Eriosoma lanigerum-ға қарсы Aphelinus mali арқылы биологиялық бақылауда фенологиялық модельдерге негізделген шешімдерді қолдау жүйесі және онжылдық дала мониторингі нәтижелері

ӨӘК: 634. 1. 055: 632. 937. 33
АЛМАНЫҢ ҚАН ҚЫЗЫЛ БІТЕСІНЕ (ERIOSOMA LANIGERUM) ҚАРСЫ ПАРАЗИТ APHELINUS MALI АРҚЫЛЫ БИОЛОГИЯЛЫҚ КҮРЕСУДЕ КЕШЕНДІ БАҚЫЛАУҒА НЕГІЗДЕЛГЕН ШЕШІМДЕРДІ ҚОЛДАУ ЖҮЙЕСІ
Каштасп А.
Қазақ ұлттық аграрлық зерттеу университеті
Андатпа
Алманың қан қызыл бітесі Eriosoma lanigerum (Homoptera: Aphidiae) дүние жүзіндегі алма бақтарында маңызды зиянкес болып табылады. Белгілі бір кең спектрлі инсектицидтерді алып тастағаннан немесе пайдалану шектелгеннен бері E. lanigerum Батыс Еуропадағы алма өсіретін аймақтардағы ең ауыр зиянкестердің біріне айналды. Қазіргі уақытта қан қызыл бітесі популяциясын тиімді шектеу химиялық күресу әдістері мен пайдалы буынаяқтылардың, әсіресе оның негізгі ерекше табиғи жауы, паразитоидты Aphelinus mali (Hymenoptera: Aphelinidae) арқылы биологиялық басу арасындағы жақсы синергияға негізделген. Білімге негізделген шешімдерді қолдау жүйесін әзірлеу үшін екі түрдің де егжей-тегжейлі мониторинг деректері далада (Алматы облысы) он жыл бойы (2010-2020) жиналды. Aphelinus mali ұшулары далада гүлдену алдында басталып, ең аз дегенде екінші ұрпақтың ұшуы аяқталғанға дейін бақыланды. E. lanigerum өсірудің маңызды кезеңдерінің маусымдық пайда болуы, мысалы, жүн түзіп басталуы немесе ауа бөліктерінде көктемде әрекеті және жорғалағыштардың колониялардан гүл шоғырларына немесе өркендерге көшуі мұқият бақыланды. Барлық алынған деректер тарихи және әдеби деректермен салыстырылды және популяция динамикасының фенологиялық үлгісінде талданды. Біздің нəтижелеріміз əзірленген модель арқылы бірінші буындағы ересек A. mali (алғашқы паразиттік белсенділік үшін маңызды жəне маусымның жалғасында А. mali ұрпақтарын жалғастыру үшін негіз) пайда болуын нақты болжауға болатындығын көрсетті. Демек, бұл ақпаратты далалық сынақтың нәтижелері көрсеткендей, дәл осы сәтте зиянды жанама әсерлері бар инсектицидтердің бүркуін болдырмау үшін пайдалануға болады. Сонымен қатар, E. lanigerum жорғалағыштарының гүл шоғырларына немесе өскіндеріне қарай қоныс аударуының басталуы модель арқылы дәл болжанады. Қорытындылай келе, біздің нәтижелеріміз модельді E. lanigerum-ды оның негізгі табиғи жауының максималды биологиялық басуы арқылы тиімді бақылауға қол жеткізу үшін бақылау процедураларының оңтайлы мерзімдері үшін шешімдерді қолдау жүйесі ретінде пайдалануға болатындығын көрсетеді.
Кілт сөздер : Қан қызыл бітесі- Eriosoma lanigerum; паразитоидты Apheninus mali; популяция динамикасы; шешімдерді қолдау
Кіріспе
Eriosoma lanigerum (Hausmann) - алма бақтарының ең зиянды зиянкестерінің бірі [1, 2, 3, 4, 5] . Ол қабығымен, бұтақтарымен, өркендерімен және тамырларымен қоректеніп, алма ағашын әлсіретеді, нәтижесінде ісіктер жиі пайда болады. Алма ағаштарының әлсіреуінен, қан қызыл бітесімен зақымданулар кішірек жемістердің пайда болуына әкеледі. Бітелер колонияларды қорғайтын қабықпен қамтамасыз ететін және оларға ерекше жүнді көрініс беретін ұзын ақ-мақта балауыз жіптерін шығарады. Ағаштың ауа бөліктерін нимфалардың («жорғалаушылар») көші-қонымен колонизациялаудан басқа, бітелер жыл бойы жетілген ағаштардың тамырларында кездеседі, олар жиі байқалмайды [6] . Екі ерекшелік (жүннен қорғайтын қабат және топырақ деңгейінен төмен жасырын тіршілік формалары) оларды бақылауды ерекше қиындатады. Батыс Еуропада коммерциялық алма бақтарында тек apterous virginoparae маңызды [7] және олар жылына 12 ұрпаққа дейін жетеді [8] . Қыстайтын иесі американдық қарағаш (Ulmus americana L. ) бельгиялық анар жемістер өндіретін аймақтарда болмағандықтан, E. lanigerum алма ағаштарында өзінің толық өмірлік циклін аяқтайды. Демек, сирек қанатты E. lanigerum морфтары мен жұмыртқалары ғана байқалады (2018 жылғы 28 қыркүйекте, 2017 жылғы 22 қыркүйекте және 2014 жылғы 7 қазандағы pcfruit бойынша бақылаулар, Керком, Синт-Труиден) . Ұрпағы тек американдық қарағашта өсетін жыныстық формалар беретін қанатты жүнді алма тлилерінің ықтимал дисперсиялық рөлі біздің аймақта анық емес. Қыстау негізінен бірінші жас нимфалар түрінде болғанымен [7], барлық жастықтар, сондай-ақ ересектер қыста Синт-Труиденнің айналасындағы бақыланатын бақтарда pcfruit арқылы байқалады.
1961 жылдан бастап мониторинг деректері Синт-Труиден аймағы үшін pcfruit-те жиналды. Бастапқыда бақылау бітедегідегі балауыз жіптердің өсуіне шоғырландырылды (осылайша, жалаңаш (балауыз жабыны жоқ) біте қайтадан белсенді болып, қоректеніп, түлеп, жүн деп аталатынды шығара бастағанда) . Біріктірілген зиянкестермен күресу (IPM) тактикасы кеңінен қабылданғаннан бері, кең спектрлі инсектицидтерден өсімдікті қорғаудың неғұрлым селективті құралдарына көшумен, қан қызыл бітемен күресудің негізгі нысанасы болды. Демек, соңғы онжылдықтарда мониторинг қызметінің бағыты осы өмірлік кезеңге ауыстырылды.
Сонымен қатар, қан қызыл бітемен қоректенетін жыртқыштардың тізімі, мысалы, сирфидтер, кокцинеллидтер, хризопидтер, карабидтер, өрмекшілер мен құлақшындар [2, 10, 11, 12, 13, 14, 15], ең маңызды табиғи жауларының бірі. E. lanigerum дүние жүзінде паразитоидты Aphelinus mali (Haldeman 1851) (Hymenoptera: Aphelinidae) [16, 17, 18] . Бұл иесіне тән (жалғыз) эндопаразитоид бітенің барлық партеногенетикалық кезеңдерін паразиттендіреді, үшінші сатыдағы нимфалар мен егде жастағы иелеріне артықшылық береді, бірақ оның иесімен салыстырғанда (жылына 4-7 ұрпақ) ұрпақтың аз болуы (жылына 4-7 ұрпақ) кемшілігі бар. [3, 19] . Оның үстіне, A. mali иесінен айырмашылығы, әсіресе ересек өмірлік кезеңде инсектицидтерге өте сезімтал [3, 5, 8, 20, 21] . A. mali бітенің өлі шыңдалған (мумияланған) денесінің ішінде толық жетілген дернәсіл немесе қуыршақ ретінде қыстайды, көктемде жаңа ересек паразитоидты аралар шығады [3, 7, 17] .
Асанте 10, 13, 15, 20, 25, 30 және 32 °C тұрақты температурада E. lanigerum жүнді алма тлилерінің apterous virginoparae өмірінің барлық кезеңдерінің даму уақытын хабарлады [22] . 10-25 °C диапазонында өмірдің барлық кезеңдері үшін даму жылдамдығы мен температура арасындағы жақсы сызықтық модель байқалды. Жалпы даму үшін төменгі және жоғарғы температура шегі сәйкесінше 5, 2 және 32 °C деп бағаланды. Бірінші, екінші, үшінші және төртінші кезеңдерді және жалпы дамуды аяқтау үшін қажетті орташа дәреже-күндік жинақтаулар сәйкесінше: 125, 6, 51, 0, 47, 7, 50, 7 және 267, 6 болды [22] .
A. mali дамуы да температураға кері негізделеді. Асанте және Дантанараяна (1992) 13-тен 30 °C-қа дейінгі диапазондағы даму жылдамдығы мен температура арасындағы жақсы сызықтық модельді байқады. Олардың нәтижелеріне сүйене отырып, өмірдің барлық кезеңдері үшін жалпы төменгі шекті мән ретінде 8, 3 °C пайдаланатын жалғыз сызықтық регрессия моделін пайдалану осы паразитоидтың маусымдық дамуын болжау үшін жеткілікті дәлдікке ие. Дамудың осы шартты шегі 8, 3 °C болғанда, екі жынысты біріктіріп, жұмыртқа салудан ересек пайда болғанға дейін толық даму үшін төменгі шекті мәннен жоғары 254, 8 градус-күн (DD) қажет болды [17] .
Температура шектері және DD температура қосындылары, 267, 6 DD кейін аптерозды virginoparae біте үшін жалпы даму уақыты 5, 2 °C ең төменгі шекпен [22] салыстырғанда ең төменгі шекті 8, 3 °C және 254, 8 DD А үшін жұмыртқадан ересекке дейін. mali оның паразитоидты A. mali-ге қарсы E. lanigerum үшін бастапқы немесе үздіксіз жетекшілігін растайды. A. mali максималды биологиялық басуы бар бітеде максималды бақылау деңгейіне жету үшін бақылау өңдеулерінің оңтайлы уақыты мен орналасуына шешім қабылдау қажет. Бұл зерттеудің мақсаты A. mali және E. lanigerum үшін әдебиеттер мен бақылау деректері негізінде фенологиялық болжау модельдерін құру және алма өсіру тәжірибесінде кешенді өсімдік қорғауда шешім қабылдау үшін олардың жарамдылығын тексеру болды. Нақтырақ айтсақ, біз A. mali-дің бірінші және екінші ұрпақтарының ұшуын болжау дәлдігін, сондай-ақ бірінші E. lanigerum жорғалағыштарының миграциясының басталуын анықтауды мақсат еттік. Өйткені, бұл бітеге қарсы дақылдарды қорғау спрейлерін қолданудың шешуші сәттері, олардың негізгі табиғи жауына (A. mali) теріс әсер етуден аулақ болу керек.
2. Зерттеу нысаны жəне əдістері2. 1. Бақылау
Келесі 11 жыл ішінде Aphelinus mali ұшуы сәуірден (жасылдан қызғылт бүршікке дейінгі фенологиялық кезең) ең аз шілдеге дейін, кем дегенде апта сайын (3-7 күн аралығы), Bug- типті 3 сары жабысқақ тұзақтағы ересектердің санын анықтау арқылы бағаланды. Scan® Yellow (Biobest, Westerlo, Бельгия) бақыланатын баққа 25 × 10 см² [23] . Алма ағашының шатырына шамамен 1, 5 м биіктікте қақпандар қойылды. 2010 жылдан бастап мониторинг Бельгияның шығысында орналасқан pcfruit зерттеу орталығының айналасындағы максималды 10 км радиуста немесе шегінде (50°46′22, 05″ N, 5) IPM алма бақтарында (Jonagold мутанттары, Гринстар немесе Браберн сорттары) мониторинг жүргізілді. Синт-Трюден төңірегінде бір радиуста алма бақтарында (бақылау жылына байланысты 2-5) қыстың аяғында және (ерте) көктемде E. lanigerum фенологиясы, әсіресе қыстауда жүн түзілуінің басталуы бақыланды. Бақыланатын әрбір бақшада осы зерттеудің әрбір мониторинг жылында жоғарыда аталған өсімдік құрылымдарының кем дегенде 10-ы (діңдегі ескі ағаштар, гүл шоғырлары, өркендер) мұқият тексерілді.
2. 2. Модельдеу
Төрт кезеңді аяқтау және жалпы даму үшін қажетті E. lanigerum даму уақыттары, сондай-ақ әртүрлі тұрақты температураларда A. mali жұмыртқасынан ересекке дейін даму уақыты 10 °C (тек E. lanigerum), 13 әдебиеттерде бар. , 15, 18, 20, 25 және 30 °C [17, 22] . Екі түр үшін де қаныққан көпмүшелік регрессия негізінде температураға негізделген фенологиялық модель құрастырылды. Әзірлеу жылдамдығы (=1/дамыту уақыты (күн) ) R studio бағдарламалық құралын пайдалану арқылы температураның (x = T (°C), f(x) = 1/әзірлеу уақыты (күн) = әзірлеу жылдамдығы) функциясы ретінде орнатылды. (R 3. 6. 2 нұсқасы) және «devRate» R бумасы [24] . Осы мақсатта «devRate» R бумасының функциялары мен деректер жиыны/тізімдері 2 полином («поли2») және көпмүшелік 4 («поли4») функциялары 5 және 6 көпмүшелік функцияларының формулаларымен кеңейтілді. S1) . «devRate» R бумасы параметрді бағалау үшін сызықты емес ең кіші квадраттарды бағалау процедурасын пайдаланады. Үлгілердің сәйкестігі диагностикалық қалдық графиктері және Akaike ақпараттық критерийі (AIC) арқылы бағаланды (Қосымша S1 кестесін қараңыз) . Тұрақты температураның функциясы ретінде даму жылдамдығының графиктері S1 және S2 қосымша суреттерінде көрсетілген. Қоршаған ортаның температурасына қатысты даму уақыты мен ұрпақтары «devRate» R-пакетінің «devRateIBM» функциясы арқылы бағаланды, бұл ретте A. mali аналықтары пайда болғаннан кейін бірден жұптасып, жұмыртқа салады (бұрынғы бақылауларға негізделген [25] ), ал apterous virginoparae қанды алма бітелері ересек болғаннан кейін бірден жаңа нимфаларды береді (бұрынғы бақылауларға негізделген [22] ) . Бірінші және екінші буындағы (G1 және G2) A. mali ересектерінің болжамды алғашқы күндері және 2010-2020 жылдар аралығындағы тәуліктік температура деректеріне негізделген жаңа бірінші virginoparae E. lanigerum «тексергіштердің» болжамды алғашқы күндері салыстырылды. осы кезеңдегі әрбір жыл үшін далалық бақылаулар деректері. E. lanigerum және A. mali үшін тиісінше минимум 5, 2 °C және 8, 3 °C төмен шекті температуралар үшін полиномдық функциялар теориялық тұрғыдан алғанда, даму қарқыны нөлге (сонымен қатар жоғары температураларда (32, 7 °C жоғары) ) орнатылды. теріс (бірақ физикалық мүмкін емес) теріс даму деңгейі) .
Болжамдардың дәлдігін сандық бағалау үшін болжау үлгілерінің өнімділігі болжанған (x) күндердегі байқалған (y) сызықтық регрессияларын пайдалана отырып, бақыланатын және болжамды күндер (31/03 кейінгі күндер саны) арасындағы келісімді тексеру арқылы бағаланды [ 26] 11 жылға (2010-2020), егжей-тегжейлі мониторинг және ауа райы деректерімен. Идеал модель r = 1 корреляция коэффициентімен және y = x регрессия сызығының ең кіші квадраттарымен дәлелденетіндей дәл және дәл болады [27] . Барлық статистикалық талдаулар Rstudio бағдарламалық құралымен орындалды (R 3. 6. 2 нұсқасы) .
2. 3. Алма бағында далалық зерттеу
Далалық сынақ 2020 жылы алма бағында E. lanigerum және A. mali біртекті болуымен жүргізілді. Сынақ өңделмеген бақылауды және Decis EW (құрамында 100 г/л дельтаметрин бар) өңдеуді қоса алғанда, 4 қайталаумен, толығымен рандомизацияланған блок дизайнында орнатылды. Бұл емдеу осы мақалада көрсетілмеген, бірақ (статистикалық) бірге талданған басқа емдеу әдістерімен үлкен сынақтың бөлігі болды. Әрбір сынақ алаңы 7 ағаштан тұрды (3, 10 м өңделген ағаш биіктігі, өсімдіктердің арақашықтығы 1, 75 м қатарда), олар моторлы рюкзактарға арналған бүріккіш (Stihl түрі, SR 430 үлгісі) арқылы бүркілген. ) бұтақтардың астыңғы жағындағы бітеге (және A. mali ересектеріне) жету үшін шығыс түтіктегі 45° дефлектор экраны бар. Decis EW 2020 жылдың 16 сәуірінде 333, 3 л/га LWA су көлемін пайдаланып, 0, 300 л/га жапырақ қабырғасының ауданы (LWA) (=4, 5 г дельтаметрин/га LWA) доза жылдамдығымен шашыранды. Алма ағаштары тік дақыл болғандықтан, өнімнің дозасының жылдамдығы мен су көлемі EPPO ұсынғандай (PP1/239(3) [30] нұсқауы) га LWA үшін көрсетілген. LWA ағаштар саны × қатардағы отырғызу қашықтығы (d) × өңделген ағаш биіктігі × 2 жағы бойынша есептеледі және сынамалы бақтағы 15 500 м2/га топырақ бетіне сәйкес келеді.
Әрбір учаске үшін жұқтырған өскіндер (2 жастағы + жаңа өсу) үшін бұрын белгіленген 10 аумақтағы жалпы біте колониясының жалпы беті (см2) тормен (0, 5-1 см2 шаршылармен) бағаланды, ол колониялар. Сонымен қатар, сол жерлерде байқалған ересек A. mali саны және кейіннен паразиттенген (мумияланған) біте саны (ашық, сондай-ақ жабық) бірдей жерлерде бағаланды (егер тығыз жүн болса, алдымен балауыз жойылды. абайлап үрлеу арқылы, біте/мумияларды жай көзбен санау үшін) . Паразитизм дәрежесі 10 см2 E. lanigerum колония бетіндегі паразиттенген біте санымен есептелді.
Сынақ нәтижелері Unistat Statistical Package, 10 нұсқасы (Unistat Ltd., Лондон, Ұлыбритания) арқылы статистикалық талдаудан өтті. Bartlett's Chi-Square және Bartlett-Box F-тестімен дисперсиялардың біртектілігін растағаннан кейін, әртүрлі шығыс нұсқалары (Anova, Таблица, қалдықтар, бірнеше салыстырулар) бар GLM (Жалпы сызықтық модель) процедурасы орындалды.
3. Зерттеу нəтижелері жəне оларды талқылау3. 1. A. mali және E. lanigerum үшін полиномдық регрессия үлгілері
Келесі көпмүшелік функция A. mali үшін температураның (°C) функциясы ретінде жұмыртқадан ересекке дейінгі даму жылдамдығын көрсетеді:
f(x) = −2, 940 × 10−3 − 1, 716 × 10−4x + 4, 895 × 10−4x2 − 5, 389 × 10−5x3 + 2, 653 × 10−6x4 − 4, 303 × 10−8x5 (1)
Төмендегі көпмүшелік функция apterous virginoparae E. lanigerum біте даму жылдамдығын температураның (°C) функциясы ретінде көрсетеді:
f(x) = 1, 013 × 10−3 − 9, 014 × 10−4x − 2, 011 × 10−4x2 + 1, 044 × 10−4x3 − 8, 425 × 10−6x4 + 2, 731 × 10−7x5 × 9) (10−7x5 −9)
Есептелген көпмүшелік регрессия үлгілері мен жеміс бақшасының температурасының нақты деректеріне сүйене отырып, екі түр үшін де үздіксіз даму есептелді және жинақталды (шекті температурадан төмен немесе одан жоғары температуралар үшін жинақталған даму қосылмаған) A. mali негізінен толық дернәсіл және A. mali жұмыртқасы ретінде қыстайтындықтан, дернәсіл мен қуыршақ даму уақыты бір-бірімен 1:4:5 [7, 17, 31] байланысты болғандықтан, бұл қыстайтын формалардың барлығы да қыстайтын деп жорамалданады. жылдың басында жұмыртқадан ересекке дейін дамудың жартысы (01/01) . Сонымен, көктемде бірінші жаңадан шыққан A. mali ересектері (= маусымда алғашқы A. mali ұшуларының басталуы) даму уақытының қалған жартысы аяқталған кезде болжанады, бұл жалпы жұмыртқаның 50% -ына сәйкес келеді. Екінші буындағы A. mali бірінші жұлдыздарының пайда болуы келесі толық ұрпақ уақытына жеткенде болжанады. E. lanigerum негізінен бірінші жастағы нимфа ретінде қыстайтындықтан [7], барлық біте жылдың басында (01/01) бірінші нимфалық кезеңде болады деп болжанған. Жаңа бірінші инстарлы virginoparae («жыртқыштар») ата-аналық колониялардан таралып, ағаштың шатырында жаңа колониялар түзетін алғашқы қоныс аударатын формалар ретінде қарастырылатындықтан [31], миграцияның басталуы E. lanigerum бір толық ұрпақты аяқтаған кезде болжанады.
3. 2. A. mali бірінші және екінші ұрпақ ересектердің пайда болуының болжамының дәлдігі
Өлшенген сағаттық температура деректеріне негізделген болжамды даму 2010 жылдан 2020 жылға дейін А. мали ересектерінің далалық бақылау деректерімен салыстырылды. 1-кестеде жаңа бірінші ұрпақ (G1) A. mali ересектері (=) үшін болжамды және бақыланатын күндер маусымда A. mali ұшуларының басталуы) және екінші буын (G2) A. mali ересектері көрсетіледі.
1-кесте. Жаңа бірінші буынның (G1) A. mali ересектерінің (маусымда A. mali ұшуларының басталуы) және екінші ұрпақтың (G2) A. mali ересектерінің болжамды және бақыланатын күндері.
1 Сары жабысқақ бақылау тақталары тексерілген соңғы күн арасындағы орташа күн, ол үшін тексерілген пластиналардың ешқайсысы А. мали ересектерінің ұсталғанын көрсетпеген және бақылау тақталарында жаңадан шыққан A. mali ересек(тері) анықталған алғашқы күн.
2 G1 рейстері аяқталғаннан кейін сары жабысқақ бақылау тақталары тексерілген және тексерілген тақталардың ешқайсысында жаңадан ұсталған А. мали ересектері көрсетілмеген соңғы күн мен жаңадан пайда болған G2 A. mali ересектерінің бірінші күні арасындағы орташа күн жабысқақ пластиналарда қайтадан анықталды.
3 A. mali G1 және G2 арасын ажырата алатын егжей-тегжейлі мониторинг деректері жоқ. Болжамдардың дәлдігі болжамды (x) күндерде байқалған (y) сызықтық регрессиялары арқылы бағаланды. Нәтижелер 1-суретте көрсетілген.
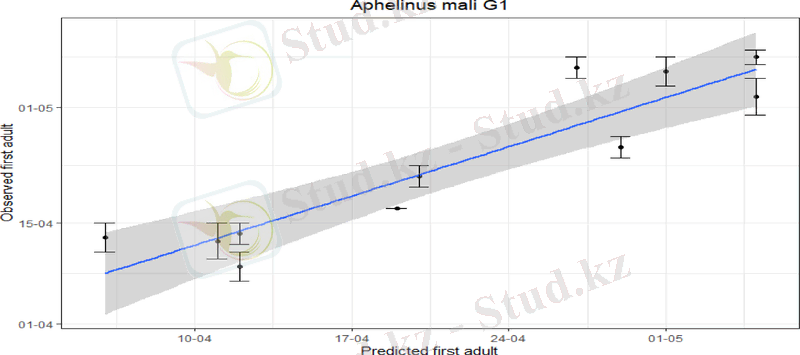
1-сурет. Алма бақтарында бақылау арқылы болжанған бірінші буындағы (G1) A. mali ересектері мен бірінші байқалған G1 A. mali ересектерінің сызықтық регрессиялық талдауы. Қате жолақтары сары жабысқақ бақылау тақталары тексерілген соңғы күнді білдіреді, ол үшін тексерілген пластиналардың ешқайсысы А. мали ересектерінің ұсталғанын көрсетпеді және бақылау тақталарында жаңадан пайда болған A. mali ересек(тері) анықталған бірінші күн.

2-сурет. Алма бақтарында бақылау арқылы болжанған екінші ұрпақтың (G2) A. mali ересектері мен байқалған екінші G2 A. mali ересектерінің сызықтық регрессиялық талдауы. Қате жолақтары G1 рейстері аяқталғаннан кейін сары жабысқақ бақылау тақталары тексерілген соңғы күнді және тексерілген тақталардың ешқайсысында жаңадан ұсталған А. мали ересектерін көрсетпеген және жаңадан пайда болған G2 A. mali ересектері қайтадан анықталған бірінші күнді білдіреді.
3. 3. E. lanigerum көші-қон болжамдарының дәлдігі
Сонымен қатар, біте үшін өлшенген сағаттық температура деректеріне негізделген болжамды даму далалық бақылау деректерімен салыстырылды. Атап айтқанда, жорғалағыштардың (маусымдағы бірінші E. lanigerum ұрпағының жаңа 1-жұлдыздары) гүл шоғырларына немесе жаңа өскіндерге көшу сәті далалық бақылау арқылы мұқият тіркелді. Алғашқы көші-қон әрекетінің болжамды және бақыланатын күндеріне шолу 2-кестеде көрсетілген.
... жалғасыСіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz