Ажырық, арпа және беде өсімдіктерінің тұз және құрғақшылық стрессіне физиологиялық-биомассалық жауаптары мен альдегид оксидазаның рөлі

3 Зерттеу нәтижелері және талдау
3. 1 Тұзды стресс жағдайында өсімдіктердің биомассасын анықтау
Өсімдіктің тұзды топырақта өсуі және ерте балғын кезеңінде стреске бейімделу механизмінің дамуы өсімдік түрінің қалыптасуында маңызды рөл атқарады. Өсімдіктің тұзды стреске төзімділігі оның тіршілік циклінің әрбір кезеңінде ауысып, өзгеріп отырады. Тұздану өсімдік тұқымының өнуіне кедергіні су тапшылығын тудыру, осмотикалық эффект әсері немесе иондық үрдістерге метаболикалық шығын келтіру арқылы тежейді. Галофитті өсімдіктер жоғары тұз мөлшеріне төзімді, және тұздың өсімдік ұлпасына енуінәртүрлі механизмдер арқылы төмендетіп, өсімдіктің булануынан сақтайды. Біз өз зерттеулерімізде сор және сортаң жерлерде мал азығы ретінде өсетін галофит-ажырықты қарастырдық. Қоршаған ортаның қолайсыз жағдайларында өскедіктен ажырық өсімдігінің жапырағы өте майда, өсімдік дәнінің аз болуы және жақсы дамыған тамыр жүйесімен ерекшеленеді. Біз тәжірибелерімізд үш түрлі тұз концентрациясында 0, 100 және 200Mm NaCl ажырық, арпа және беде өсімдігінің өсуі, биомассасын зерттедік. 28 күндік өсімдіктердің жер асты және жер үсті мүшелерін тұзды стресс жағдайында бақыладық. Зерттеу нәтижесі бойынша ажырық өсімдігінің биомассасы тұз мөлшері артқан сайын, кеміді.

Ажырық өсімдігінің жапырақ биомассасы тұз мөлшері артқан сайын кеми бастады. 200Mm NaCl тұз мөлшерінде жапырақ биомассасы бақылауға қарағанда 20%-ға төмендеді. Ал тамыр биомассасының оптимальды мөлшері бақылаумен салыстырғанда 100Мм NaCl-да 54%-ға артса, 200Мм NaCl-да 18%-ға жоғарылады.
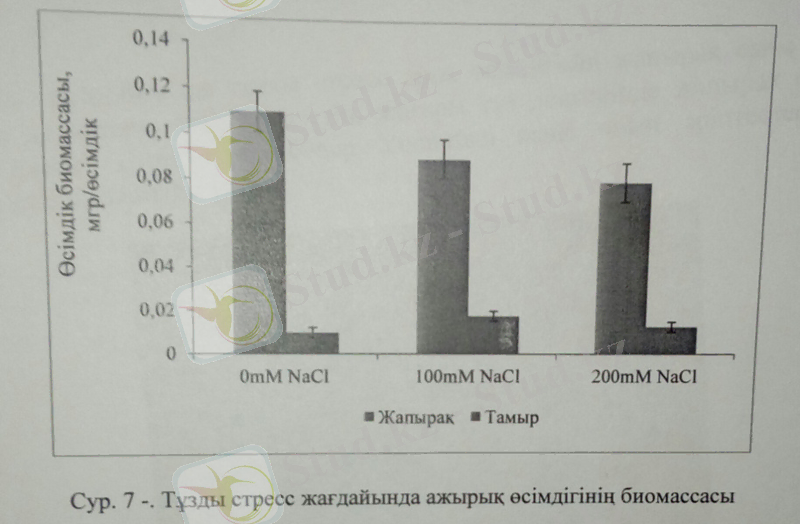
Әдетте тұзды стресс жағдайында тұздың уытты әсері өсімдіктің тамыр мүшесін айтарлықтай зақымдап, галофитті өсімдіктерде тұз концентрациясы артса да, оның уытты әсері төмендеп, K+/Na+ қатынасы жоғары мөлшерде болады. K+/Na+ қатынасының жоғары деңгейде артуы өсімдіктердің тұзды стреске төзімділіктің жоғары болуының айқын белгісі. Ажырық өсімдігі топырақтағы тұздың мөлшері 1%-ға дейінгі мөлшерде төзімді(Li and Liu 1994) және геномында хромосома саны өте аз (2n=14) . Өсімдіктер абиотикалық және биотикалық стресс жағдайында әртүрлі әсер етеді, оның ішінде тұзды стресті де атап өтуге де болады. Сондықтан астық тұқымдас өкідерінің, соның бірі-ажырық өсімдіктерінің тұзға төзімділік деңгейін анықтау, селекционерлерге тұзға төзімді өсімдік культураларын дамытуда аса маңызды мәлімет болып табылады.
Тұзды топырақта өскен галофит түрлері биомассасының жоғары мөлшері төменгі тұз концентрациясында байқалады. Бұл нәтижелер Kelly тағы басқалары, Gorham және Harrouni әріптестерінің еңбектерінде баяндалады. Олардың еңбектерінде тұздың төмен концентрациясы кейбір галофит түрлерінің өсуін арттырады, бірақ, жоғары тұзды стресс өсу мен биомасса өнімділігін төмендетеді деген ортақ тұжырымға келді. Ал Mass-тың зерттеулеріне сүйенсек көптеген галофиттердің өсуі тұз мөлшері артқанда төмендейтінін байқап, әрбір галофиттердің өздеріне тән төзімділік шегі бар екенін мәлімдейді.
Келесі тәжірибелерімізде біз тұзды стрестің арпа өсімдігіне әсері зерттелді. Тұзды стресс арпа өсімдігінің фенологиялық аспектілеріне, жапырақ санына, жапырақ алақанына, тамырдың өну қарқынына, тамыр/жерүсті мүшелерінің қатынасына, және жалпы өсімдіктің құрғақ массасына теріс әсер етеді.
Біздің тәжірибемізде тұзды стресс арпа өсімдігінің жапырақ саны мен мөлшеріне де айтарлықтай әсер етіп, жоғары тұз деңгейінде жапырақ саны азайды. Осыған құқсас тәжірибелер Yumurtaci және оның әріптестерінің еңбегінде қайталанады.
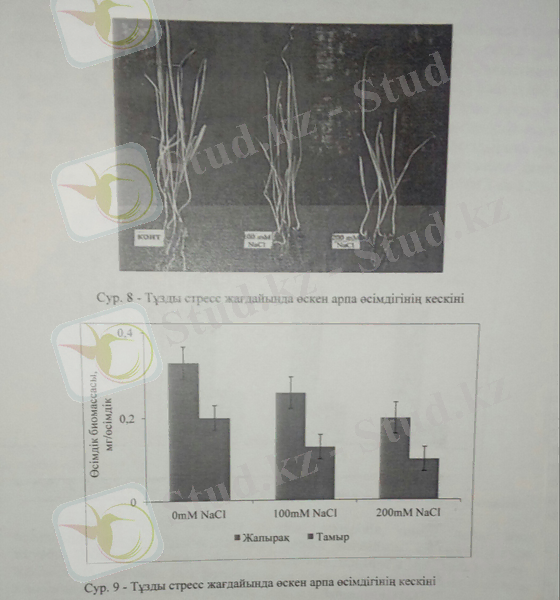
Арпа өсімдігінің дамуына тұзды стресс айтарлықтай әсер етті. Жапырақ биомассасы тұз мөлшері артқан сайын төмендеді. Орташа тұз мөлшерінде жапырақ биомассасы 22%-ға кемісе, жоғары тқз деңгейінде 40%-ға төмендеді. Ал тамыр биомассасы бақылаумен салыстырғанда орташа тұз жағдайында 35%-ға кемісе, 200Мм NaCl-да 50%-ға төмендеді. Тұзды стресс барысында арпа өсімдігінің жер асты және жер үсті мүшелерінің төмендеуі NaCl мөлшерінің токсинді әсері және иондық баланстың бұзылуымен байланысты. Тұздың жоғары мөлшерінде өсімдік өсуінің төмендеуі топырақтан суды сіңіру қабілетінің төмендеуімен және осмос қысымының артуымен де байланысты.
Арпа өсімдігінің биомассасымен бірге тзды стресс жағдайында біз өркен және тамырдың ұзындығында зерттедік. Арпа өсімдігінің сабақтарының ұзындығы жоғары тұз деңгейінде 200Мм NaCl-да бақылаумен салыстырғанда 15%-ға кемісе, 100Мм NaCl сабақ ұзындығына айтарлықтай кері әсер еткен жоқ. Өсімдіктердің өсіп-дамуы біз зерттеген өсімдіктерде тұзданудың әртүрлі деңгейінде бақылаумен салыстырғанда кеміді. Ал тамыр ұзындығына келер болсақ, тұздану деңгейі артқан сайын кеміп, өркен ұзындығына ұқсас тенденция көрсетті. Бақылаумен салыстырғанда орташа тұздылық деңгейі тамыр ұзындығы 28%-ға кемісе, жоғары тұз деңгейінде 200Мм NaCl-да 52%-ға төмендеді.
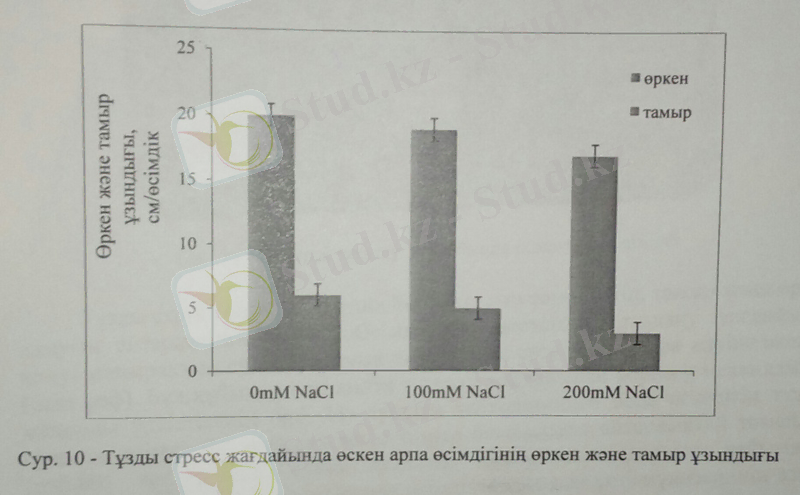
Тұздануға қарсы көптеген өсімдік түрлерінің, соның ішінде арпа өсімдігінде кемуі олардың қолайсыз стресс жағдайына қайтарған жалпы жауабы болып табылады. Арпа өсімдігінің жапырағы да тұзды стресс жағдайында сарғайады, бұл фотосинтез үрдісінің төмендеп, өткізгіш шоқтарда жиналған NaCl иондарының токсинді әсерімен байланысты.
Беде(Trifolium repens L. ) -тұзды стреске аса сезімтал, бұршақ тұқымдасына жататын көп жылдық кейде бір жылдық шөптесін өсімдіктер. Республиканың барлық жерінде, шалғындықтарда, ойпаң жерлерде, өзен, жол бойларында, тау бөктерлерінде өседі. Оның биікт. 15-50 см-дей, сабағы жұмыр, тамыры жуан болады. Жапырағы күрделі, 5-9 жапырақшалары бар. Гүлі ұсақ ұызыл, қызғылт, сары не ақ түсті, шар тәрізді домалақ немесе сопақша келген шашақ гүлшоғырға топталған. Жемісі-бұршақ, оның 1-2, кейде 3-6 дәні бар. Бедені таза күйінде немесе еркек шөппен, бидайықпен, тарғақшөппен қосып себеді. Бұл жағдайда ол мол өнім береді әрі мал сүйсініп жейді. Беденің құрамында протеин, каротин, витамин, азотсыз заттар, клетчатка, май, күл және фосфор көп болады, жылына 3-4 рет орылады. Беденің топырақтың сортаңданбауына, эрозияға қарса күресте тигізетін әсері зор, сонымен қатар гүлдеген кезінде одан жақсы бал жиналады.
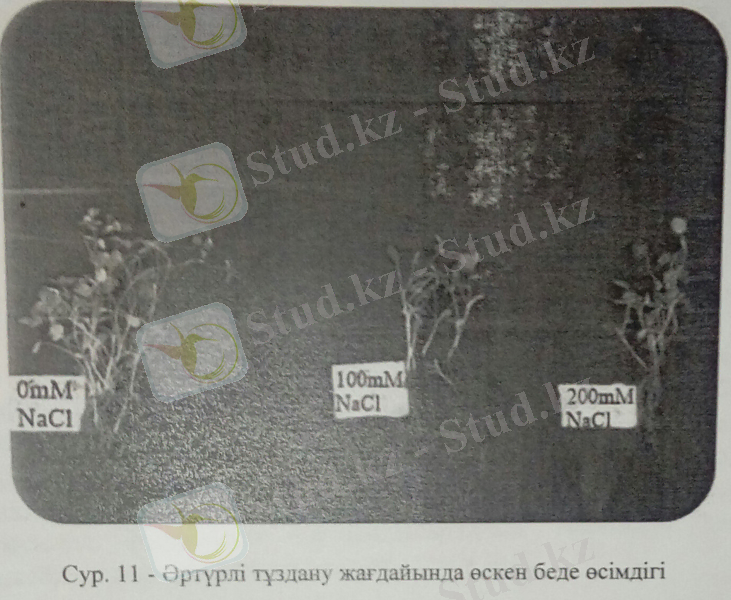
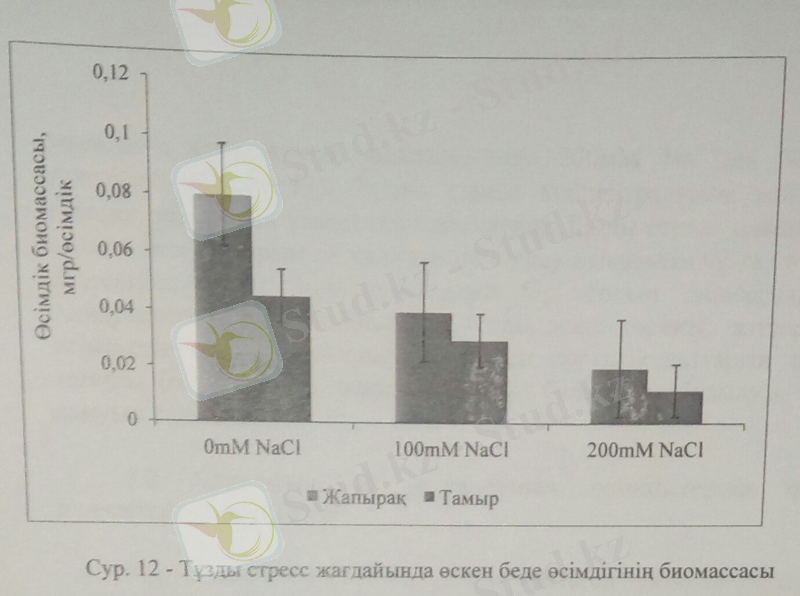
Тұзды стресс беде өсімдігінің биомассасын төмендетіп, тамыр және өркен дамуын айтарлықтай кемітті. Өсімдік биомассасының тұздану жағдайында кемуі жапрыақтың суды булануын төмендетіп, ерте ескіруі мен өлуіне әкеледі. Бұл құбылыс өсімдіктің тұздану жағдайынан қорғану механизмінің маңызды үрдістерінің бірі. 11-суретте келтірілгендей беде өсімдігін тұзбен суарсақ, өсімдіктің жапырақ және тамыр биомассасы айтарлықтай төмендеді. Кейбір ғалымдардың пайымдауы бойынша тұзды стресс жағдайында өсімдіктердің жер үсті мүшелірінің азаюы осмостық стрестің қарқынды жзеге асуымен байланысты.
Жапырақ биомассасы бақылаумен саластырғанда жоғары тұз мөлшерінде 200Мм NaCl-да 37, 5%-ға төмендесе, тамыр биомассасы 30%-ға кеміді. Өсімдік биомассасының тұздану жағдайында кемуі фотосинтез үрдісінің төмендеуімен байланысты. Фотосинтез қарқындылығының төмендеуі хлорофилл синтезінің тұздану барысында шектелуімен түсіндіруге болады.
Стресс жағдайында хлорофилл мөлшерінің төмендеуі оның мембранамен байланысуымен ал хлорофилл мөлшерінің тұрақтылығы мембаранының стабилдігімен байланысты. Тұзды стресс барысында бұл тұрақтылық жиі өзгеріп отырады. Хлорофилдің төмендеуі тұхзға сезімтал өсімдіктерде тұзға төзімді өсімдіктермен салыстырғанда жиі кездеседі. Бұл нәтижелер тұзды стресс барысында фотосинтез қарқындылығының төмендеуі хлорофилл мөлшерін тежейді деген тұжырымды растай түседі.
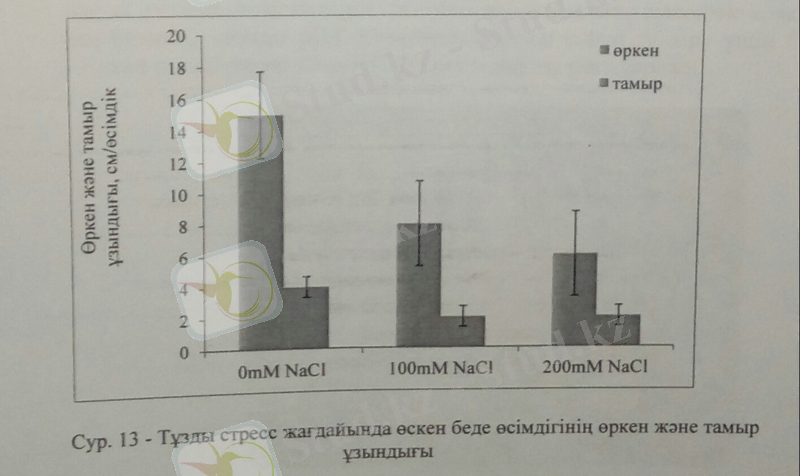
Тәжірибе соңында 28 күннен соң тұзды стресс барысында өсімдіктің өркені мен тамырының ұзындығы төмендеді. Беде өсімдігінің өркені мен тамыр ұзындығы бақылаумен салыстырғанда 200Мм NaCl-да 30 және 50%-ға төмендеді. Тұзды стресс концентрациясы жоғарылаған сайын, өсімдік сабағының ұзындығы төмендейді. Тұзды стресс жағдайында өсімдіктің сабағы мен тамырының ұзындығын өлшеу өсімдіктің тұзға төзімділік деңгейін анықтауда басты индикаторлардың бірі болып табылады, себебі, тамыр топырақпен бірінші байланысып, суды және қоректік заттарын сіңіріп, қалыпты дамиды. Тұздану жағдайында бұл баланстың бұзылуы, өсімдіктің өсіп-дамуын тежейді.
3. 2 Құрғақшылық жағдайында өсімдіктердің физиологиялық параметрлерін анықтау
Құрғақшылық стресі өсімдіктің көптеген физиологиялық аспектілеріне кері әсерін тигізеді, соның ішінде өсімдіктің фотосинтез қабілетін төмендетеді. Ал егер стресс ұзақ уақытқа созылса, онда өсімдік өсуі және биомассасы айтарлықтай төмендеді. Өсімдіктерде қоршаған ортаның қолайсыз әсерлеріне бейімделу үшін көптеген физиологиялық және биохимиялық механизмдері қарқында түрде жұмыс жасайды. Ұзаққа созылған құрғақшылық стресі өсімдіктің көптеген маңызды қызметтерін тежейді, соның ішінде көмірсулардың балмасуы бұзылады. Ал ол фотосинтез процесін бұзып, өсімдіктің өсу-дамуын және өнімділігіне кері әсер етеді. Құрғақшылыққа қарса жауап ретінде өсімдіктерде лептесіктер жабылып, жапырақ алақанында СО2 диффузиясы артады. Құрғақшылық жағдайын тудыру үшін біз жоғарыда аталған өсімдіктерді 10 күнде бір рет және екі рет суардық.

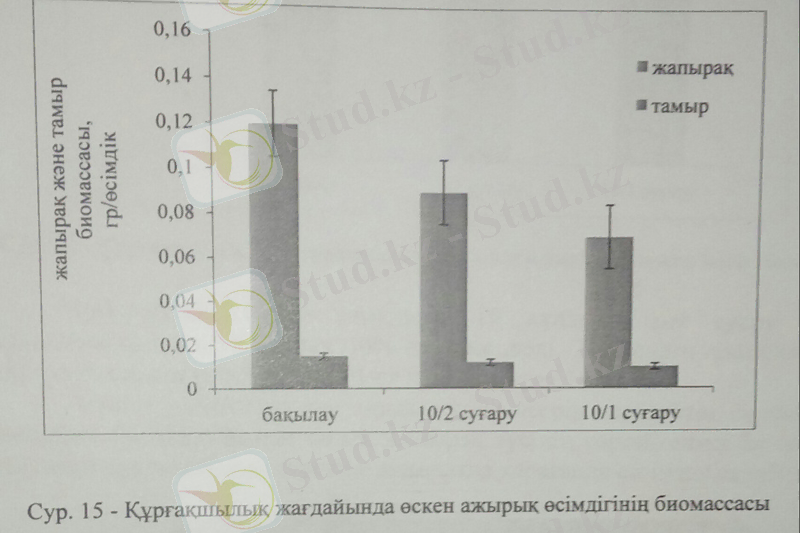
Біздің зерттеулеріміздің негізгі мақсаты-ажырық, арпа және беде өсімдіктерінің құрғақшылық стресіне жауабын зерттеу болып табылады. Стресс жағдайында өсімдіктің жалпы биомассасын анықтау өсімдіктің қаншалықты төзімді екенін анықтайтын негізгі индикатор болып табылады. Khan et al. (2007) және оның әріптестерінің тұжырымдауы бойынша өсімдіктің жоғары биомассасы құрғақшылықта анықталды. Ал біздің зерттеулерімізде, үш өсімдікте де жоғары биомасса бақылау өсімдіктерінде анықталды. Әр өсімдіктің құрғақшылыққа берер жауабы әртүрлі су деңгейін ұлпасында сақтау қабілетімен байланысты. Мысалы, ажырық өсімдігінің жапырақ биомассасы бақылаумен салыстырғанда 10 күнде 1рет суару өсімдік өсуіне айтарлықтай теріс әсер етпесе де, 10 күнде 2 рет суару барысында 40%-ға төмендеді. Тамырға келер болсақ, 10 күнде 2 рет сурау барысында тамыр биомассасы бақылаумен салыстырғанда 33%-ға төмендеді.
Ал өсімдіктің жер үсті және жер асты мүшелерінің ұзындығы да құрғақшылық стресі қлғайған сайын төмендей бастады.
Құрғақшылық жағдайында өсімдіктің жасушасының созылуы және өсуі төмен тургор қысымына байланысты тежеледі. Жасуша тургоры әдетте осмостық қысымының реттелуімен байланысты түсіндіріледі (Shao et al 2008) . Өсімдіктің биомассасы стресс жағдайында олардың төзімділігін анықтайтын факторлардың бірі. Khan және оның әріптестерінің тұжырымы бойынша құрғақшылық жағдайындағы жоғары биомасса өсімдік төзімділігінің жоғары екенін білдіреді. Ажырық өсімдігінің 10 күнде екі рет суғарылуы өз кезегінде оның биомассасына айтарлықтай әсер етпеді. Арпа өсімдігімен салыстырғанда оның құрғақшылық стересіне төзімділігі жоғарырақ екендігін көреміз. Құрғақшылық жағдайында өсімдік ұзындығының бәсеңдеуі жасуша созылуының шектелуімен және қартаю үрдісінің жеделдеуімен байланысты.
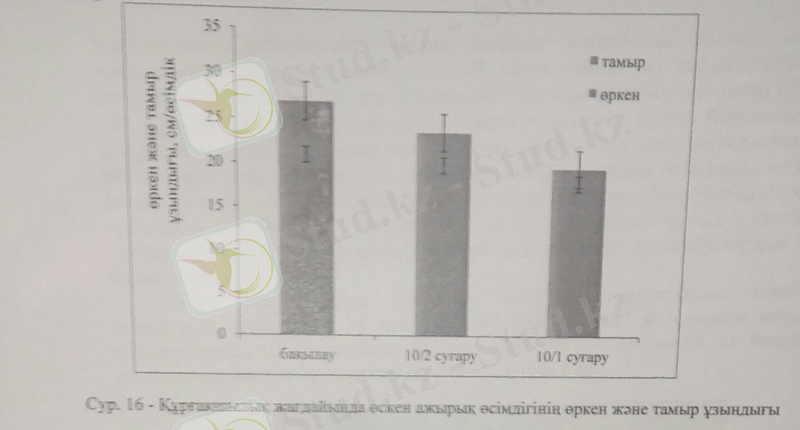
Ажырықтың өркен ұзындығы 10 күнде 2 рет суару барысында бақылаумен салыстырғанда 14%-ға төмендеді. Тамырдың ұзындығы жоғары құрғақшылық жағдайында 28%-ға кеміді.
Арпа-өсімдігіне құрғақшылықтың әсерін зерттейтін болсақ, олардың жапырақ биомассасынан бөлек жапырақ түсінің сарғайғанын байқаймыз. Бұл арпаның құрғақшылық стресіне ажырыққа қарағанда сезімтал екенін байқаймыз.

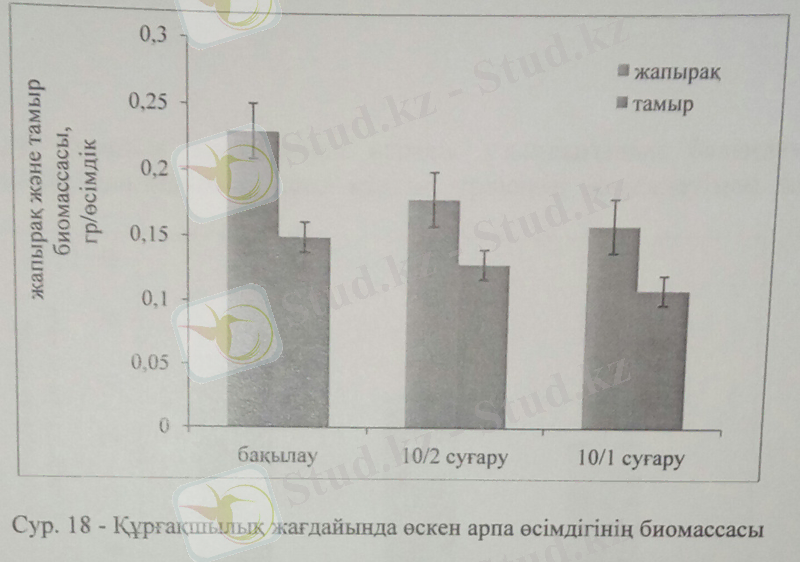
Құрғақшылық барысында фотосинтез процесі төмендеп, хлорофилл пигменттерінің синтезделуі азайды. Арпаны 10 күнде 1 рет суарғанда, жапырақ сарғайып, биомассасы төмендеді. Арпа өсімдігі құрғақшылық стресіне ұшырағанда жапырақ және тамыр биомассасы төмендеді. 10 күнде бір рет суарғанда жапырақ биомассасы бақылаумен салыстырғанда 45%-ға төмендетсе, екі рет суарғанда 25%-ға кеміді. Ал тамыр биомассасы 10 күнде екі рет суарғанда 38%-ға азайды.
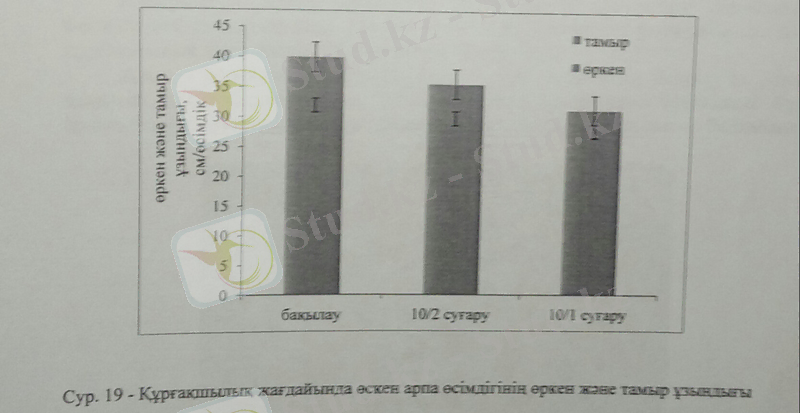
Құрғақшылық стресі өсімдіктің салмағына ғана емес, өркен және тамырдың дамуына да кері әсерін тигізді. Дегенмен, құрғақшылық стресі тамырға қарағанда өсімдіктің өркеніне теріс әсері айқын бйқалады. Өркен ұзындығының оптимальды мөлшері бақылау жағдайында 33см-ге жетсе, жоғары құрғақшылық деңгейінде ол 28см-ге азайды, ал тамыр ұзындығы бақылауда 12см болса, жоғары құрғақшылықта 8см-ге дейін төмендеді. Құрғақшылықтың орташа деңгейі арпа өсімдігінің жалпы биомассасына және өркен, тамыр ұзындықтарына айтарлықтай теріс әсер көрсетпесе де, жоғары құрғақшылықтың ұзақ уақыт орын алуы соңында өсімдіктің өліміне әкелді. Жалпы осы өсімдіктердің қоршаған ортаның қолайсыз жағдайларна төзімділігі мен бейімделу механизмін анықтау келешекте тұздану, құрғаққа төзімді өсімдік топтарын дамытуға бағытталған негізгі алғышарттардың бірі болып табылады.
Келесі тәжірибемізде біз беде өсімдігінің құрғақшылық стресіне төзімділігін анықтадық. 28 күн өсірілген беде өсімдігі жоғарыдағы өсімідік культураларындай 10 тәулікте бір рет және екі рет суарылды, ал бақылау жағдайында қажетті су мөлшері күнделікті суғарылды.
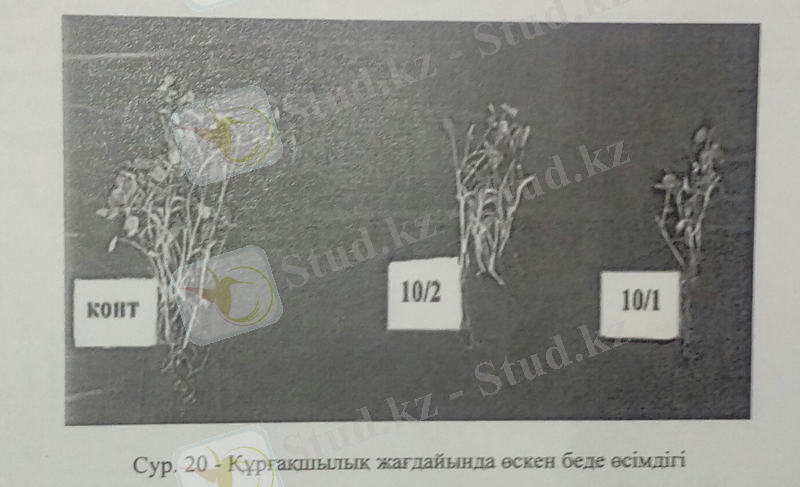
Беде өсімдігінің құрғақшылыққа сезімталдық деңгейі ажырық және арпа өсімдіктерімен салыстырғанда өте жоғары екендігін 20, 21-суреттен байқауға болады.
Беде жапырағының биомассасы оташа құрғақшылық жағдайында 20%-ға төмендесе, жоғары құрғақшылықта 47%-ға азайды. Ал тамыр биомассасы жоғары құрғақшылық деңгейінде бақылаумен салыстырғанда 38%-ға төмендегенін байқаймыз.
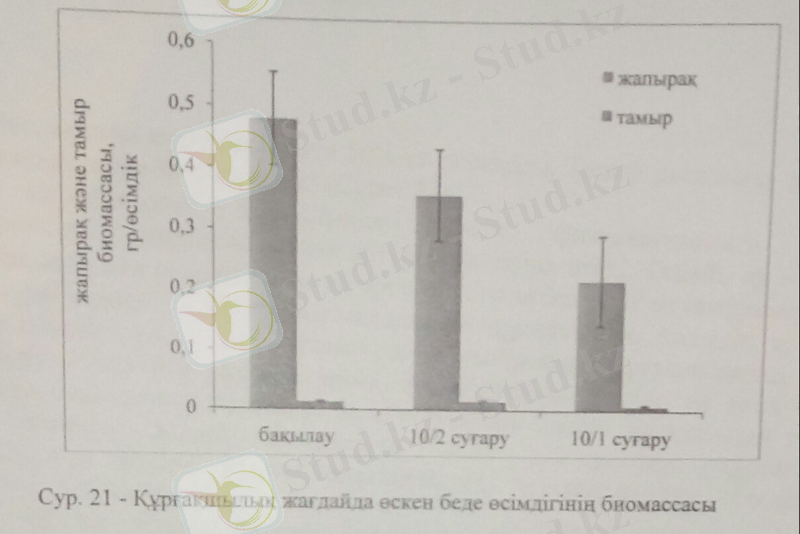
Тамыр биомассасының құрғақшылық жағдайында төмендеуі, өсімдіктің тіршілігі үшін маңызыды минералды тұздардың сінірілуінің тежеліп, азот пен фосфор дефицитімен байланысты. Нәтижесінде қажетті элементтердің қоры төменгі жапырақтардан алынып, алдымен олардың өліміне, кейін басқа мүшелердің өліміне әкеледі. Өсімдіктер тыныштық күйіне ауысады немесе тіршілігін толығымен тоқтатады.
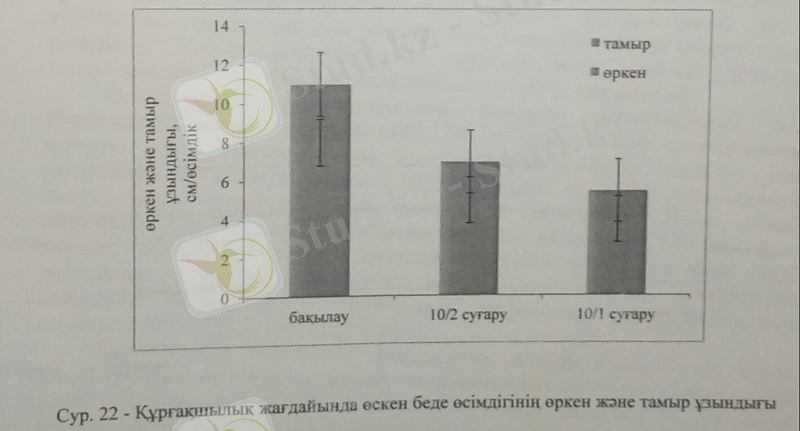
Беде өсімдігінің тамыр және өркен ұзындықтары құрғақшылық стресіне айтарлықтай жоғары сезімталдық көрсетті. Өркеннің оптимальды ұзындығы бақылау жағдайында жоғары болса, орташа құрғақшылық жағдайында 37%-ға кемісе, жоғары құрғақшылықта 50%-ға қысқарды. Тамыр ұзындығы да жоғары құрғақшылықта 50%-ға кеміді.
Құрғақшылық жағдайында өсімдік физиологиясында болатын ерекшеліктер әртүрлі өсімдікте әртүрлі жағдайда өтеді. Себебі, әр өсімдіктің құрғақшылыққа сезімталдығы да әртүрлі. Сол себепті біз өз тәжірибелерімізде үш түрлі өсімдіктің құрғақшылыққа әсерін қарастырдық. Ажырық, арпа және беде өсімдіктерінде байқағанымыздай салыстырмалы құрғақшылыққа жоғары төзімділік деңгейін ажырық өсімдігінде байқауға болады, ал арпа өсімдігі орташа сезімталдық көрсетсе, беде өсімдігінің дамуы айтарлықтай тежелді. Өсімдіктердің жалпы биомассасы және ұзындықтары құрғақшылық жағдайында кеміп, жоғары құрғақшылық деңгейінде үш өсімдікте де айтарлықтай азайды.
Өсімдік биомассасының құрғақшылық жағдайында азаюы өсімдік жапырағында тургор қысымының төмендеуі мен фотосинтез қарқындылығы кеміп, фитогормондар синтезі тежелді. Фитогормондардың кемуі жер үсті мүшелерінің өсуін тежеп, кейін тамыр мүшелерінің де дамуына кедергі келтіреді. Жасушалардың ұзарып өсуінің тоқтауы, өсімдік мүшелерінің баяу өсуіне әкеледі.
Соңғы он жылдық зерттеулер барысында көптеген ауыл шаруашылық дақылдарын дәстүрлі сұрыптау әдісі арқылы құрғақшылққа төзімді өсімдіктерді дамыту кең етек жая бастады. Осындай әдістер арқылы төзімді өсімдіктер тобын анықтау және құрғақшылққа төзімді бейімделу механизмдерін түсіну бұл жағдайлардың алдын-алуға көмектеседі.
Өсімдіктердегі су тапшылығы көптеген физиологиялық және биохимиялық өзгерістерді тудырады, соның ішінде осмос потенциалының өзгеруі, ион гомеостазының ауытқуы, осмолиттердің жиналуы және фотосинтез пигменттерінің азаюы. Сонымен қатар бос радикалдардың артуы, липид пероксидациясы, хлоропласт пигментінің азаюы өсімдік жасушасында тотығу стресін тудырады. Осындай жағдайларды құрғақшылыққа төзімді өсімдік топтарын анықтау, құрғақшылыұ тудыратын теріс жағдайлардың алдын-алуға көмектеседі. Сол себепті, өсімдіктердің бейімделушілік қабілеттері анатомиялық, морфологиялық, жасушалық және молекулалық деңгейде зерттелуі қажет.
Құрғақшылық барыснда өсімдіктердің жоғары балғын және құрғақ биомассасының сақталуы Khan және оның әріптестерінің зерттеуі бойынша аталған стреске төзімділігінің жоғары болуымен сипатталады. Ал біздің зрттеулеріміз бойынша үш өсімдікте де балғын биомасса құрғақшылықтың орташа және жоғары деңгейінде де төмендегенін көруге болады. Сонымен бірге өсімдіктердің өркен және тамыр ұзындықтары да құрғақшылықтың жоғары деңгейінде үш өсімдікте де айтарлықтай кеміді. Бұл өзгеріс өсімдіктердегі құрғақшылық жағдайындағы физиологиялық өзгерістермен тікелей байланысты. Атап айтар болсақ, фотосинтез қарқынының төмендеуі, өсімдік транспирациясының артуы, минералды элементтердің сіңірілуінің баяулауы т. б.
3. 3 Абиотикалық стресс жағдайында альдегид оксидаза ферментінің рөлі
Альдегид оксидаза(AO) цитоплазмада кездесетін, молекулалық салмағы 300кДа болатын гомодимерлі фермент. Альдегид оксидаза ферменті жоғары сатылы өсімдіктерде абсциз альдегидті абсциз қышқылына айналдыратын молибдофермент. Берілген өсімдіктердің тұзды, құрғақшылық және басқа да абиотикалық стрестерге төзімділігі олардың физиологиялық және генетикалық ерекшеліктеріне байланысты.
АО ферментінің активтілігі нативті-полиакриламидті гелде анықталды, субстрат ретінде ванилин және индол-3-сірке қышқылы қолданылады. АО-ферменті субстратты ауқымы кең, өсімдік фитогормоны, абсциз қышқылының соңғы сатысын катализдеуші фермент.
АО-ферментінің активтілігі арпа, ажырық және беде өсімдіктерінің тұзды стресс жағдайында, құрғақшылық жағдайында 28 күнде, жапырақ ұлпасынан бөліп алынған экстакттен анықталды. Құрғақшылық жағдайында бақылау, 1 күнде 1 рет және 2 рет суару барысында арпа, ажырық және беде өсімдіктерінде анықталды. 23 суретте көрсетілгендей арпа өсімдігінде оптимальды АО активтілігі жоғары құрғақшылық жағдайында анықталды. Бақылаумен салыстырғанда АО активтілігі арпада жоғары құрғақшылықта 2, 5 есеге артқанын байқаймыз. Ал ажырық өсімдігінде 10/1 және 10/2 құрғақшылық жағдайында АО активтілігі бақылаумен салыстырғанда 4 және 5 есе жоғарылығанын байқаймыз. Беде өсімдігін сипаттар болсақ, АО активтілігі құрғақшылық жағдайында бақылаумен салыстырғанда 10/1 және 10/2 суарғанда 4, 5 және 6 есеге дейін артты.
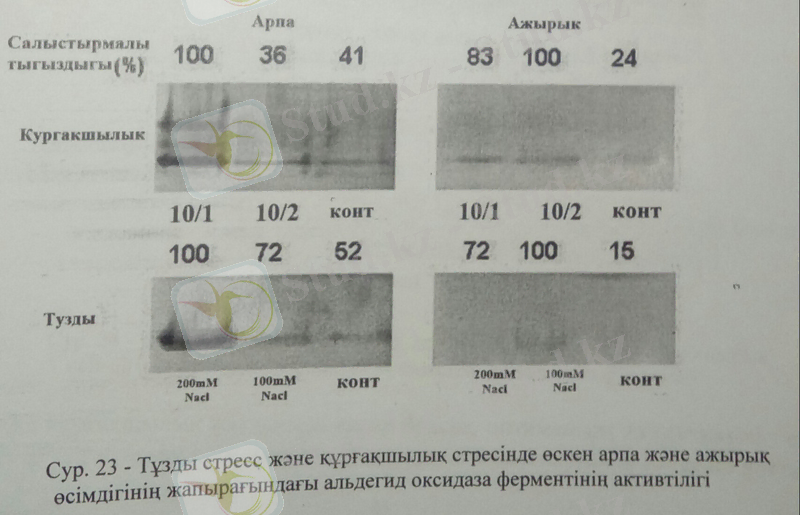
Ал тұзды стреске келер болсақ, біз өз тәжірибелерімізде үш түрлі тұз концентрациясын пайдаландық-0, 100 және 200Мм NaCl. АО- активтілігі арпа, ажырық және беде өсімдіктерінде зерттелді. 23 суретте көрсетілгендей тұзды стресс жағдайында арпа өсімдігінде АО оптимальды активтілігі жоғары тұз деңгейінде 200мМ NaCl-да ең жоғары АО-активтілігі табылды.
АО ферментінің активтілігін келесі тәжірибелерімізде арпа және ажырық өсімдіктерінің тамыр ұлпасынан тұздану және құрғақшылық жағдайында қарастырдық. Арпа өсімдігінде тұзды стресс жағдайында, яғни тұз мөлшері жоғарылыған сайын АО активтілігі артса, құрғақшылық жағдайында да бақылаумен салыстырғанда АО активтілігінің артқанын байқаймыз. Ал ажырық өсімдігіне келер болсақ, ажырық өсімдігінде АО активтілігі тұздану барысында АО активтілігі төмендесе, бақылауда 2, 5 есеге артты. Бұндай жоғарылауды аталған өсімдіктердің тұзды стреске айтарлықтай жоғары болуымен түсіндірсе болады, себебі, галофитті өсімдіктер үшін тұзсыз орта кей жағдайларда керісінше өсуді тежеп, олардың қалыпты дамуы үшін тұздың белгілі бір мөлшері қажеттілігімен байланысты болуы мүмкін. Тұзды стрестің әсері тамырдағы АО-ферментінің активтілігіне қарқынды әсер етті. Себебі, тамыр АБҚ-биомөлшеріне қатысады, ал жапырақ өте аз мөлшердегі АБҚ синтездейді.
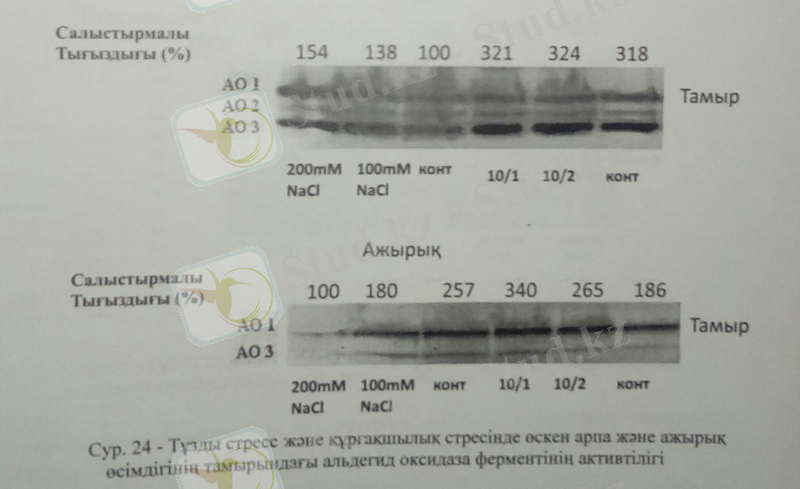
Ал құрғақшылық жағдайына келер болсақ, оптимальды АО активтілігі он күнде бір рет суарған өсімдіктерде бақылаумен салыстырғанда екі есеге артты. Яғни құрғақшылық стресі ажырық өсімдігі үшін айтарлықтай теріс әсер етіп, АО активтілігі бақылаумен салыстырғанда артты. Құрғақшылық жағдайында АО артуы құрғақшылық барысында жоғарылыған абсциз қышқылының артуымен байланысты болуы мүмкін.
Келесі тәжірибемізде біз беде өсімдігінде тұзды стресте АО ферменттерінің активтілігін арпа және ажырық өсімдіктерінде салыстырмалы түрде анықтадық. Өсімдік жапырағындағы АО ферментінің активтілігінің оптимальды мөлшері 200Мм NaCl-да анықталды.
Тұзды стрестің 100Мм NaCl деңгейінде беде өсімдігінің жапырағындағы АО-ферментінің активтілігі бақылаумен салыстырғанда 4, 6-есеге артса, жоғары тұздылықта 200Мм NaCl-да 5, 5 есеге артқандығын көруге болады. Осыған ұқсас нәтижелер яғни, АО-ферментінің активтілігі тұзды стресте артқан нәтижелер арпа, райграс, және қызанақ өсімдіктерінен де табылды. Стрестер тудырған гормондық тұрақтылықтың өзгеруі өсімдіктің стреске төзімділік деңгейіне тіклей байланысты. Абсциз қышқылының мөлшері қоршаған ортаның қолайсыз факторларында:төменгі температура, тұзды стресс, құрғақшылықта артады. Бұл нәтижелер тұзды стрессбарысында АБҚ-ның артуымен және АБҚ-ның өсімдіктің жер үсті мүшелеріне транспортының қарқынды жүруімен байланысты. Себебі, абсциз қышқылы өсімдіктің стресс барысында бейімделу механизміне қатысады және АО-ферментінің апобелогын кодтайтын ген экспрессиясының жоғарылауына және ақуыз молекуласының пост-трансляциялық активациясына байланысты.
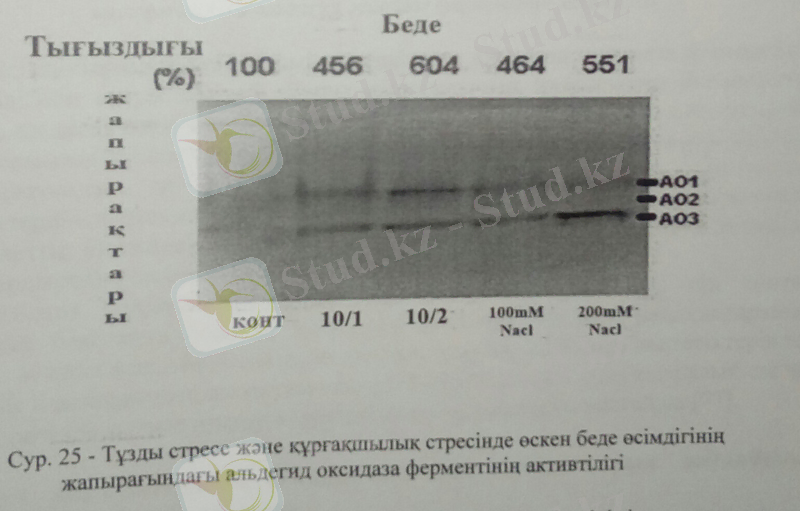
Келер тәжірибемізде біз АО активтілігін беде өсімдігінің тамырынан анықтадық. Беде өсімдігінің тамыр жүйесіндегі тұздану жағдайында АО ферментінің активтілігіне келер болсақ, оптимальды АО активтілігі 100Мм NaCl-да табылса, құрғақшылық жағдайында бақылау өсімдігінде анықталды.
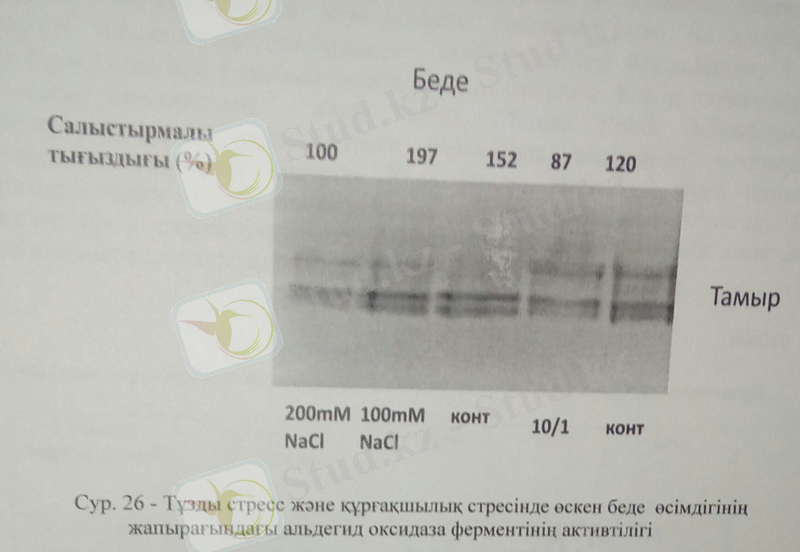
Өсімдіктердің құрғақшылық жағдайында альдегид оксидаза ферментінің активтілігінің артуы тікелей абсциз қышқылының жұмысымен байланысты. Себебі, альдегид оксидаза ферменті өсімдік фитогормоны абсциз қышқылының соңғы сатысын катализдейді. АО активтілігінің артуы абсциз қышқылының жоғарылауымен тікелей байланысты. Құрғақшылық барысында өсімдіктердің лептесіктері жабылып, абсциз қышқылының синтезі қарқынды түрде артты. Нәтижесінде өсімдік жапырағындағы АО ферментінің активтілігі де айтарлықтай жоғарылады.
... жалғасыСіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz