранасының бірігу механизмі қазіргі уақытта жұмбақ болып табылады. Бұл процестің ең ықтимал қатысушысы-динамин тәрізді ақуыз, бірақ оның митохондриялық синтезге қалай қатысатыны қазіргі уақытта белгісіз.

Ашытқыдағы митохондриялық бірігуінің молекулалық аппаратының моделі.
Екі fzo1 молекуласы екі іргелес митохондрияны параллельге қарсы спираль тәрізді бұралған с-терминал аймағын құрастыру арқылы байланыстырады. Митохондриядағы Fzo1 белгісіз құрамдағы 800 kDa ірі кешенінің бөлігі болып табылады (16) . Ugo 1-де суретте көрсетілгендей мембрананы қатайтатын бес аймаққа дейін бар, алайда бірнеше ондаған трансмембраналық аймақтары бар басқа балама топология ұсынылған. Ugo 1-нің қай бөлігі Fzo1 және Mgm1-мен әрекеттесетіні белгісіз. Mgm 1 екі формада болады митохондрия, ішкі мембранада трансмембраналық ауданы бар ұзын форма және Pcp1 бөлінген кезде бұл аймақсыз қысқа форма. Mgm1 доменінің қайсысы сыртқы мембрана компоненттерімен әрекеттесетіні белгісіз. Әрбір полипептидтің N-ұштары белгіленген; cc, спираль тәрізді бұралған; GED, gtpase эффекторлық домен; IM, ішкі мембрана; IMS, мембрана аралық кеңістік MD, орта домен; OM, сыртқы мембрана
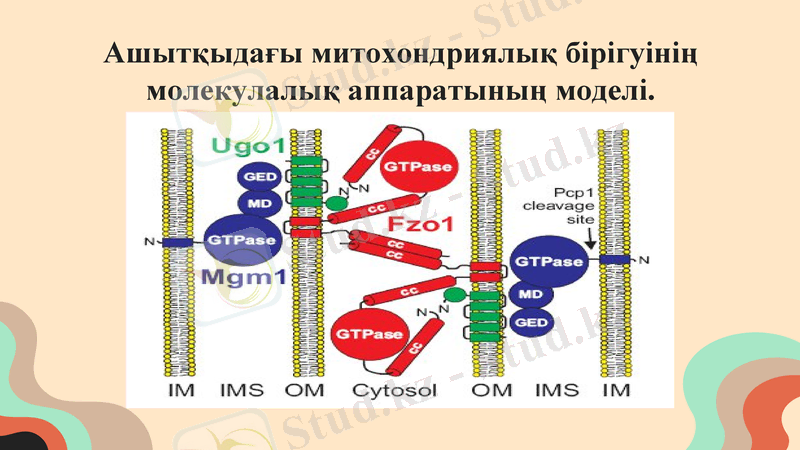
Ашытқыдағы митохондриялық бірігуінің молекулалық аппаратының моделі.
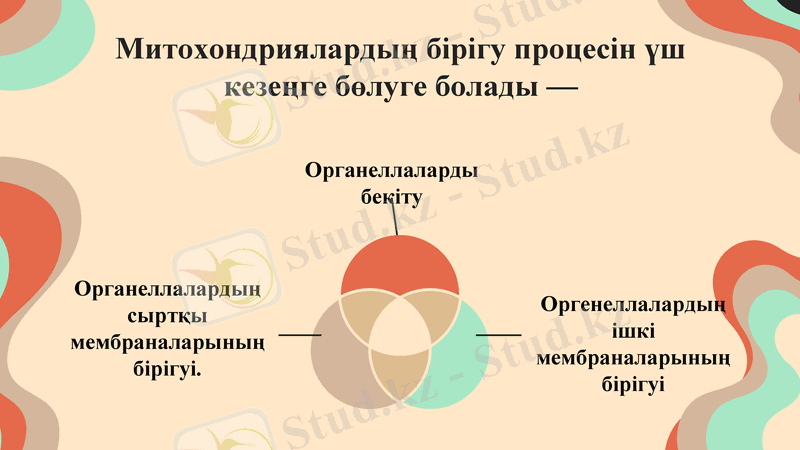
Митохондриялардың бірігу процесін үш кезеңге бөлуге болады -
Органеллаларды бекіту
Органеллалардың сыртқы мембраналарының бірігуі.
Оргенеллалардың ішкі мембраналарының бірігуі

- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz