SV40 вирусының молекулалық вектор ретінде қолданылуы және түрлендірілуі

Әл-Фараби атындағы Қазақ Ұлттық Университеті

Факультет: Биология және биотехнология
РЕФЕРАТ
Тақырыбы: «SV40 вирусы молекулалық вектор ретінде»
Орындаған: Әбілова Дана Талғатқызы
(БТ17-18К)
Қабылдаған: Бисенбаев А. К.
Алматы 2020жыл
Жоспар:
І. Кіріспе:
ІІ. Негізгі бөлім:
- 2. 1 SV40 негізіндегі векторлар
- 2. 2 SV40 векторларын түрлендіру
ІІІ. Қорытынды:
ІV. Пайдаланған әдебиет:
І. Кіріспе:
SV40 вирусы- бұл маймыл жасушаларында бетаполомавирус тұқымынан табылған полиомавирустың бір түрі, бұл типке жатады. Басқа полиомавирустар сияқты, SV40 геномы дөңгелек қос ішекті ДНҚ-мен ұсынылған. Миллиондаған адамдар SV40 вирусын жұқтырды, өйткені ол 1960 жылдары полиомиелитке қарсы вакцинада болған.
Вирус 1960 жылы полиомиелитке қарсы вакцинаны өндіруде қолданылатын Rhesus macaque бүйрек жасушаларының культурасында оқшауланды. Вирус жұқтырған жасыл маймыл жасушалары ерекше вакуольдер құрады. 1978 жылы вирустың толық геномын Гент университетінде Уолтер Файерс реттеді. Резус макака жасушаларын жұқтыру асимптоматикалық емес, вирус табиғатта көптеген маймылдардың популяцияларында кездеседі. Дені сау адамдарда SV40 ауруды сирек тудырады, бірақ маймылдың иммун тапшылығы вирусын (SVM) жұқтырған маймылдарда SV40 адамның JC және BK полиомавирустары сияқты әрекет етеді, олар прогрессивті мультифокалды лейкоэнцефалопатияға ұқсас бүйрек аурулары мен демиелинацияны тудырады. Гамбустар сияқты басқа түрлерде SV40 әртүрлі ісіктерді тудырады, әдетте саркомалар. Бөртпелерде SV40 онкогенді Т-антигені нейроектодермальды ісіктер және медуллобластомалар сияқты ми ісіктерін модельдеу үшін қолданылады.
SV40 вирусында геномдық ДНҚ-ның 5000-ға жуық базалық жұбы бар мозозедральды вирус бар. Вирион VP1 гликопротеинді қолдана отырып, жасуша бетіндегі MHC 1-ші рецепторларға қосылады. Жасуша ядросының ішінде клеткалық РНҚ полимераза II ерте гендерді білдіреді. Транскрипцияланған мРНҚ үлкен және кішкентай Т-антигендерін кодтайтын екі бөлікке бөлінген. Үлкен Т-антигеннің шамамен 5% -ы клетканың плазмалық мембранасына, ал 95% -ы ядроға енеді. Үлкен Т-антигені вирустық ДНҚ-ның үш учаскесімен байланысады, I және II учаскелермен байланысады, ерте РНҚ-лардың синтезін реттейді, екінші жасушамен байланыстыру әрбір жасуша циклінде II кездеседі. I сайтқа байланыстыру репликацияның басында ДНҚ репликациясын тудырады. Вириондар жасуша ядросында жиналады.
2. 1 SV40 негізіндегі векторлар
Вирус және оның геномы. SV40 - сфералық вирус; оның геномы бір жабық дөңгелек қос ішекті ДНҚ-дан тұрады (5. 34-сурет) . Вирустық бөлшектің құрамында үш құрылымды вирус кодталған ақуыз бар. VP1 - вирионның негізгі ақуызы, капсидтің негізгі компоненті; VP2 және VP3 аз мөлшерде болады. Вирустық бөлшектегі ДНҚ хроматин түрінде болады және төрт гистонмен байланысады (2-сурет) . SV40-тің табиғи қожайыны - резус-макака (Masas malatta), бірақ зертханада африкалық жасыл маймыл бүйрек жасушалары (Cercopithecus aethiops) негізінен хост ретінде қолданылады. Бүкіл SV40 геномының нуклеотидтер тізбегі 5243 а. к. -тан тұратын шифрдан шығарылды, бес вирустық ақуызды кодтайтын бөлімдер анықталды: кішкентай Т-антиген, ірі Т-антиген, VP1, VP2 және VP3. Суретте. 5. 34-те вирустық геномның картасы көрсетілген. Бүкіл геном 10 бөлікке бөлінген; көрсеткілердегі сандар нуклеотид жұптарының санын көрсетеді. Бірінші жұп ДНҚ репликациясының (ori) бастауында.
Virion SV40 ұяшыққа еніп, ядроға жіберіледі. Мұнда ол «белдеулер» жасайды, содан кейін қатаң уақыт тізбегінде - гендерді экспрессиялау. Жаңа вирустық бөлшектер маймыл бүйрек жасушаларының монолайзерін жұқтырғаннан кейін шамамен 30 сағаттан кейін пайда болады және 4 күннен кейін барлық жасушалар жойылады. Екі ерте ақуыздың гені, кіші және үлкен Т антигендері, алдымен сағат тіліне қарсы транскрипцияланған. Полиадениляция учаскесі картада 2550 нуклеотидінің жанында орналасқан. Үлкен Т-антигендері де жетілген вирустық бөлшектердің құрамына кірмейді, бірақ үлкен Т-антигендері вирустық ДНҚ-ның репликациясында маңызды рөл атқарады, ол ерте ақуыздар пайда болғаннан кейін басталады. Репликация ori-ден басталып, екі бағытта да жүреді және репликативті шанышқылардың жиналу нүктесінде аяқталады. Содан кейін, қызықты қос ішекті ДНҚ молекулаларының өзара байланысы тұйық сақиналы құрылымдардың пайда болуымен жүреді. Әрбір жұқтырған жасушада вирустық геномның 105 данасынан астамы түзіледі. ДНҚ репликациясының басталуынан көп ұзамай ақуыздың ерте синтезі баяулайды және «кеш» гендердің транскрипциясы іске қосылады, олар ориге жақындайды және сағат тілімен бағытта қозғалады, нәтижесінде VP1, VP2 және VP3 ақуыздары синтезделеді.
ДНҚ молекулалары мен вирион ақуыздарының жинақталуы жаңа вирустық бөлшектердің жиналуымен, олардың шығарылуымен және жасушалардың өлімімен аяқталады. Тазартылған SV40 ДНҚ-мен трансфекция нәтижесінде инфекциялық процесс толығымен басталуы мүмкін. SV40 векторларының бірнеше түрлері. SV40 негізіндегі векторларды құрудың жалпы принциптері фаг λ негізінде қолданылатындарға ұқсас. Тиісті шектеу эндонуклеазаларын қолдана отырып, вирустық геномның белгілі бір аймақтары алынып, басқа ДНҚ сегменттерімен алмастырылады. Вирустық ДНҚ-ның қандай нақты сегменті алынып тасталына байланысты, осы немесе басқа вектор түзіледі. SV40 вирусының негізінде құрылған векторлардың үш түрі бар. Олардың әрқайсысының әртүрлі тәжірибелік жүйелерде қолданған кездегі артықшылықтары мен кемшіліктері бар. Біріншіден, бұл сәйкес келетін жасушаларда (маймыл бүйрек жасушаларында) көбейетін және вириондарға жиналатын SV40 векторлары. Олар фаг λ негізінде құрылған мамандандырылған өткізгіш фазалар мен векторларға ұқсас. Екіншіден, бұл SV40 векторлары, олар көбейте алады, бірақ жинақталмайды. Оларды плазмид SV40 векторлары деп атайды. Ақырында, көбейтуге де, орауға да қабілетсіз векторлар бар. Олар ген экспрессиясына ықпал ететін SV40 геномының кіші сегменттерін қамтитын молекулалар.
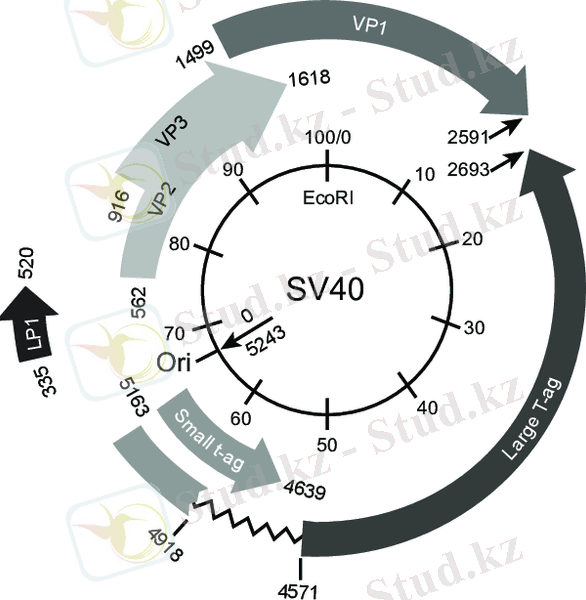
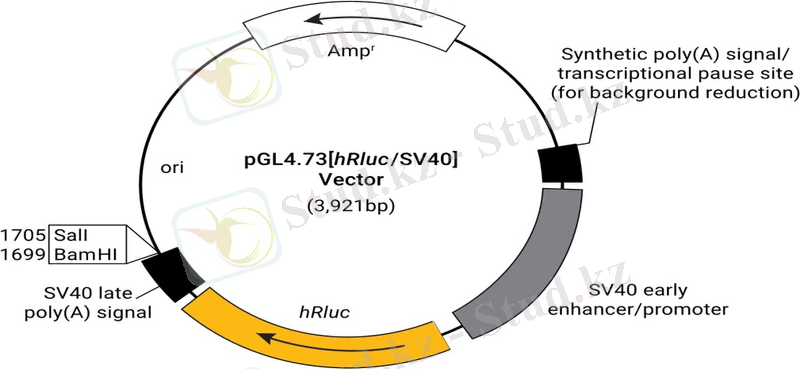
2. 2 SV40 векторларын түрлендіру
Вирустық бөлшектерді қалыптастыру үшін SV40 векторын орау үшін үш маңызды талап орындалуы керек. Біріншіден, векторда ori нүктесін қосқанда 300 а. к. кесінді болуы керек. Бұл вирустық ДНҚ синтезін және мРНҚ синтезіне жауап беретін реттілік тізбегін қамтамасыз ету үшін қажет. Екіншіден, рекомбинантты ДНҚ-ның жалпы ұзындығы 5300 бит / с аспауы керек және 3900 бит / мин-ден аз болуы керек, яғни векторда ДНҚ-ны вирустық бөлшектерге буып-түю үшін қажетті белгілі бір мөлшер болуы керек. Үшіншіден, ірі Т-антигенді ақуыздар, VP1, VP2 және VP3 болуы керек, бірақ олардың рекомбинантты геноммен кодталуы міндетті емес. Қаптамада болуы мүмкін жағдай бұзылмауы үшін SV40 геномына ДНҚ фрагментін қосу іс жүзінде мүмкін емес. Сондықтан жалпы стратегия - вектордың кодтау аймағын жою, оны қажетті ДНҚ фрагментімен алмастыру және жоғалған гендік өнімнің синтезін көмекші вирустың немесе хосттың геномының бөлігі болып табылатын ұқсас генді қолдана отырып қамтамасыз ету (яғни, транс позициядағы геннің өтелуі) . Мысалы, жабайы типтегі көмекші вирус T немесе VP1, VP2 және VP3 ақуыздарын өз қажеттіліктерін де, рекомбинантты вирустың қажеттіліктерін қанағаттандыру үшін жеткілікті мөлшерде синтездей алады.
... жалғасыСіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz