Хлоропласттар: құрылысы, қызметі және фотосинтез өнімдері

Қазақстан Республикасының Ауыл шаруашылығы Министрлігі
Қазақ Ұлттық Аграрлық Зерттеу Университеті
« Агробиология» факультеті
«Өсімдік қорғау және карантин» мамандығы
РЕФЕРАТ
Тақырыбы: Хлоропласттар. Фотосинтездің өнімдері
Орындаған: Бахтыгизова Айгерім
Тобы: ЗР-107Қ
Қабылдаған: Мусапиров Д. А.
Алматы 2020ж
Пластидтер
(грек тілінен
plastides
- жасайтын, тузейтін) - эукариотты өсімдік жасушасының органоидтары. Әрбір пластидта кос мембраналық құрылысы бар. Саңырауқұлақтар мен кейбір балдырларда болмайды. Олар пішіні, мөлшері, құрылысы мен қызметтері бойынша әртүрлі. Түсі бойынша жасыл пластидтер (хлоропласттар), сары-ашық қызыл және қызыл (хромопласттар) және түссіз (лейкопласттар) ажыратылады. Солардың ішінде қарастыратынымыз -
хлоропласттар
, яғни
жасыл пигментті пластидтер
. Хлоропласт балдырлардың талломдарындарында, жоғары сатыдағы өсімдіктердің жапырағында, жас сабақта, піспеген(шикі) жемістердің клеткаларында болады. Хлоропласттардың мөлшері жоғары сатыдағы өсімдіктерде 2-7 мкм, қалыңдығы 1-2 мкм болады. . Хлоропластарды ішкі және сыртқы мембраналар қоршап жатады, олардың, қалыңдьғы 7 нм. Тірі хлоропласт құрамында 30% ақуыздар, 33% липидтер, 5-10% хлорофиллдер, 1-2% каротиноидтар және аз мөлшерде РНҚ мен ДНҚ болады. Хлоропластардың матриксында нуклеин қышқылдары (ДНҚ, РНҚ) және рибосомалар орналасады. Хлоропластардың рибосомалары ақуыз синтезін жүзеге асырады. Митохондриялар мен пластидтер - бұл
жартылай автономды (дербес) органоидтер
. Өзінің генетикалық ақпаратын жүзеге асыра алатындықтан осылай аталған. Яғни олардың құрамында өзінің ДНҚ-сы, РНҚ-ның қажетті барлық түрлері және рибосомалар бар. Олар өздерінің ДНҚ-сында жазылған тұқымқуалаушылық ақпарат бойынша нәруыздарын синтездеуге қабілетті. Зерттеулер көрсеткеніндей, митохондриялар мен пластидтерде сақинатәрізді
ядродан тыс
ДНҚ
болады. Бұл ұғымды ядролық тұқымқуалаушылыққа немесе хромосома ДНҚ-сына қарсы
цитоплазмалық ДНҚ
немесе
цитоплазмалық тұқымқуалаушылық
деп атау қабылданған.
Жартылай автономды (дербес) органоидтер ДНҚ-сының молекуласы бактерия ДНҚ-сына ұқсайды. Ядро хромосомасынан айырмашылығы митохондриялар мен пластидтердің ДНҚ-сы сақинатәрізді пішінді және нәруыздармен (гистондармен) байланыспаған. Олардың ДНҚ-сының құрамына осы органоидтің тіршілік әрекетін қамтамасыз етуді талап ететін көптеген нәруыздар мен РНҚ кодталған. Бірақ өздерінің рибосомаларының қалыпты түзілуі мен қызмет етуі үшін пластидтерге ядро хромосомасында кодталған рРНҚ-ның бірнеше молекуласы керек. Яғни жартылай автономды органоидтер мүлде жасушасыз әрекет ете алмайды. Олар химиялық құрамы цитоплазмаға ұқсайтын, бірақ көбейе алмайтын қоректік ортада тіршілік етуге қабілетті. Жартылай автономды органоидтердің рибосомалары едәуір ұсақ және бактерияларға ұқсайды.
Жоғары сатыдағы өсімдіктерде хлоропласт пішіні дөңгелек немесе сопақша, линза тәрізді көбінесе дәнге ұқсас болып келеді. Сондықтан, олардың клеткаларындағы хлоропластты хлорофилл дәндері деп, ал балдырлар клеткасындағы хлоропластты хромотофор дейді. . Көп уақытқа дейін пластидтердің структурасы біркелкі / гомогенді/ болады деп келген еді.
1903 жылы К. А. Тимирязев хлоропластың құрылысы бірыңғай емес, гетерогенді гранулды екендігін эксперимент арқылы дәлелдеді. Пластидті қант ерітіндісіне салып және оған аздап су қосып отырғанда хлоропластың сыртқы жасыл қабаты ортаңғы түссіз қабатынан ажырағанын байқаған. Тимирязев хлорофилл хлоропластың ішінде емес, оның сыртқы қабатындағы гранулда бірыңғай таралған деген қорытындыға келген. Кейіннен А. А. Табенцкий /1948 ж/ Тимирязевтің бұл пікірін жете зерттеп, мақұлдады. Жоғарыда аталған пластидтер қаншама бір-бірінен өзгеше болса да, олардың барлығының да құрылысы, принципі бірдей - олар түссіз протоплазматикалық негізі стромадан және боялғыш заттардан тұрады.
Қазіргі кездегі электроскопиялық зерттеулер хлорофилдің құрылысы гранулды екендігін толық дәлелдеді. Табенцкийдің зерттеулері бойынша хлоропласт гранулалары стромаға қарағанда өте тығыз, төзімді екендігін және олардың, кейбір жағдайларда, стромада оңай бөлінетіндігін анықтады. Сонымен бірге Табенцкийдің зерттеулері хлорофилл дәнінің тұрақты статистикалық дене емес, белгілі бір заңдылыққа лайық өзгеріп отыратын динамикалық дене екендігін дәлелдеді. Мысалы: жас жапырақтың хлоропластары бірыңғай құрылысты, майда гранулды болып келсе, жетілген жапырақта ірі гранулды болып келеді, ал сарғайып келе жатқан ескі жапырақтарда хлоропластың стромасы суланып, кейде ол протоплазмаға жайылып кетеді де, ірі гранулдар протоплазмада бос жатады. Хлоропластың химиялық құрамын алып қарайтын болсақ, оның структуралық негізі белок екендігін көреміз. Хлоропластың денесі 30-45%-ға дейін белоктан /протоплазманың 80-95 %-ы белок/, 20-40% липоидты заттардан /пртоплазманың 2-3%-ы липоид/ тұрады.
Н. М. Сисакян, М. С. Черняк пластидтерде нуклеин қышқылдарын да тапқан. Хлоропласта өсімдік жапырағындағы барлық темірдің - 80%-ы, мырыштың- 70%-ы, мыстың - 50%-ы жинақталған. Бұл жағдайлар хлоропластың структуралық физика-химиялық құрылыстары да өте күрделі екендігін көрсетеді. Хлоропласт денесінің 75%-ы су болып келеді. Академик Н. М. Сисакянның зерттеулері бойынша хлоропласта күрделі ферменттер жинағы бар. Мұнда гидролитикалық та, тотықтыру, тотықсыздандыру ферменттері де бар. Сондықтан хлоропласта тек фотосинтез жүретін орын ғана деу қате болады. Бұлардың жалпы биологиялық рөлі өте үлкен.
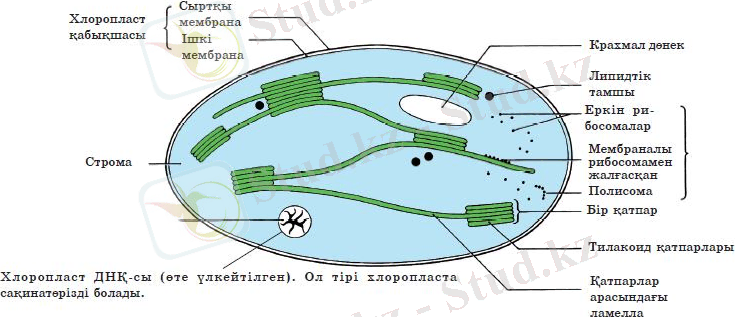
Хлоропластың екі мембранасы болады және сыртқы мембранасы - тегіс. Ол хлоропласты қоршаған цитоплазмадан шектейді.
Сыртқы мембранадан
хлорофилл электрондарының өтпеуі фотосинтездің жүзеге асуына мүмкіндік береді. Сонымен қатар кез келген сыртқы мембрана сияқты ол іріктемелі өткізгіштікті және хлоропласт пен «қоршаған ортасы» (цитоплазма) арасындағы зат алмасуды жүзеге асырады.
Ішкі мембрана
- жарықтың
хлорофилмен өзара әрекеттесуі
жүретін хлоропластың маңызды бөлігі. Осы жерде жарық энергиясы АТФ-тың химиялық энергиясына айналады. Яғни хлорофилл молекулалары мен басқа қосымша пигменттер хлоропластың ішкі мембранасына бекиді. Ішкі мембрана енді тегіс емес, ол ішіне томпиып шығып тұратын қатпарлар түзеді. Олар
тилакоидтар
деп аталады. Тилакоидтар мембранадан хлоропласт ішіне батып тұратын «қатпарлар» - грана тилакоидтарын немесе жай граналар түзеді (7-сурет) . Осы «қатпарлар» арасында жалғыз мембрана тартылысы немесе өткелдер -
строма тилакоидтары
немесе
ламеллалар
бар. Строма сұйықтығының ішінде хлоропластың ұзын өсін бойлай өте жұқа мембраналар бірінің үстіне бірі жиналған теңгелер сияқты орналасқан. Бұл мембраналар
ламелла
деп аталады.
Строма
- хлоропласт ішіндегі сұйықтық. Онда сақинатәрізді ДНҚ, РНҚ-ның алуан түрлері және рибосомалар бар. Олар хлоропластың тіршілік үдерістерін және фотосинтездің химиялық реакцияларын жүзеге асыратын барлық ферменттерді синтездейді. Мысалы, стромада органикалық заттар түзіледі - фотосинтез барысында стромада көмірсулар (глюкоза, қанттар, крахмал) синтезі жүреді. Сонымен қатар стромада бос оттек бөлінетін су ыдырайды. Барлығымыз тыныс алатын газ фотосинтез кезеңдерінің бірінде түзіледі. Осы реакциялар табысты жүзеге асуы үшін стромада барлық қажетті ферменттер синтезделеді. Хлоропластардың ішіне орналасқан стромалардың ламеллаларын құрайтын және тилакоидтардың құрамына кіретін мембраналарды байқауға болады. Строманың ламеллаларын құрайтын мембраналар жалпақ қапшықтар тәрізді: олар хлоропластардың ішкі мембранасымен тығыз байланысып, бір- біріне параллель орналасады. Жалпақ, жабық мембраналы қапшықтар - тилакоидтар бірінің үстіне бірі орналасып, граналар түзеді. Бір гранада 50-ге дейін тилакоидтар бар. Хлоропластың барлық граналары өзара мембраналар арқылы байланыскан. Хлорофилл осы граналарда шоғырланғандықтан, мұнда фотосинтез процесі жүреді.
7-сурет. Хлоропластың құрылысы. Өсiмдiктер ағзасын энергиямен қамтамасыз ететiн және глюкозаны синтездейтiн жартылай дербес органоид

Хлорофилл - көміртек, сутек, оттек және азот атомдары мен магний атомдарынан тұратын күрделі органикалық қосылыс. Ол фотосинтез процесіне тікелей қатысатын пигмент. Бұл топтың қазіргі кезде оншақты түрі белгілі. Хлорофиллдің түрлері: «а» хлорофилл - C35H72O5N4Mg көкшіл-жасыл пигмент, «б» хлорофилл - C55H70O6N4Mg сарғыш жасыл пигмент, «с» хлорофилл - C40H56 қызғылт сары пигмент каротин, «д» хлорофилл - C40H56O2 сары түсті пигмент ксантофилл. Фотосинтездік қабілеті бар организмдердің хлоропластарында міндетті түрде а-хлорофилл болады. Жоғары сатыдағы өсімдіктерде б-хлорофилл, қоңыр, диатом балдырларда қосымша с-хлорофилл, қызыл балдырларда д-хлорофилл кезеседі. Химиялық құрылымы жағынан барлық хлорофилдер пирролды магнийлі тұздары болып есептеледі. Олардың молекулалық құрамында 4 пиррол сакиналары өзара метилдік көпіршелері арқылы байланысып порфирин ядросын құрайды. Химиялық тегі жағынан қарағанда хлорофилдер дикарбонды, қосымша метил ж/е фитол спирті бар, хлорофиллин қышқылдарының күрделі эфирі. Жоғары сатыдағы өсімдіктер мен балдырларда болатын б-хлорофилдің а-хлорофилден айырмашылығы 2-пиррол сақинасындағы метил (СН3) тобы альдегид тобымен ауысқанында. Хлорофилл молекулаларында көміртегі мен азот атомдарының өзара үйлесімді, кезектескен жалпы ж/е қос байланыстардың нәтижесінде “ү мүшелі уақытша жүйе пайда болады. Қатты күйдегі хлорофилл көгілдір-қара түсті аморфты зат.
Хлоропластардың негізгі қызметі - күн сәулесін сіңіріп оны фотосинтез процесінде пайда болатын заттардың химиялық байланыстарындағы энергияға айналдыру . Фотосинтез (гр. фотос - жарық және синтез) - жоғары сатыдағы жасыл өсімдіктердің, балдырлардың, фотосинтездеуші хлорофилл және басқа дафотосинтездік пигменттер арқылы күн сәулесі энергиясын сіңіруі нәтижесінде қарапайым қосылыстардан (көмірқышқыл газы, су) өздерінің және басқа организмдердің тіршілігіне қажетті күрделі органикалық заттар түзуі. Фотосинтез нәтижесінде жер жүзіндегі өсімдіктер жыл сайын 100 млрд т-дан астам органикалық заттар түзеді (мұның жартысынан көбін теңіз, мұхит өсімдіктері түзеді) және бұлкезде олар 200 млрд-тай СО2 сіңіреді, оттегін бөледі.
Фотосинтезді алғаш зерттеушілер Швейцария ғалымдары Ж. Сенебье, Н. Соссюр және неміс химигі Ю. Майер болды. 19 ғасырдың 2-жартысында К. А. Тимирязев күн сәулесі энергиясы фотосинтез процесінде хлорофилл арқылы сіңірілетінін анықтады. 20 ғ-ң басында фотосинтездің физиологиясы мен экологиясына арналған маңызды зерттеулер жүргізіледі (В. В. Сапожников, С. П. Костычев, В. Н. Любименко, А. А. Ничипорович т. б. ) . 20 ғасырдың орта кезінен бастап фотосинтезді зерттеуде жаңа әдістер (газ анализі, радиоизотопты әдіс спектроскопмя. Электрондық микроскоп т. б. ) дамыды.
Жоғары сатыдағы жасыл өсімдіктер, балдырлар (көп клеткалы жасыл, қоңыр, қызыл, сондай-ақ бір клеткалы эвглена, динофлагеллят, диатом балдырлар) фотосинтезінде сутек доноры және шығарылатын оттек көзі су, ал сутек атомның негізгі акцепторы және көміртек көзі - көмірқышқыл газ. Фотосинтезге тек СО2 мен Н2О пайдаланылса углевод түзіледі. Фотосинтез процесіне өсімдік углевод түзумен қатар құрамында азоты және күкірті бар аминқышқылдарын, белок, молекуласы құрамында азот болатын хлорофилл де түзеді. Бұл жағдайда көмірқышқыл газбен қатар сутек атомының акцепторы және азот, күкірт көзі нитрат және сульфат болады. Фотосинтездеуші бактериялар молекула оттекті пайдаланбайды, оны бөліп шығармайды (бұлардың көбі анаэробтар) . Бұл бактериялар су орнына донор ретінде электрондарды не органикалық емес қосылыстарды (күкіртті сутек, тиосульфат, газ тәрізді сутекті) немесе органикалық заттарды (сүт қышқылы, изопропил спиртін) пайдаланады.
Фотосинтез аппаратының негізі - клетка ішіндегі органелла-хлоропластар (көк жапырақ клеткасында 20-100 болады) . Балдырлардың көпшілігінде фотосинтездік аппарат - клетка ішіндегі арнайы органелла-хроматофорлар, ал фотосинтездеуші бактериялар мен көк-жасыл балдырларда тилакоидтер. өсімдік фотосинтез процесінің негізі - тотығу-тотықсыздану. Мұнда квант энергиясы әсерінен 4 электрон мен протон су дәрежесінен (оның тотығуы) углевод дәрежесіне дейін көтеріледі. (СО2-ның тотықсыздануы) . Сөйтіп углеводтар фотосинтезі былай өтеді: СО2+Н2О С(Н2О) +О2+120 ккал/моль яғни СО2-ның бір молекуласының углевод дәрежесіне дейін тотықсыздануының бос энергиясы 120 ккал/моль болады. Демек, өсімдік фотосинтезі кезінде кем дегенде 3 квант («қызыл» кванттар энергиясы 40 ккал/моль) сіңірілуі қажет. әр түрлі жағдайда жасалған тәжірибе СО2-ның әр молекуласының тотықсыздануына 8-10 квант қажет екенін көрсетті. Көмірқышқыл газ да, су да, жарықты тікелей сіңірмейді, бұл қосылыстардың квантпен байланысқа түсуін хлоропласт не хроматофор структурасындағы хлорофилл а қамтамасыз етеді. Фотосинтездің биосферадағы маңызы да үлкен. Жер жүзіндеге, мысалы, көміртек, суттек, оттек, сондай-ақ N, S, P, Mg, Ca т. б. элементтер айналымы процесіне қатысы бар. Жер қалыптасқаннан бері фотосинтез нәтижесінде маңызды элементтер мен заттар бірнеше мың рет толық цикл айналымынан өткен. өсімдік өнімін арттырудың бір жолы - өсімдіктің фотосинтездік әрекетін үдету. Бұл үшін жапырақ көлемін үлкейту, жапырақ тіршілігін ұзарту, егістіктегі өсімдік жиілігін реттеу керек. СО2, ауа, су, топырақтағы қоректік элементтер жеткілікті болуы қажет. Фотосинтез аппаратының активтілігі жапырақтың анатомиялық құрылысына, фермент жүйесі активтілігіне, көміртек метабализмі типіне байланысты болады. өсімдік селекциясының, яғни СО2 ассимиляциясы тез жүретін өсімдік сорттарын шығарудыңда үлкен маңызы бар.
Афтотрофты және гетеротрофты жасушалар.
Пластикалық (анобализм) алмасу сипатына сәйкес табиғаттағы барлық жасушалар екі топқа бөлінеді. Хлорофилі бар өсімдік жасушаларының тірі табиғат үшін маңызы өте зор, себебі онда өзіне тән ерекше әрекеттер (процестер) жүріп жатады. Ол әрекеттер фотосинтез деген атпен ғылымға әйгілі. Фотосинтез дегеніміз күн сәулесі энергиясын химиялық байланыстар энергиясына айналдыратын күрделі механизмді әрекет.
Тынысалу және фотосинтез. Ағзалар тыныс алғанда қоректік заттар толық ыдырау үшін оттегі қажет екендігі баршамызға белгілі. Тынысалудың ең соңғы өнімі - көміртегі оксиді су жене бос энергия. Бұл соңғы өнімдер - фотосинтезге кажеттi негiзгi косылыстар болып табылады. Сондьктан, тынысалу фотосинтез кезiндегi энергияны жоққа шығарады. Алайда, тынысалу кезiнде жұмсалған пайдалы энергия фотосинтез кезiндегi алынған күн энергиясынан аз болатындығын төменгi тiзбектен көруге болады.
Энергияның ең көбi - күн сәулесiнiкi, коректiк заттар одан аз, ең азы кемiртегi оксидi, су жене оттегi. Фотосинтез кепсатылы күрделi әрекет. Мұнда күн сәулесi энергиясын химиялык байланыс энергиясына айналдыруда басты рөлдi хлоропластар атқарады. Пластиттердiң үш түрге бөлінетіндігi белгiлi, олар: лейкопластар, хромопласт жене хлоропласт. Бул үшеуiнiң де негiэi - строма деп аталатын ақуыз. Ал, фотосинтез әрекетi хлорофилл пигментi (жасыл түс беретiн) бар хлоропласт жасушасында жүредi. Ол үшiн хлоропласт жасушасының құрылысымен танысайык.
Биологиядағы барлық органоидтар сияқты, хлоропластың құрамы оның қызметiне сай күрделi болады. Хлорофилдер көк және қызыл түсті сәулелерді жұтып, жасылды шағылыстырады. Ол сәуле хлоропласт жасушасын жасыл етіп көрсетеді.
Фотосинтез пигменттері.
Жарық өсімдік ағзасына әсер етуі және соның ішінде фотосинтез үдерісінде қолданылуы үшін оны фоторецептор-пигменттер сіңіруі керек. Пигменттер - толқын ұзындығы белгілі бір жарықты сіңіретін боялған заттар. Жарық спектрінің сіңірілмеген бөліктері шағылысады да, пигменттің боялуына себеп болады. Сонымен, хлорофилл жасыл пигменті қызыл және көк сәулелерді сіңіреді, ал жасыл сәуле негізінен шағылысады. Пигменттердің құрамы ағзалар тобының жүйеленімдегі (систематикалық) орнына тәуелді. Фотосинтездеуші бактериялар мен балдырлардың пигменттік құрамы өте алуан түрлі (хлорофилдер, бактериохлорофилдер, бактериородопсин, каротиноидтер, фикобилиндер) .
Олардың жиынтығы мен арақатынасы әртүрлі топ үшін ерекше және көбінесе ағзалардың тіршілік ортасына тәуелді.
Хлоропластарда хлорофилдерден басқа сары, коңыр, кызғылт сары түстi каротиноидтар болады. Жоғары сатыдағы өсімдіктерде фотосинтез пигменттері едәуір аздау. Пластидтерде шоғырланған пигменттерді
хлорофилдер, каротиноидтер, фикобилиндер
деп үш топқа бөлуге болады.
Фикобилиндер
- цианобактериялар мен кейбір балдырларда кездесетін қызыл және көк түстің ерекше пигменттері.
Каротиноидтер
(латынша carota - сәбіз) - сары, күрең сары, қызыл түстердің пигменті. Негізінен бактериялар, саңырауқұлақтар және жоғарғы сатылы өсімдіктермен синтезделеді. Ол пигменттер ұзындығы баска толкындағы сәулелердi шағылыстырып, өз энергиясын хлорофильдерге берiп, фотосинтездiң жүрiсiн тездетедi. Каротиноидтар жасыл хлорофилдермен бүркенiп, көрiнбейдi, бiрақ күзде, тхлорофилдер бұзылганнан кейiн, оның жарқыраған түсi көрiнедi. Сондықтан да күзде жапырақтардың түсi сары жене кызғылт көрiнiс бередi. Кейбiр фотосинтезге қатысатын молекулалар мен пигменттер хлоропластағы фотосинтетикалық кабықшаны құрастыруға қатысады. Фотосинтетикалық қабықшалардың строма немесе хлоропластың негiзгi заты қоршайды. Строманың өзi хлоропласт жене жасушаның цитоплазмасын бөлетiн қабықшадан тұрады.
Ағзалар тыныс алғанда қоректік заттар толық ыдырау үшін оттегі қажет екендігі баршамызға белгілі. Тыныс алудың ең соңғы өнімі - көміртегі оксиді су жене бос энергия. Бұл соңғы өнімдер - фотосинтезге қажеттi негiзгi қосылыстар болып табылады. Сондықтан, тыныс алу фотосинтез кезiндегi энергияны жоққа шығарады. Алайда, тыныс алу кезiнде жұмсалған пайдалы энергия фотосинтез кезiндегi алынған күн энергиясынан аз болатындығын төменгi тiзбектен көруге болады.
Энергияның ең көбi - күн сәулесiнiкi, қоректiк заттар одан аз, ең азы көмiртек оксидi, су жене оттегi. Фотосинтез көпсатылы күрделi әрекет. Мұнда күн сәулесi энергиясын химиялық байланыс энергиясына айналдыруда басты рөлдi хлоропластар атқарады.
Фотосинтез әрекетi кезінде, АДФ-тiң ағзаларда атқаратын рөлi зор. АДФ - ағзалар деп отырғанымыз АТФ синтезiне Н - қоймасындағы энергияны пайдаланатын ферменттер. Аденозинтрифосфат АТФ. Жасушаның қимылдауына, ондагы жаңа ақуыз молекулаларының синтезделуi мен тасымалдануына, артық заттардың шығарылуына, яғни зат айналысының үздiксiз жүрiп тұруына осы АТФ-тiң энергиясы жұмсалады. Күн энергиясының АТФ түрiнде сакталған химиялық энергияга айналуы фотосинтездегi қоректiк заттардың калыптасуындағы маңызды кезең. АТФ тiрi ағзалардың өмiр сүруiндегi энергияның орталығы болады.
Фотосинтез кезiнде өсiмдiктер күн энергиясын органикалық заттардың молекулаларында сақтайды, ал тыныс алғанда қоректiк заттардың молекуласы ыдырап, ондағы энергия босап шығады. Яғни осы құбылыс АТФ-тiң синтезiне энергия екелетiнi жоғарыда көрсетiлген. АТФ молекуласьиның құрамында жоғары энергетикалы екi фосфат тобы болады. Бұл екi байланыс үзiлгенде басқа кез келген коваленттi байланыспен салыстырғанда көп энергия болiнедi. АТФ молекуласындағы фосфат тобының шеткi бiр молекуласы үзiлгенде 40 кДж энергия болiнедi, бұл энергияны жасуша пайдаланады. Осы кезде АДФ (аденозиндифосфат және босаған бейорганикалық фосфат қыскаша Фн деп жазылады) пайда болады. Қайтадан АТФ пайда болу үшiн АДФ пен фосфат тобы қосылу керек. Оған көп энергия жұмсалады, ол энергия фосфат тобының ыдырауынан және тыныс алудан алынады. Сонымен АТФ-тiң пайда болуьиның бiр жолы - ол АДФ-нiн басқа молекулалардан фосфат қосып алуы аркылы жүредi екен Гликолиз әрекетi кезiнде көптеген АТФ молекуласы түзiледi, мұнымен катар АТФ-тiң негiзгi бөлiгi химио-осмос барысы кезiнде пайда болады. АТФ молекуласының синтезделуінің осы жолын алпысыншы жылдары химио-осмос әрекетi деп атаган.
Химио-осмос хлоропластарда фотосинтез ксзiнде және митохондрияларда жасуша тыныс алғанда жүредi. Ол екi кезеңнен тұрады.
1. Энергияның жиналуы.
2. Жиналған энергияны АТФ синтезiне пайдалану.
Химио-осмос кезінде пайдаланылатын энергия - ол электрлiк заряды бар бөлшектер - иондардың қатысуына байланысты болатын электрхимиялык энергия қарсы зарядталған бөлшектер бiрiн-бiрi тартады. Егер осы белшектердiң қосылуына кедергi жасалса, электрхимияльиқ энергия жиналады:
Иондардың арасындағы кедергілерді ашса, электрохимиялық энергия жұмыс істейді.
Химио-осмос жоғарыда көрсетілген сызбанұсқаның негізінде жүреді. Хлоропластар мен митохондрияларда кедергінің рөлін органоидтар ішіндегі жарғақшалар атқарады.
Сіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz