Буаз және сүт түзуші анемияға ұшыраған егеуқұйрықтардың сүт безі микросомаларындағы липидтік асқын тотығу мен антиоксиданттық жүйеге биологиялық белсенді заттардың әсері

ДИПЛОМДЫҚ ЖҰМЫС
Тақырыбы: Анемияға ұшыраған егеуқұйрықтардың антиоксиданттық жүйесіне биологиялық белсенді заттардың әсерін зерттеу
Кіріспе
Қазіргі уақытта анемия организмнің қызмет етуші жүйелерінің үйлесімді жұмысының бұзылуына әкелетін ең кең тараған жағымсыз аурулардың бірі болып табылады. Жүкті және бала емізуші әйелдердегі анемияның зардабын шешу жолдары әлі де толық зерттелмеген. Әйел ағзасындағы анемияның алдын алу және емдеу үшін биологиялық белсенді заттарды қолдану қызығушылық тудыруда.
Соңғы уақытта зерттеушілер табиғатта кең тараған табиғи антиокиданттар - биологиялық белсенді заттарға көп көңіл бөлуде. Көптеген ғалымдар жүргізілген зерттеулер көрсеткендей фикоцианин (цианобактерия Spirulina platensis өнімі) иммундық жүйені күшейтеді және организімнің лимфалық белсенділігін жоғарылатады. Спирулинаның құрамында көптеген витаминдер мен минералды заттар, сонымен қатар организмге қажетті ұлпалардың дамуына және жүйке жүйелерінің қалыптасуына қажетті В 12 витамині де бар. Дегенмен, спирулина құрамындағы фикоцианиннің организмге әсері аз зерттелген.
Анемия - гематологиялық синдромды ауру болып табылады. Анемия байқалған кезде қан құрамындағы эритроциттерде кездесетін гемоглобин (оттегі тасымалдағыш ақуыз) кеміп кетуі орын алады. Гемоглобинмен бірге эритроциттердің кеміп кетуі де мүмкін. Анемия - жүктілік кезде өте жиі кездесетін ауру. Жүкті әйелдердегі анемия жүктіліктің екінші жартысында қатты байқалады. Басқа адамдарға қарағанда жүкті әйелдер арасында анемия жиі кездеседі. Оның себебі жүктілік кезде әйел ағзасында болатын өзгерістерге, қажетті минералдар мен дәрумендерге деген сұраныстың артуына байланысты болады. Жиі кездесетін анемия - темір тапшылығынан болатын анемия. Анемия ауруларының 90 % бөлігін темір тапшылығындағы анемия құрайды. Гемоглобиннің құрамында ағзаға қажетті темір элементі болады. Темір ионы қанның құрамындағы гемоглобиннің негізгі құраушысы болып табылады. Ол оттегі тасымалдауға қатысады. Егер темір ионы аз болса, гемоглобинде аз болады [1, 2] .
Гемоглобин аз болса, денеге таралатын қанның құрамында оттегі аз болады. Оттегінің аз болуы ағзаны жаймен өлтіре бастайды, өле бастаған ағза ауруға бейім тұрады. Иммунитеттің мықты болмайтындығы да сондықтан. Қаны аз болашақ аналарда жүктіліктің барысында көптеген қиындықтар болады. Мысалы, анемияға шалдыққан жүкті әйелдерде токсикоз 1, 5 есе жиі кездеседі. Анемияға шалдыққан әйелдер арасында гестоздың пайда болу жиілігі 40 пайызды құраса, түсік тастау жиілігінің кездесуі жиілігі 15-42 пайызды құрайды. Босану кезінде түрлі асқынуларға анемия себеп болады. Анемияға шалдыққан әйелден туылған бала өмірге келгенде алғашқы жылдарында анемияға жиі тап болады. Одан бөлек, түрлі ауруларды жұқтыруға бейім келеді атап айтсақ, тыныс жолдарындағы вирустық аурулар жиі кездеседі [4, 5, 6] .
Антиоксиданттар клетка құрылымын бос радикалдардың зақымдануынан сақтап, жалпы клетка қорғауда үлкен рөл ойнайды. Сондықтан да витамин жиынтықтарынан құралған биологиялық белсенді препараттарды қоршаған ортаның зиянды әсерінен қорғаудың үлкен ғылыми және практикалық маңыздылығы бар [7] .
Жұмыстың мақсаты: анемияға ұшыраған буаз және сүт түзуші егеуқұйрықтардың сүт безі микросомаларындағы липидтердің асқын тотығу үдерісі мен антиоксиданттық жүйесіне биологиялық белсенді заттардың әсерін зерттеу.
Жұмыстың мақсатына байланысты келесі міндеттер қойылды:
1. Буаз және сүт түзуші егеуқұйрықтардың сүт безі микросомаларындағы липидтердің асқын тотығу үдерісіне анемияның әсерін зерттеу.
2. Анемияға ұшыраған ұрғашы егеуқұйрықтардың қанында Е және С витаминдерінің мөлшері мен каталаза белсенділігін анықтау.
3. Анемияға ұшыраған буаз және сүт түзуші егеуқұйрықтардың сүт безі мембраналарының төзімділігі мен қан құрамындағы Е және С витаминдерінің мөлшеріне және каталаза белсенділігіне биологиялық белсенді заттардың әсерін зерттеу.
- Әдебиетке шолуЛактация кезіндегі сүт безінің секреторлы клетка мембранасына сипаттама
Сүттің түзілуі - сүт бөлетін жануарлардың барлық жүйесі қатысатын көп кезеңді процесс. Безді ұлпалар көптеген альвеолдардан тұрады, олар сауу бөлігінің жоғары бөлігінде төмен қарай тармақталған цистернаға жанаса орналады. Альвеолалар диаметрі 0, 1-0, 4 мм болатын тұйықталған көпіршіктерден тұрады. Альвеолалардың қабырғасы ішкі жағынан бос ұшы алвеола қуысына бағытталған секреторлы клетка қабаттарымен (эпителиалды) жабылған. Базальды мембрана бұл клетканың негізі болып табылады, ол арқылы қаннан сүттің негізгі ізашары сіңіріледі, ал альвеола қуысына қараған жоғарғы бөлігіндегі альвеол саңылауларына секрет бөлінеді.
Сүт безінің негізгі құрылымды-функционалды бірлігі сүт түзеді, оны альвеол деп атайды. Альвеолалар бөліктерге бөлінген, сырт жағынан қарағанда бір шоқ тәрізді, ал бөліктер өте ірі бөлшектерге бөлінген. Сүт безінде әдетте 15-тен 25-ке дейін бөліктер болады. Альвеола сүттің барлық құрамды компоненттерін өндіретін (белок, май, қант) секреторлы эпителидің моноқабатты клеткасынан тұрады. Альвеола сыртқы жағынан базальды мембранамен қапталған, ол клетканың жоғары бүйірлі тығыз қосылыстарымен бірлесе отырып қабырғалардың таңдамалы өткізгіштігін анықтайды және гемато-сүтті тосқауылды құрады. Альвеолалардың қысқартылатын аппараты - миоэпителиалды клеткалар - секреторлы клеткаға тығыз жанасқан сирек тор құрады. Миоэпителиалды сүттің азаюы кезінде альвеоладан сүттің ағымы жіберіледі. Сүттің ағымы өте мол, ол сүтті қорда сақтау қызметін атқарады. Сүттің түзілуіне бір клеткада жүретін үш түрлі секреторлы процестер қатысады. Белок синтезі рибосомада жүреді. Сонымен, белок молекулаларын құрайтын амин қышқылдары хабарлы РНҚ мини-бағдарламасында жазылған жүйе арқылы құрастырылады. Ішкі клеткалық құрылымдардың жұмыс істеуі үшін энергияны митохондрияға жеткізеді.
Клетка ішінде сүт белогын яғни секреторлы өнімдердің ауысуын қамтамасыз ететін клетка ішілік каналдар (эндоплазмалық ретикулум) жүйесі болады. Рибосомада синтезделетін белок тізбегі канал кеңістігінің ішкі жағына тартылып, Гольджи комплексіне түседі. Дәл сол жерде болашақ концентрация және секреторлы өнімнің "жиынтығы" пайда болады. Бір уақытта Гольджи комплексті құрылымында лактоза синтезделеді.
Тегіс эндоплазмалық торда сүт майларының кішкене бөлшектері синтезделеді. Ұсақ тамшылар ірі болып біріктіріледі; ірі май тамшылары плазмалық мембранаға жақындайды. Мембрана май тамшыларын бүркемелей отырып, тарылады және қайта байланады. Май тамшыларын ұстап қалатын жұқа бөгет үзіледі де май альвеол қуысына түседі. Бір уақытта мембранаға белок пен лактозадан тұратын көпіршіктер жақындайды. Көпіршік мембранасы плазмалық мембранамен бірігеді және көпіршік қуыстары толығымен ашылады да оның құрамындағы заттар алвеолаға босатылады (1 сурет) .
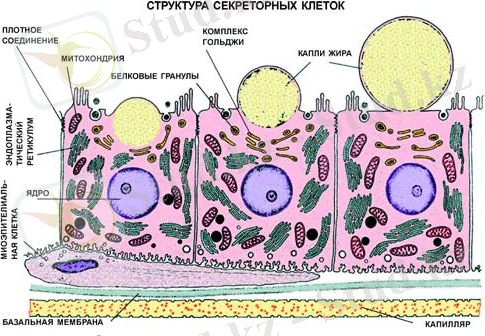
Сурет 1. Сүт безі секреторлы клеткасының құрылысы
Сүт безі секреторлы клеткаларының жіктелуіне тән ерекшеліктер, олардың құрылымы мен тинкториалды қасиеті бойынша ажыратылатын төбелік және базальдық аймақтарға бөлінуі болып табылады. Секреторлы клетка тіршілігіндегі өзгешелік осы клетканың құрылымдық-функционалдық поляризациясымен анықталады, ол «клетка ішілік секреторлы конвейердің» типтік ұйымдастырылуы болып табылады және секреторлы өнімдердің алмасуына сәйкес келеді [5, 6] .
Сүт безі секреторлы клеткалары мембранасының барлық жоғары бетінде негізгі лактогенді гормондардың - пролактин, окситоцин рецепторлары орналасқан. Сүт безі секреторлы клеткаларының плазмалық мембранасының құрылымында заттарды трансмембраналық тасымалдау тәрізді маңызды қызметін көрсететін арнайы функционалдық ерекшелік болады.
Сүт безі секреторлы клеткаларының мембранасындағы төбелік, базалды және базолатералды аймақтарының құрылымы бойынша айырмашылықтары болады. Төбелік плазмалық мембрана цитомембраналық жіктелу есебінен клеткалардан май тамшыларының бөлінуі процесі кезінде үнемі жаңарып отырады, нәтижесінде төбелік мембрана Гольджи аппаратының везикуласымен толықтырылып отырады. Демек, гепотоцит мембранасымен салыстырғанда мембранадағы майлы шариктер, ганглиозидтер санының аз болатындығымен түсіндіруге болады. Төбелік мембрананың құрылысы латералды және базалды аймақтарға қарағанда белгілі-бір айырмашылықтары болады.
Базалды және базолатералды аймақта интегралды белок - тасымалдаушы Na, K- АТФаза орналасқан, ол сүттің ізашарын - заттарды белсенді трансмембраналық тасымалдау арқылы жүзеге асырады және рецепторлы жүйе арқылы үнемі клетка ішілік иондармен қамтамасыз етеді. Сүт безі секреторлы клеткаларының мембранасында кальции насосы ретінде әсер ететін Mg 2 - тәуелді Ca 2+ - АТФаза болады және қанда Ca 2+ ионы белсенді түрде тасымалданады. Ca 2+ ионы алвеолада сүттің экструзия процесін жүзеге асыруы үшін қажетті сүт козейнді белогын ірі мицелииге жинақтау үшін қажет болып табылады. Лактация белсенділігінің деңгейі неғұрлым жоғары болса, соғұрлым клетка ішіндегі Ca 2+ мөлшері де артады. Ca 2+ ионының концентрациясы сүтте 30 мМ жетуі мүмкін [6] .
Секреторлы клетканың базальды мембранасы капиллярлардың миоэпителиалды мембранасымен және клетка ішілік байланыстырушы құрылымдармен тікелей байланысты, нәтижесінде мембрана арқылы тасымалдау қызметін атқаратын тығыз гистогемотологиялық тосқауыл (барьер) түзіледі, ол сонымен қатар ірі молекулалар мен бактериалар және т. б. үшін «молекулалық сит» болып табылады. Базальды мембранаға «өткізу қабілетін» арттыратын қатпарлар, цитоплазма ішілік каналдар тән [7] .
Тірі организм клеткаларына тән ерекшеліктер мембрананың бар болып табылуында. Мембрана клетка ішілік заттарды қоршаған ортасынан ажыратады және мембраналық қосқабаттың екі жағына да органикалық және бейорганикалық заттарды ассиматериялық жолмен таратуды қамтамасыз етеді, сондай-ақ химиялық заттардың сигналының мөлшерін байқауға мүмкіндік беретін рецепторлардың көмегімен сыртқы ортадағы хабарламаларды қабылдайды, мысалы, гормондарды айтуға болады. Биологиялық мембрана бөгде және өзіндік токсиндерді бейтараптандыра отырып, иммунитетті қамтамасыз етеді. Биологиялық мембрананың әртүрлі қызметінің тиімді атқарылуы мембраналық компоненттерінде түзілетін бірегей қасиеттеріне байланысты болуы мүмкін.
Қазіргі кездегі мембрана құрылымының негізінде сұйық-фазалы концепция жатыр. Осы ұсынысқа сәйкес қос қабат сұйық құрылымды болып табылады, онда түзілетін липид молекулалары әртүрлі қозғалыстарды жүзеге асыруға қабілетті: сегменталды қозғалыс, айналмалы қозғалыс және латералды диффузияны жатқызуға болады. Аз жылдамдықта олар қос қабаттың екінші бетіне өтуге және шығуға қабілетті.
Дәл осы липидтер қос қабатты матрикс құра отырып мембрананың тұтас құрылымын анықтайды. Мембрана липидтері құрылымдық элемент ретінде ғана емес, сонымен қатар ферменттік катализ, рецептор және өткізгіштікті реттеуші ретінде мембрананың жұмыс атқаруына қатысады.
Жануарлар клеткасының мембранасында май қышқылы, сфингозин және глицериннің туындысы - фосфолипидтер басым болады. Осы қосылыстардан басқа фосфолипид молекуласының құрамына фосфор қышқылының қалдығы және молекулалардың полярлы басын түзетін қосымша гидрофильді радикалдар кіреді. Фосфолипидтер полярлы басының құрылымы бойынша ажыратуға болады. Алайда молекулалардың гидрофобты бөлігіне кіретін май қышқылдары тізбегінің жиынтығында да айырмашылықтар бар. Олар қаныққан ацилді тізбекті немесе бір-екі қос байланысты тұтастай қамтамасыз етуі мүмкін. Біршама жиі кездесетін 16-көміртеткті қаныққан пальмитинді май қышқылы, 18-көміртекті қаныққан стеаринді және С 9 көміртек атомының тізбегінде қос байланыс болатын 18-көміртекті қаныққан олеин қышқылы. Линолды, арахидонды және дезоксигександы қышқылдар аз кездеседі. Қаныққан май қышқылдары фосфатидилинозит және фосфатидилглицеринге өте бай. Барлық фосфолипид құрамындағы қаныққан май қышқылдары көпшілігінде қаныққан цис-конфигурацияны басытқылайтын созылыңқы транс-конфигурация түрінде болады. Осы себепті фосфолипид молекулаларының қаныққан және қанықпаған май қышқылдарымен бірігуі қос қабаттың бейімді құрылымын және тығыз қабатын бір уақытта қамтамасыз етеді.
Мембрана липидтерінің құрамына стериндер де кіреді. Жануарларда бұл холестерин. Холестерин және оның аналогы қос қабаттағы фосфолипидтердің тығыз қабатын бақылап отырады. Сондай-ақ биомембрана құрамына гликолипидтер, цереброзидтер, сульфатидтер және ганглиозидтер кіреді. Бірақ жануарлар мембранасында липидтердің соңғы келтірілген класы аз мөлшерде кездеседі. Гликолипидтер мембрананың спецификалық қызметінде, әсіресе нерв клеткасында өте маңызды болып саналады. Олар нерв клеткаларының бөліну процесін бақылап отыратындығы дәлелденген.
Мембрананың барлық липидтері бірыңғай құрылымды болады: олардың құрамына туыстығы жағынан суға жақындығы аз және липофильділігі өте жоғары ұзын тізбекті көміртек қалдықтары және гидрофилді полярлы топ - полярлы басы кіреді. Мембранадағы липидтер молекуласы нақты ұйымдастырылған түрде болады: май қышқылды гидрофобты «ұшы» қосқабаттың ішкі жағына бағытталған, ал полярлы «басы» гидрофильді ортаға бағытталған [10] .
Нәтижесінде осындай молекулалар бір-бірімен тәртіп бойынша қарым-қатынас жасайды. Цис-конфигурацияда иілген май қышқылдарының «ұшының» болуынан түзіліс құра алатын конструкция борпылдақ болады және көршілес жатқан фосфолипид молекулалары арасындағы саңылаулар арқылы стреин (холестерин) молеулаларына еніп кетуі мүмкін, ол қабатты тығыздап, гидрофобты қарым-қатынасты қарқындандыра түседі. Қос қабаттағы липид молекулаларының ұйымдастырылуы олардың жылдамдығын шектейді және молекулаларды ретке келтіріп, олардың қатынасын арттырады, бірақ жылдамдығын мүлде тоқтатып тастамайды. Белоктар қос қабатқа еніп, оны модификациялайды. Әртүрлі мембраналардың құрамына кіретін біршама белоктар бар. Белоктар мембрана қос қабатына спецификалық қасиет береді. Қандай да болмасын белоктың болмауы мембрананың функционалдық белсенділігіне әсер етеді.
Белоктар лабилді болып табылады. Белоктың латералды қозғалысы қасиетіне байланысты ғана анықталмайды, ол сонымен қатар липид айналасындағы микротұтқырлыққа және липидтердің фазалық күйіне байланысты анықталады. Осылайша белок молекулаларының қозғалысы және олардың мембранадағы ассоциациясы липидтермен бақыланады.
Мембранамен қатынасу ерекшеліктеріне байланысты оларды екі дәрежеге бөледі: липидті қос қабатпен тығыз қатынастағы интегралды белоктар және белоктың біреуімен ғана қатынасқа түсетін - перифериялық белоктар. Перифриялық белоктардың интегралды белоктардан айырмашылығы, олар қос қабатқа тереңірек енбейді және әлсіз белок-липидті қатынастан тұрады [11] .
Перифериялық белоктар фосфолипидтердің ацилді тізбегінің қозғалысына, күйіне анағұрлым аз әсер етеді. Олар әлсіз әсерлер кезінде яғни осмостық шок, ортаның қышқылдығы және Са +2 ионының концентрациясы өзгергенде мембранамен байланысты жоғалтады [8] .
Мембрана белоктарының функционалдық белсенділігі олардың жоғары конформациялық лабилділігімен анықталады, оның негізінде түскен салмақты босаңсуға немесе керісінше босаңсудан салмақ түсуге конформациялық өткізуді жүзеге асыратын белок молекулаларының қабілеті жатыр. Мембранадағы белоктарды конформациялық өткізу энергиясы мембрана липидтерінің тығыз қабатына (микротұтқырлық) және белокты қоршаған аннулярлы липид молекулаларының жағдайына тәуелді.
Қос қабаттың барлық қабатына енетін перифериялық белоктар оның екі қабатының біреуінің эластикалық деформациясын туғызады. Дәл осындай өзгерістер гормондар мен мембрана рецепторларының қарым-қатынасын жеңілдетеді.
Мембранада көміртек молекулалары бос күйінде кездеспейді, бірақ олар белок және липидтермен комплексті түрде кездесіп, әртүрлі қызмет атқарады [9] .
Биологиялық мембраналарға тән ерекшеліктер қос қабаттың екі жағында да липидтер құрамының әртүрлі болып табылатындығында. Сонымен қатар қос қабат асимметриясы липидтік зат алмасудың ферменттерімен және липидтерді тасымалдайтын белоктармен қамтамасыз етіледі. Қос қабат ассиметриясын құрайтын және оны қолдайтын ферменттерге липаза, холестерин алмасу жүйесі және метилаза жатады. Клетка мембранасының белокты, липидті және көміртекті құрамы клетканың барлық тіршілік циклінде динамикалық тепе-теңдік күйінде болады. Бұл тепе-теңдік эволюцияны ерте бағындыру болып табылады. Бірақ бейімделу процесі кезінде оң немесе патологиялық сипатқа ие болатын икемделу өзгерісі пайда болады, нәтижесінде мембрана құрылымын едәуір айқындалған өзгеріске ұшырататын жекелеген компоненттер құрамының өзгеруін тудырады [8] .
Мембрананың бұзылуы оны құраушы бөлiктердiң, негiзiнен липидтер мен белоктардың ацидоз кезiнде белсендiлiгi жоғарылайтын лизосомды ферменттер әсерiнен зақымдануына, қуаттың жетiспеушiлiгiнен мембрана липидтерi мен белоктарының түзiлуiнiң төмендеуiне және клетка мембранасы липидтерiнiң асқын тотығуының артуына байланысты болуы мүмкiн. Әртүрлi ұлпалардың клеткалары липидтерінiң асқын тотығуының шектен тыс жоғарылауы мембрананың бұзылуы мен энергияның жетiспеушiлiгiне, иондардың таралуы мен клеткадағы ферменттердiң зақымдалуына, клетка мембранасы құрылымы мен әртүрлi заттар үшiн мембрана өткiзгiштiгiнiң өзгеруiне, мембранада орналасқан рецепторлардың сезгiштiк қасиетiнiң жойылуына алып келедi [9] . Оттегі тапшылығынан клетка құрамында қайта қалыптасқан қосылыстар жоғарылайды және осыған байланысты О 2 түзілу мүмкіншілігі артады. Тотығу процесі кезінде олардың өнімдерінің жоғарылауы немесе клетканы қорғау механизмдерінің өзгеруі себепті тотыққан метаболиттердің зияндылығы артады [10] . Мембрана белоктарының, сондай-ақ рецепторлар қызметтерінің бұзылуы мембрананың липидтік құрамының арақатынасындағы өзгерістерге, мембрана липидтерінің құрамына кіретін қанықпаған май қышқылының артуы немесе төмендеуіне, асқын тотықтардың жоғарылауына себепшi болады. Осылардың салдарынан биомембраналарда патологиялық өзгерiстер туындайды. Патологиялық ауытқу мембрана қызметінің тізбекті өзгерiстерiне алып келедi [11] .
Әр түрлi мүшелердiң клетка мембранасында липидтердiң асқын тотығу процестерiнiң жүруі жоғарылайтындығы анықталды. Бұған мембранасынадағы ацилгидроасқынтотығы, диенді конъюгаттар, малонды диальдегидтер тәрізді липидтердің ыдырау өнімдерінің жоғарылауы мен супероксиддисмутаза, каталазаның жоғарылап, глутатионредуктазаның төмендеуі сияқты антиоксиданттық ферменттер белсенділігінің өзгеруі дәлел болады [11] .
Сондай-ақ, липидтердiң асқын тотығу процесі клеткаларда қалыпты жағдайларда да жүреді. Олар микросомдық оксигеназаға тәуелді НАДФ(Н), циклооксигеназа мен липооксигеназа сияқты көптеген ферменттермен реттеледі. Мұндай бос радикалды реакция радикалдарды қосып алатын қабілеті бар антиоксидантты заттар (токоферолдар, витаминдер) көмегімен немесе антиоксиданттық жүйе ферменттерiнiң (супероксиддисмутаза, глутатионпероксидаза, глутатионредуктаза, каталаза, т. б. ) белсендiлiгiнің жоғарылауы нәтижесiнде басытқыланады [12] .
Биологиялық мембрана қалпының бұзылуына алып келетін ферменттердің құрылымы мен қызметінің өзгеруі, белоктар, нуклеин қышқылдары, липидтер тәрізді макромолекулалардың қайтымсыз өзгерістері стресс факторлардың молекулалық деңгейдегі негізгі зақымдаушы әсерлерi болып табылады.
1. 3 Организмдегі липидті асқын тотық пен антиоксиданттық жүйелер және мембраналардың физиологиялық жағдайы
Көптеген әдебиеттерде анемияның организмге әсері және сондай-ақ әйелдердің жүктілік пен лактация кезеңіндегі әсері туралы аумақты мәліметтер кеңінен қарастырылған.
ОБФ-ның шектен тыс өнімдерінің көбеюі немесе антиоксиданттардың жеткіліксіздігінен ОБФ (оттектің белсенді формалары) генерацияланып және басқа радикалдардың антиокисданттармен тазартуы күшейіп өгерістер туындайды.
Тәжірибелік анемияның әсері бірнеше зерттеулерде жүктіліктің бала көтеру кезеңіне және ерте түсік тастауға әсерін тигізуі мүмкін дейді. Антиокисданттар (C және E витаминдері сияқты) және атиоксидантты кофакторлар(селен, мырыш және мыс сияқты) пайда болған ОБФ-ын тазартатын, төмендететін байланыстар болып табылады [13] .
Антиоксиданттар әйелдердің репродуктивті жүйесінде маңызды рөл атқарады. Бір қатар зерттеулерде жүктілік кезеңіндегі анемия мен оның асқынулар қарастырылған. Антиоксидантты жағдайының төмендеуі түсік тастауға алып келуі мүмкін [14] .
Вурал және басқалар көрсеткендей аскрабин қышқылы мен α-токоферол аяқ астынан түсік тастаған әйелдерде аса төмен болғанын көрсеткен, плазмадағы антиоксиданттар аса төмен денгейінде болуының нәтижесінде спонтанды түсік тастау пайда болуы да мүмкін [14] . Жүкті әйелдердің плазмасында тиолдың, плацентарлық липидтердің тотықтануының көбеюі және антиоксиданттардың экспрессиясының төмендеуі тіркелген.
Диенді коньюгаттар, асқын тотығудың біріншілік өнімі, ол май қышқылының көмірсу ұштары гидрофобты полярлығын көбейтіп мембрананың липидті қос қабатын құрайды. Физиологиялық үрдіс кезінде клетка белсенділігінің реттелуі полярлығы жоғарылаған көмірсу ұштары мембрананың терең қабаттарынан жоғары қарай ығысып мембрананың өзіндік жаңару үрдісін жеңілдетеді және оның өткізгіштігімен ионды тасымалдануына әсерін тигізеді. Қоршаған ортаның қолайсыз және басқа патологиялық жағдайларында оттегінің бос радикалды формаларының үздіксіз түзілуінен ЛАТ үдейді қанықпаған липидтердің толық бұзылуына, белоктар мен басқа молекулалардың құрылымы мен қызметінің бұзылуына әкеледі және нәтижесінде клетканың тіршілігін тоқтатады [13] .
Малонды диальдегиді (МДА) организмде оттегі реактивті формаларының әсерінен полиқанықпаған майлар бұзылғанда липидтердің асқын тотығуы мен оксидативті стресстің маркері ретінде қызмет атқарады. МДА бірқатар басқа да «тиобарбитур реактивті заттармен» тиобарбитур қышқылымен реакцияға түсіп флуоресцентті қызыл түске боялады, нәтижесінде МДА құрамын спектроморфометриялық талдау арқылы жүргізуге болады.
Глутатионпероксидаза сутек асқын тотығының бұзу нәтижесінде болған әсерінен қорғайды. құрамында селеннің болуы ол, клетканы асқын тотық радикалдардың уланунан сақтайтын маңызды рөл атқарады. Глутатионпероксидаза сутек тотығының құрамын бұзу қызметін атқарады, ферменттіңәсер ету механизмі- құрамында күкірт бар амин қышқылдары мен SH-тобына ие, глутатионнан пайда болған коферменттің болуы.
Тотыққан глутатион глутатионредуктаза НАДФ-Н-ты қолдана отырып, қалпына келеді. Стресс факторларының әсерінен Антиоксиданттық жүйедегі (АОЖ) ферменттік бөлімге әсерін тигізіп, глутатионредуктазаның, супроксиддисмутазаның, және каталаза, глюкоза-6- фазалық белсенділігінде өзгерістер байқалады.
... жалғасыСіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz