Үлкен нейрондық деректерді k-орташа кластерлеу және бас миының визуализациясы

МAЗМҰНЫ
Нейронды желілердегі деректерді өңдеудің әдістерінің
және алгоритмдерінің анализі
КІРІСПЕ
Қазіргі әлемде ақпарат маңызды байлықтардың бірі болып табылады, ал ақпараттық жүйе (АЖ) барлық қызмет саласында қажетті құрал болып танылады.
Ақпараттық жүйенің көмегімен шешілетін міндеттердің әртүрлілігі, құрастырылу қағидасымен және оларға енгізілген ақпаратты өңдеу ережесімен айрықшаланатын көптеген әртүрлі жүйелердің пайда болуына алып келді.
Диссертациялық жұмысымның тақырыбы осы ақпараттық жүйеге қатысты «нейрондық желілерді тереңдетіп оқытуды зерттеу» болып табылады. Бұл жұмыстың мақсаты ақпараттық жүйенің пәндік аймақ ретінде нейрондық желілерді зерттеу болып саналады.
Диссертациялық жұмыстың мақсаты:
- Диссертациялық жұмыста үлкен көлемдегі мәліметтерді кластерлеу мәселелері қарастырыл-ған. Дипломдық жұмыстың мақсаты - k-орташа кластерлеу әдісі арқылы адамның миымен байланысты аурулардың дәл диагнозын анықтайтын тапсырмаларды оңтайландыру.
- k-means Кластерлеу әдісін талдау
Диссертациялық жұмыста қойылатын талаптар:
- диссертация тақырыбына байланысты танымдық белсенділікті көрсету мақсатында мәліметтер жинақтау;
- диссерациялық жұмыстар тақырыптарын анықтау кезінде өндіріс, білім және ғылымның нақты мәселелерін ескеру;
- қорғалып отырған диссертациялық жұмыстың тақырыбын ашу мақсатында, оны жеткізу ретінде зерттеу жұмыстарын енгізу;
- диссертациялық жұмыс тапсырмаға сәйкес перспективті теориялық және практикалық мәселелерді қамту.
Диссертациялық жұмыстың өзектілігі:
- Қазіргі заманғы нейробиология үлкен көлемді және егжей-тегжейлі «үлкен нейрондық деректерді» жеткізетін миды картаға түсіру және жазу технологиясының дамуына негізделген трансформацияның ортасында тұр. Желілік ғылым нейрондық жүйелерді модельдеуге және талдауға, жеке нейрондардан бастап, бүкіл миды қамтитын тізбектерге негізделген принциптердің бірі ретінде пайда болды. Ми желілеріндегі зерттеулер ұлттық және халықаралық конференцияларда, жетекші ғылыми журналдардың арнайы басылымдарында және АҚШ-тың Адамның Коннектом жобасы, BRAIN бастамасы және Еуропалық ми сияқты ми жұмысын түсінуге бағытталған кең ауқымды бастамалардың бір бөлігі болды. Шекті криптожүйелер құпия ақпаратты кеңістікте бөлуді ұйымдастырады, оған сәйкес шекті схеманың n абоненттерінің тек құпияны қалпына келтіре алады. Шекті схемалар ауқымы ақпараттарды жасырын беру жүйелеріне, желідегі компьютерлер арасында деректерді берудің динамикалық тепе-теңдігіне және мәліметтер қорын қорғауға кеңейеді.
- ӘДЕБИ ШОЛУ
1. 1 Brain networks тарихы
«Танымдық сәулет» термині адамның танымындағы ережелерді түсіндіруге күш салған (Фодор мен Пылышин, 1988) когнитивтік немесе компьютерлік ғалымдардың толығымен қолданатын тұжырымдамаларына қатысты (1-жолақты қараңыз) . биологиялық субстрат - адамның миы. Заман өзгерді. Танымал архитектураның жаңа суреті пайда бола бастады, бұл арнайы шығарылымға қосқан үлестерімен дәлелдеді. Көптеген «когнитивтік сәулет» қазіргі кезде белгілі бір міндеттер жиынтығын немесе белгілі бір функцияларды орындауға ықпал ететін ми аймақтарының жиынтығы ретінде қарастырылады. Көбінесе бұл архитектуралар нақты түрде желілер деп аталады, мысалы, әдепкі режимдік желі (Райхль және басқалар, 2001), назар аудару желілері және т. б. (мысалы, (Корбетта және Шульман, 2002) ) .
Алайда, «желі» терминінің мәні өте өзгергіш. Көптеген жағдайларда, желі термині осы аймақтар арасындағы байланыстарға нақты сілтеме жасамай, фМРИ-ге қатысты зерттеулер жиынтығы кезінде іске қосылатын қарапайым аймақтар жиынтығына бейресми түрде қолданылады. Желілердің осы бейресми түсінігіне керісінше аймақтар жиынтығы ретінде осы мақалада қабылданған желіні неғұрлым ресми анықтайды. Желі - бұл жүйенің элементтерінің арасындағы түйіндердің жиынтығын байланыстыратын жиектердің жиынтығы ретінде ұсынылған жүйелік қатынастардың жиынтығы. Нейробиологиялық желілер нейрондардың ішіндегі жасушаларға тән метаболикалық немесе реттелетін жолдардан кортикальды аймақтар мен субортикалық ядролардың өзара әрекеттесуіне дейін масштабтың көптеген деңгейлеріне келеді (1-суретті қараңыз) . Әр деңгейде (нейрондар, нейрондық тізбектер және популяциялар, жүйелер) әр түрлі желілер бар, маңыздылығы әртүрлі. Осы деңгейлердің әрқайсысында жеке элементтердің қалай жұмыс жасайтынын түсіну ғана емес, сонымен бірге элементтерді үлкен өзара байланысқан жүйенің контекстіне енгізетін жұптасқан қатынастар жиынтығын түсіну маңызды (Sporns, 2011) . Кейбір ерекшеліктермен қатар, когнитивті сәулет негізінен осы жоғары деңгейдегі құрылымдар мен механизмдерді қамтиды (Сежновски и Черчилл, 1989) . Осы мақала үшін біз мидың барлық немесе көп бөлігіне қатысты желілерді түсіну үшін осы ең жоғары деңгейлерге назар аударғымыз келеді. Біз осы басылымның басқа мақалаларында қарастырылған нақтырақ архитектураларды қамтитын кең көлемді архитектуралық принциптер мен қасиеттерді зерттегіміз келеді.
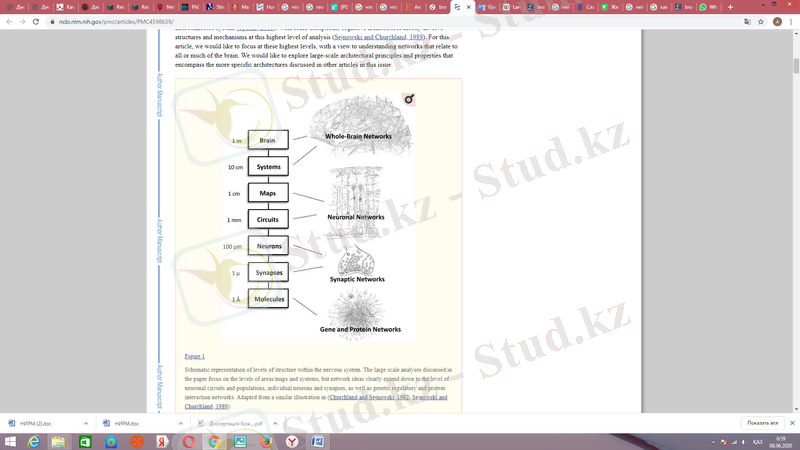
1-сурет, ми деңгейлері
Неврологиядағы желілер туралы түсінік 19-ғасырдың екінші жартысында Вернике, Лихтейм, Лиепман, Дежерин және басқалары негізінен клиникалық-патологиялық корреляциялар негізінде айтылған «ажырату синдромдары» гипотезасының пайда болуымен басталды. Тұжырымдама байланысты салалардағы екі дамуды қолдады: 1) когнитивтік функцияның «ассоциативті» модельдерінің кеңінен қабылдануы (1) адамның миының беткі морфологиясын сипаттаудан ақ заттағы нейроанатомиялық проекция жолдарын тікелей сипаттауға дейінгі қозғалыс және 2) авторы Джеймс және Фрейд . Алайда, ертедегі «диаграмма жасаушылар», өздерінің зияндылары деп атағандықтан, ХХ ғасырдың ортасында миды ұйымдастырудың (мысалы, жаппай әрекет ету) бәсекелес теорияларына негіз қалдырып кетті. Ақпаратты өңдеуді бөлу және интеграциялау үшін құрылған ми желісінің орталық тұжырымдамасы 1960 жылдары Норман Гешвинд ағылшын тілінде сөйлейтін әлемге енгізілгенге дейін қайтадан әсер етпеді . Гешвиндтің мұрасы кортико-кортикальды байланыс ұйымын тереңірек зерттеуге шақырғанмен, инновационды емес бейнелеу әдістері бұл қатынастарды in vivo-да зерттеу үшін әлі кең қол жетімді болмады.
Соңғы бірнеше онжылдықта нейрохимиялық түсірудің көп мөлшерлі емес құрылымдық және функционалды әдістерінің көптігі үнемді болды, ал бұл әдістер үшін деректерді өңдеудің негізгі әдістері жетілдірілді. Сонымен қатар, статистикалық физика саласындағы оқиғалар қазіргі кездегі «күрделі желілік ғылым» деп аталатын жаңа пәнаралық өрістің пайда болуына әкелді, ол әртүрлі күрделі нақты жағдайлардың ұйымдастырылуын және пайда болатын әрекеттерін түсінудің математикалық және тұжырымдамалық құралдарын ұсынады. өмір желілері; қол жетімді кіріспе үшін және техникалық жағдайды жан-жақты қарастыру үшін бөлімін қараңыз. Күрделі желілік ғылымнан пайда болған негізгі түсінік - бұл әр түрлі жүйелер негізгі ұйымдастырушылық принциптерді ортақ қолдана алатындығында. Сонымен қатар, кеңістік пен уақыттың әр түрлі ауқымында болатын ми сияқты күрделі жүйеде желінің қасиеттері жоғары деңгейде сақталуы немесе масштабты өзгермейтіні көрінеді. Мысалы, микроскопиялық ұялы желілер де, нейроэмизинг деректерінен алынған макроскопиялық желілер де модульділік, концентратор түйіндерінің болуы және сымдардың минималды шығындары үшін ақпарат берудің жоғары тиімділігі сияқты изоморфты қасиеттерін көрсете алады; Жалпы нейробиологияға қатысты күрделі желілерді соңғы шолу үшін қараңыз. Бұл мақалада біз адам денсаулығындағы және ауруындағы мидың күрделі желілік қасиеттерін түсіндіру үшін негізінен графикалық теориядан алынған математикалық құралдарды қолдана отырып жасалған соңғы зерттеулерге тоқталамыз.
1. 2. Желілік талдаудың графикалық теориялық түсініктері
Графикалық теорияда желі шеттермен (немесе сызықтармен) байланысқан түйіндер жиынтығы ретінде абстрактілі сипаттамаға дейін азаяды; 2-суретті қараңыз. Жиектерді бағыттауға немесе бағыттамауға болады, ал салмақталған немесе өлшенбеген. Қазіргі кездегі ми желілерінің графикалық теориялық талдауларында басқарылмаған, өлшенбейтін графиктің қарапайым жағдайы қарастырылған.
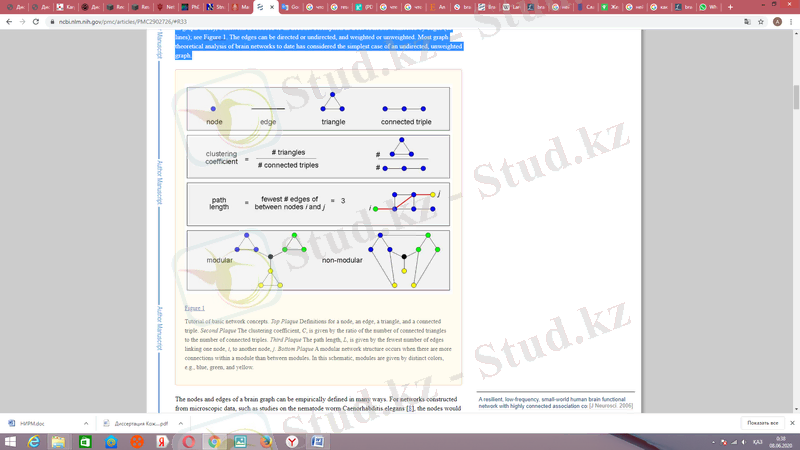
2-сурет, түйіндер жиынтығы
Ми диаграммасының түйіндері мен жиектерін эмпирикалық жолмен анықтауға болады. Caenorhabditis elegans нематодты құрт туралы зерттеулер сияқты микроскопиялық мәліметтерден құрылған желілер үшін түйіндер нейрондар болады, ал жиектер нейрондарды бір-бірімен байланыстыратын аксондар болып табылады. Екінші жағынан, адамның нейрохимиялық мәліметтерінің макроскопиялық масштабында мидың желілерін құру үшін біз түйіндердің негізгі субкортикалық ядролардың және кортикальды аймақтардың және жиектердің арасындағы статистикалық байланыстың, мысалы, корреляцияның немесе өзара ақпараттың байланысы болғандығын көрсете аламыз. аймақтар ; 3-суретті қараңыз.
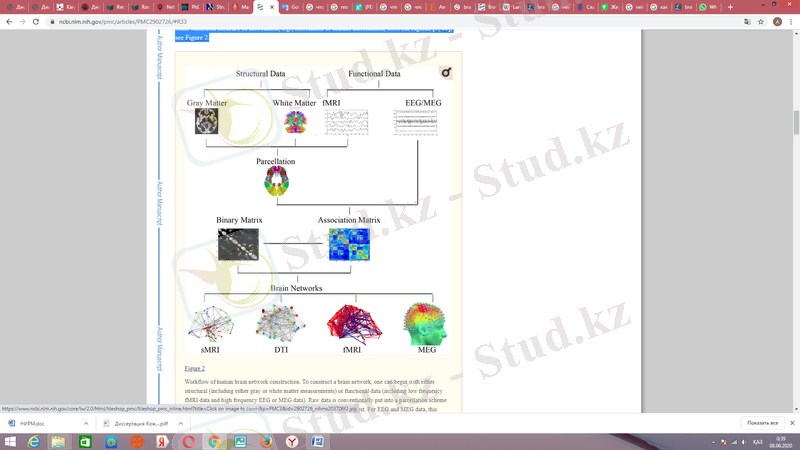
3-сурет, корреляцияның немесе өзара ақпараттың байланысы
Желіні графикалық түрде көрсеткеннен кейін оның топологиялық қасиеттерін өлшеуге болады. Кілттік топологиялық метрика - бұл әр түйіннің дәрежесі, бұл оны желінің қалған бөлігімен байланыстыратын жиектер саны. Желінің барлық түйіндері бойынша дәреженің ықтималдығы бөлінуі дәреженің таралуы деп аталады және көбінесе жоғары дәрежелі түйіндердің немесе желілік хабтардың болуымен үйлесетін көп немесе аз кесілген қуат заңына ие.
Тағы бір кілт метрикасы - бұл бір түйіннен басқа түйінге өту үшін өту керек жиектердің саны; жақын көршілерге арналған жұп түйіндер үшін жолдың ұзындығы 1; Кейде сипаттамалық жолдың ұзындығы деп аталатын барлық мүмкін болатын түйіндердің орташа жолының ұзындығы параллель ақпарат беру үшін желінің жаһандық тиімділігіне кері байланысты. Кластерлеу коэффициенті индекстік түйіннің жақын көршілері арасындағы қосылыстардың тығыздығын өлшейді: жоғары кластерлік коэффициенттер тығыз байланысқан көршілер клетінің бөлігі болып табылатын түйіндерді білдіреді. Ми желілеріндегі осы және басқа өлшемдерді эталондық желілердегі шамалармен салыстыруға болады, мысалы, кездейсоқ желілер (аз кластерлер мен қысқа жол ұзындығы бар) немесе тұрақты торлар (жоғары кластерлік және ұзақ жол ұзындығы) .
Уоттс пен Строгат 1998 жылы өздерінің өте ықпалды мақалаларында қарапайым есептеу моделін қолданып, қарапайым тор мен кездейсоқ желінің шектеулі жағдайлары арасында жоғары кластерлеу мен үйлесімділіктің сипатталатын желілердің аралық режимі бар екенін көрсетті. қысқа жол ұзындығы. Олар әлеуметтанудың алдыңғы бақылауларына сүйене отырып, желілердің бұл класын «шағын әлем» деп атады, олар әлеуметтік желілерде көбінесе жергілікті аудандардағы қарым-қатынастардың жоғары кластерлілігі немесе өзара байланысының ұқсас қасиеттері бар, таңдалған кез-келген екі адамның таңқаларлық қысқа аралық танысу тізбегі бар. үлкен популяциядан кездейсоқ. Бұл талдау нейроциологтар үшін тартымды екендігі дәлелденді, өйткені ол миды ұйымдастырудың локализацияланған және таратылған модельдерінің арасындағы ұзақ уақытқа созылған шиеленісті шешеді: негізінен, шағын әлем желісі жергілікті немесе мамандандырылған аудандарда топологиялық субстратты қамтамасыз ете алады. жоғары кластерлі түйіндер, және қысқа сипаттамалық жолдың ұзындығы бар жоғары тиімді желіде жаһандық таратылған немесе біріктірілген өңдеу . Жақында жүргізілген жұмыс күрделі желілер ғылымынан бастап нейробиология және нейрохимия саласындағы қолданбаларға дейінгі өлшемдердің ауқымын едәуір кеңейтті.
1. 3. Мидың құрылымдық желілері
Адамның миының құрылымдық желілері екі жолмен жасалған: не жанама түрде сұр зат көлемінің аймақтық ковариациясынан немесе құрылымдық магниттік-резонанстық бейнелердегі қалыңдығын өлшеу, sMRI; диффузиялық тензормен, DTI (немесе қатысты әдістермен) қамтамасыз етілген сұр заттың аймақтарының арасындағы ақ заттардың қосылыстарын өлшеуден немесе одан көп. Құрылымдық желілердің тораптары әдетте алдын-ала анықталған анатомиялық іріктеу сызбасының аймақтары ретінде анықталды, мысалы, кортекті шамамен Бродман аудандарына бөлетін автоматтандырылған анатомиялық таңбалау (AAL) шаблон кескіні . СМРИ деректері негізінде сұр заттарды желілер жасау үшін түйіндер арасындағы жиектер аймақтық көлем немесе кортикальды қалыңдық өлшеу арасындағы корреляция күші арқылы анықталады . Мысалы, 100 адамның таңдауында оң және сол жақ фронтальды гирустың кортикальды қалыңдығы арасында тығыз байланыс болса, осы түйіндер арасында жиілік сызықты сызықтық сызықпен сызылады, бұл орташа үлгіні білдіреді. DTI негізіндегі ақ материя желілерінің басты артықшылығы - ықтималды трактография сияқты әдістерді бір тақырыптағы кез-келген аймақтық түйіндер арасындағы байланыс мүмкіндігін анықтау үшін қолдануға болады.
Сонымен қатар, бір тақырыптағы траграфияға негізделген анатомиялық желілер сұр заттардың аймақаралық ковариациясына негізделген желілерге қарағанда, аймақтар арасындағы аксональдық проекцияға тікелей байланысты болып көрінуі мүмкін (дегенмен, осы екі көзқарасты салыстырып тексеру үшін Лерч және басқаларын қараңыз) . . Алайда, DTI негізіндегі желілер үшін қазіргі кездегі күрделі мәселе - трактографияның көптеген әдістері кеңістікте кең бөлінген аймақтар арасындағы байланыстардың ықтималдығын шамадан тыс бағалайды, бұл нәтижесінде пайда болған желілерде алыс қашықтықтардың салыстырмалы түрде таралуына әкеледі.
2. 1 Мидың ірі желілеріне жақындау
Мақаланың негізгі бөлігі екі байланысты зерттеулерде мидың ұйымдастырылуына нақты желілік перспективаны қабылдаудан туындайтын кейбір тұжырымдамалар мен нәтижелерді қарастырады.
Біз алдымен барлық функционалдық белсенділік жүретін анатомиялық желілерді түсіндіруге бағытталған жұмысқа жүгінеміз. Анатомиялық желілер нейрондық сигналдар мен ақпараттың өтуіне кедергі келтіретін қаңқаны қамтамасыз етеді, бұл біздің ойымызды, түсінуіміз бен іс-әрекетімізді қалыптастыру үшін өте маңызды.
Ми желісін зерттеудің екінші бір негізгі әдісі - бұл функционалды MRI BOLD сигналының корреляцияланған ауытқуы (фл. Қуат және басқалар, 2014) . Бұл зерттеулер көбінесе «функционалдық қосылыс» деп аталатын жағдайды құратын нақты міндеттерсіз осы корреляцияны бақылайды. Бұл жұмыс маңызды демонстрациядан басталды, тіпті тыныштықта да fMRI BOLD сигналының ауытқуы мидың анатомиялық спецификалық жолдарымен корреляцияланады. Мысалы, мотор функциясына қатысты көптеген аймақтар ешқандай міндет болмаған кезде бір-бірімен тығыз байланысты. RSFC ұйымы мотор жүйесінен тыс көптеген ми аймақтары арасындағы ортақ функционалды қатынастар туралы түсінік беру үшін көрсетілді. Мақаланың екінші негізгі бөлімінде осы зерттеулер ұсынған кейбір негізгі бақылаулар мен қасиеттер қарастырылған.
Қорытынды бөлімде біз когнитивтік архитектураның негізін құрайтын биологиялық механизмдерді түсіну үшін маңызды деп санайтын құрылымдық және функционалды желілердің өзара байланысын зерттейміз (2-жолақты қараңыз) . Жақында жүргізілген жұмыс мидың осы екі түрінің арасындағы кейбір қатынастарды ашты, құрылымдық байланыстардың функционалды желілерді қалай шектейтіні және көптеген шектеулер бірнеше уақыт шкалаларында қалай ойнайтындығы туралы толық түсініксіз болып қала береді. Құрылымы мен функциясы бойынша желілерді интегративті зерттеу болашақ үшін маңызды мақсат болып табылады, және біз мақалада осы жолдың кейбір қадамдарын диаграммамен аяқтаймыз.
2. 2 Функциялық коррелиция желілері
Кіріспеде айтылғандай, мидың ауқымды желілеріне бір тәсіл - бұл RSFC қолдану. Қазіргі уақытта, осы «айқын емес» жағдайлардағы корреляциялар нені білдіретіні туралы көптеген адамдар қатты алаңдаушылық білдіруде. Алайда, RSFC бақылауының топтық және жеке деңгейде жоғары сенімділік пен репродуктивтілік деңгейін көрсететіні күмән тудырмайды. Шынында да, RSFC желілік деректерін көптеген бақылаулар «тыныштықтың» әр түрлі түрлерінде сақталады (көздер ашық және көздері жабық (МакАвой соавт. 2012), жеңіл анестезия, ұйқының ерте кезеңдері (Horovitz соавт, 2009 ж. ) ; Larson-Prior және басқалар, 2011) және т. Б. ) . Сонымен қатар, жалпы желілік қатынастардың көбісі міндеттер күйінде сақталатын көрінеді (Коул және басқалар, 2014) . Соған қарамастан, міндеттер тынығу кезінде болатын негізгі желілерде тыныштықты тудырады (Коул және басқалар, 2014; Дэвисон және басқалар, 2015) және терең ұйқы және терең анестезия (Гейн және басқалар, 2012) сонымен қатар функционалды бұзылуларды тудырады. сәулет сәулеті.
Осылайша, RSFC тербелістерінің нені білдіретіні туралы нақты сұрақтар туындайды. Олар негізгі анатомиялық қатынастармен шектелген сияқты, бірақ жалпы олар анатомиялық қатынастарды қайталамайды (келесі бөлімді қараңыз) . Ми аймақтары арасында өте күшті функционалды корреляцияларды табуға болады, олар ешқандай тікелей (бір сатылы) анатомиялық байланыспен байланысты емес. Мысалы, сол жақ мотор кортексі мен оң жақ ми қантамырларының арасында функционалды байланыстарды табуға болады (Бакнер және басқалар, 2011), анатомиялық тұрғыдан бір-бірінен бірнеше қадам алыс орналасқан екі құрылым (сурет 4) . Сол жақ және оң жақ жарты шарлардағы бастапқы визуалды кортекстің эксцентрлік көріністері, сонымен қатар, тікелей анатомиялық байланыстарсыз да айқын байланысты (Винсент соавт., 2007) .
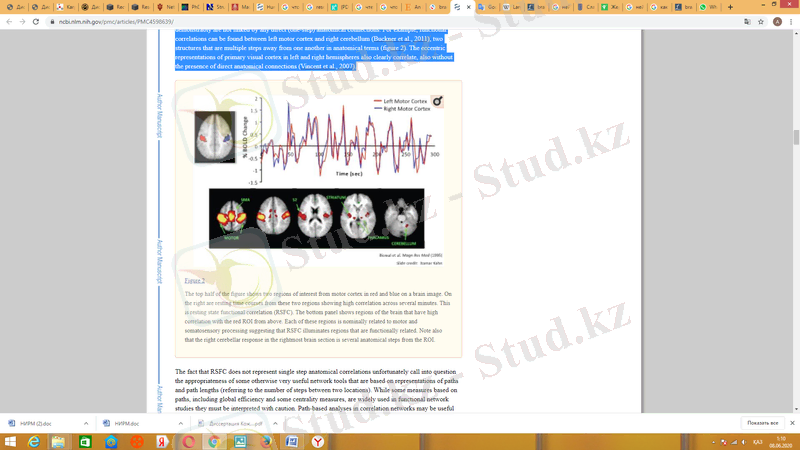
4-сурет, ми құрылымдары және RSFC тербелістері
RSFC бір сатылы анатомиялық корреляцияны көрсетпеуі, өкінішке орай, жолдар мен жолдардың ұзындығына негізделген (екі орналасу арасындағы қадамдар санына негізделген) кейбір басқа өте пайдалы желілік құралдардың орындылығын күмән тудырады. Функционалды желіні зерттеуде кең жолақты тиімділік пен центрлік шараларды қоса алғанда, жолдарға негізделген кейбір шаралар кеңінен қолданылғанмен, оларды абайлап түсіндіру керек. Корреляциялық желілердегі жол негізіндегі талдаулар мидың функционалды жүйелері арасындағы иерархиялық қатынастар мен көп сатылы бірлестіктерді бұзу үшін пайдалы болуы мүмкін. Алайда, ақпараттық ағынға немесе функционалдық қосылыстар бойымен байланысқа тікелей сілтеме жасайтын интерпретация желілері негізгі анатомиялық желілердегі корреляциялық желілерге қатысты күрделі және жанама түрде жүзеге асырылуы мүмкін. Нейрондық сигналдар анатомиялық жолдармен өтеді, нәтижесінде функционалды байланыстар пайда болады.
Демек, функционалды байланыстарды мидың екі аймағының тікелей байланысы ретінде түсіндірудің орнына, RSFC корреляциясы екі аймақтың арасындағы көптеген немесе барлық анатомиялық жолдардағы функционалдық қатынастардың құрама (мүмкін салмақталған сомасын) құрайды деп болжауға болады. , мысалы қарау үшін (Power et al., 2014) қараңыз. Маңыздысы, бұл корреляциялар анатомиялық қатысуды ғана емес, сонымен қатар осы байланыстардың синаптикалық тиімділігін де көрсетеді. Сонымен, функционалдық қатынастар жолдардың құрылымдық орналасуы нәтижесінде ғана емес, сонымен қатар осы жолдар бойындағы синаптикалық тиімділіктің өзгеруі нәтижесінде де өзгеруі мүмкін. Мысалы, тапсырманы көп рет орындау жалпы белсендірілген аймақтар арасындағы байланысты арттыруы мүмкін, мысалы. (Льюис және басқалар, 2009; Макей және басқалар, 2011) . Сонымен мидың жалпы корреляциялық құрылымы туралы ойланудың қызықты тәсілі - бұл анатомиямен шектелген (бірақ толық анықталмаған) аймақтар арасындағы тарихи коактивацияның жоғары деңгейдегі статистикалық көрінісі.
RSFC көмегімен алғашқы зерттеулер тұқымға негізделген әдісті қолданды. Бұл әдіс тұқымдық немесе аралас воксельді жинап алып, мидағы қалған барлық воксидтердің тұқыммен қалай байланысқанын қарастырды (Бисвал және басқалар, 1995) . Көптеген жағдайларда корреляциялық құрылым функционалды түрде байланысты аймақтарды білдіретін сияқты. Айта кету керек, 2003 жылы (Greicius соавт., 2003) жүргізген зерттеу әдепкі режим желісіне байланысты кейінгі цингуляция аймағында тұқым орналастыру әдепкі режимді қосқан аймақтарға ұқсас болатын корреляциялар жиынтығын анықтағанын көрсетті. сол жақ мотор кортексінің қосылыстарын себетіндіктен, мотор жүйелерінің көп бөлігі ашылады. Әдеттегі режим желісі, егер субъект әр түрлі белсенді тапсырмалар күйіне енетін болса, олардың белсенділігінің төмендеуінің ерекше қасиеті бар аймақтар жиынтығы ретінде сипатталған. Бұл зерттеу ұқсас нәтижелері бар қосымша зерттеулердің ағынын ашты. Әр түрлі тұқымдарды қолдана отырып, назар аударудың әр түрлі жүйелері анықталды, және бұл желілер демалудың корреляциялық заңдылықтары тапсырмалар кезінде коактивация заңдылықтарын ұстанатынын көрсетті.
2. 3 Нейрон желiлерiнің жасанды интелектпен байланысы
Процедура арқылы нейрон желiсiнiң керi таратуын үйрену алгоритмі қарастырылған алдыңғы тарауда басқа дәл осылай және мақсаттық шығыс түр кiретiн және желi көрсететiн қайсыбiр сыртқы буынның бар болуын түсiндіредi. Ұқсас концепцияны қолданатын алгоритмдар оқытушымен үйренудiң алгоритмдары деп аталады. Олардың табысты функционалдауы үшін алдыңғы сатыда әр кіріс бейнесіне эталонды шығысты құратын сарапшылардың болуы қажет.
... жалғасыСіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz