Бидай мен күріш өскіндерінің тамыр түктерінің морфометриялық талдауы: тұздың әсері

ҚАЗАҚСТАН РЕСПУБЛИКАСЫНЫҢ БІЛІМ ЖӘНЕ ҒЫЛЫМ
МИНИСТІРЛІГІ
ӘЛ-ФАРАБИ АТЫНДАҒЫ ҚАЗАҚ ҰЛТТЫҚ УНИВЕРСИТЕТІ
Биология факультеті
Генетика және молекулалық биология кафедрасы
Магистрлік диссертация
Бидай және күріш өскіндерінің тамыр түктерін морфометриялық талдау
Орындаған 1 курс магистранты Іркітбай А. "" 2011 ж.
Ғылыми жетекшісі, б. ғ. д Айташева З. Ғ. "" 2011ж.
Кафедра меңгерушісінің
рұқсатымен
қорғауға жіберілді, б. ғ. д Айташева З. Ғ. "" 2011ж.
Алматы 2011
РЕЗЮМЕ
Дипломная работа в общей сложности состоит из 43 листа, в том числе из 15 схем, 6 таблиц. Число используемых литератур 51, из них 2 Казахская литература, 17 русская литература, 36 на англисском языке.
Ключевые слова: корневые волоски, пшеница, рис, проростки, солоноватая среда, ядро.
Цель работы - Изучение морфометрических признаков корневых волосков из проростка пшеницы и риса
Обязанности:
- Дать характеристику на морфологию корневых волосков из проростка пшеницы и риса;
- Анализ на длину корневых волосков;
- Анализ на объем ядро корневых волосков.
МАЗМҰНЫ
АНЫҚТАМАЛАР
ҚЫСҚАРТЫЛҒАН СӨЗДЕР
КІРІСПЕ
НЕГІЗГІ БӨЛІМ
4
5
6
8
1 ӘДЕБИЕТТЕРГЕ ШОЛУ
8
1. 1 Тамыр түктерінің жалпы сипаттамасы
8
1. 2 Тамыр түктерінің атқаратын қызметі
13
1. 3 Өсімдік тамырының құрылысы
13
1. 4 Өсімдіктердің тамыр арқылы қоректенуі мен элементтерді сіңіруі
14
1. 5 Тамыр түктерінің өсу кезеңдері
16
1. 6 Тамыр түктері жөніндегі зерттеулер
18
1. 7 Тұздың өсімдіктерге және тамыр түктеріне әсері
21
1. 8 Бидай мен күріштің шаруашылықтық құндылығы
22
2 ЗЕРТТЕУ МАТЕРИАЛДАРЫ МЕН ӘДІСТЕРІ
26
3 НӘТИЖЕЛЕР МЕН ОЛАРДЫ ТАЛҚЫЛАУЛАР
3. 1Тамыр түктерінің морфологиялық сипаттамасы
3. 2Тамыр түктерінің морфометриялық белгілеріне
тұздың әсері
3. 2. 1Бидай өскінінің тамыр түктерінің морфометриялық
белгілеріне тұздың әсері
3. 2. 2Күріш өскінінің тамыр түктерінің морфометриялық
белгілеріне тұздың әсері
3. 3Тамыр түктерінің ядро құрылымын салыстырмалы талдау
29
29
30
31
33
37
ҚОРЫТЫНДЫ
39
ҚОЛДАНЫЛҒАН ӘДЕБИЕТТЕР ТІЗІМІ
40
Анықтамалар
Тамыр түктері- тамыр түктері тамыр бетінің эпидермис (ризодермис) клеткаларының кішкене, түткшелі, 1-2 мм ұзындықтағы созылған бөлігі.
Ядро- тұқымқуалаушылық мәліметтерін сақтайтын және тасымалдайтын органелла
Өскіндер- топырақ бетіне шыққан сабағы мен жапырағы бар жас өсімдік.
Қысқартылған сөздер
Каз-126. Казахстанская-126
Сар-29. Саратовская-29
Ген-1. Генотроф-1
Вел-2401. Велютинум-2401
Лют-19001. Лютесценс-19001
КІРІСПЕ
Жоғары сатыдағы өсімдіктердегі түктердің атқаратын қызметі алуан түрлі. Бірақ олардың өсімдіктер үшін маңызы әлі күнге дейін толық анықталмаған, тек өсімдіктерде маманданған түктердің қызметі ғана толықтай зерттелген. Мысалы, жәндікке улы өсімдіктер маманданған түктердің көмегімен насеком денесінен қоректік заттарды сорып алуға қабілетті болып келеді. Қазіргі кезде минералды қоректену элементтерін сіңіріп алуға қабілетті тамыр түктерінің қызметі де терең зерттелуде [1-5] .
Өсімдіктердің тамыр түктері тамыр бетінің эпидермис (ризодермис) клеткаларының кішкене, түткшелі, 1-2 мм ұзындықтағы созылған бөлігі. Тамыр түктерінің бірінші кезектегі қызметі тамырдың беткі ауданын ұлғайтып, өсімдікті P, Fe, Zn, Si және басқа да микроэлементтермен қоректенедіру болып табылады [6] . Тамыр түктерінің өзгерісін ағылшын ғалымы Caradus (1979) алғаш рет Clover деген өсімдікте зерттеген [7] . Томат өсімдігінің әдеттегі тамыр түктеріне қарағанда мутантты түктері ұзынырақ болатындығы және тамыр түктері Arabidopsis thilana өсімдігінде терең зерттелген [8] . Бидайың тамыр түктерін Gahoonia және Szareiko деген ғалымдар зерттеген. Тамыр түктері өсімдіктердің өсіуінде айтарлықтай қажет емес, мысалы, бидайдың, күріштің, жүгерінің тамыр түктері жоқ мутантты формаларын қоректік заттар дайын ортаға өсірсе, олар жақсы өсе алады [9-10] . Қазіргі күнде Arabidopsis thilana өсімдігінің тамыр түктерінде 40-жуық тіркескен гендер белгілі [11] .
Ылғалды ауада өскен тамырдың пішіні цилиндр немесе конус тәрізді болады. Топырақта қатты бөлшектер әсерінен түктер жабысады, қисық болады немесе ісініп кетеді. Сулы ортада өсетін өсімдіктерде негізінде тамыр түктері болмайды. Топырақтағы тамыр түктерінің саны ондағы ылғалдылыққа байланысты. Егер өсімдікке ылғал жетіспесе өсімдік түктеніп кетеді, ал егер топырық құрғақтана бастаса, тамыр түктері жойылады және оның орнына жаңа тамыр түктері өсіп шыға алмайды [12] .
Arabidopsis өсімдігінің тамыр түктеріне көптеген ғалымдар назар аударуда. Көптеген зерттеу нәтижелері бойынша Arabidopsis өсімдігінің тамыр түктері клеткалық биология және физиологияның негізгі зерттеулеріне және тәжірибелік зерттеулері үшін де қолайлы болып табылады. Тамыр түктері 1мкм/мин аз емес жылдамдықпен тез өсуге қабілетті, сондықтан оған зерттеулер жүргізу қиындықты туғызбайды.
Тамыр түктерінің дамуы тамырдың ұшындағы қатарласып орналасқан клеткаларда біртіндеп байқалады, тамыр түктері қалыпытасуы барысындағы клеткалық өзгерістер тамыр түктерінің дамуын пайдалы ораймен қамтамасыз етеді [13] .
Негізгі азықтық дәнді дақылдардың, оның ішінде әсіресе, бидайдың және күріштің абиотикалық және биотикалық стрестерге төзімділігін арттыру қуаңшылық климат жағдайларымен сипатталатын Қазақстанның дән егілетін аймақтары үшін маңызды мәселелердің бірі болып табылады [14] . Бидай мен күріш дүниедегі экономикалық және азықтық маңызы бар егіс дақылы. Дүниенің көптеген елдерінде күріш жаңбырлы, ылғалды аймақтарда өседі, сондықтан куаңшылық, судың тапшылығы күрішті өсірудегі маңызды мәселелердің бірі болып табылады. Зерттеушілердің арасында қуаңшылыққа төзімді егіс дақалдырын зерттеуде біраз қиншылықтар бар. Бүкіл дүние жүзінде ғалымдар бидайдың және күріштің куаңшылыққа төзімді сортын шығаруға назар аударуда. Көптеген зерттеушілер былай дейді: күріштің қуаңшылыққа төзімділігі суды буландыру жылдамдылығымен және тамырдың түріне байланысты, өйткені суды буландыру жылдамдылығымен және тамырдың түрі күріштің судың қысымына төзімділік дәрежесін анықтауда маңызды болып саналады [15] .
Дәнді дақылдарда өнімділік бірінші кезекте өсімдіктерде өнімділік көлемінің потенциалымен корреляцияланатын тамыр жүйесінің көлемімен анықталады. Қазіргі уақытта өсімдіктердің тамыр жүйесінің ерекшелігі генетикалық детерминацияланғаны белгілі. Оның сорғыш және метаболитикалық функциясы өзіне тек өсімдіктің өнімділігі жөнінде ғана ақпаратты тасымалдап қоймай, сонымен қатар оның стрестерге қарсы реакциясы жөнінде ақпаратты тасымалдайды [16] .
Судың және қоректік заттардың сіңірілуі негізінен тамырдың созылу аймағының клеткаларымен және тамырдың сорғыш ауданын ұлғайтатын тамыр түктері арқылы жүзеге асырылады [6] . Оптималды жағдайда яғни су және қоректік заттардың өсімдікке сіңірілуі жеңіл болған кезде тамыр түктері өсімдіктердің өсуі үшін абсолютті қажетті болып табылмайды [17], бірақ стресс жағдайында олардың рөлі артады, мысалы стресс жағдайында өсіп жатқан тамыр түктері K + және Сl - сияқты осмотикалық белсенді иондарды белсенді түрде жинақтайтыны жөнінде ақпараттар бар [18] .
НЕГІЗГІ БӨЛІМ
1 ӘДЕБИЕТТЕРГЕ ШОЛУ
1. 1 Тамыр түктеріне жалпы сипаттама
Тамыр түктері тамырдың эпидермис (ризодермис) клеткаларынан түзіледі және жоғары қарай, акропетальдік өсуімен сипатталады. Кейбір жағдайларда тамыр түктері арнайы клеткалардан (трихобласт) түзеледі. Бұл жағдайда түзілген түктер (тамырдың трихомалар) қысқа болып, көршілес эпидермис клеткаларына ұқсастығын білдіреді. Көптеген зерттеу нәтижелері көрсеткендей, ерекше жағдайларда астыңғы, гиподермальды клеткаларда тамыр түктерін өндіруге қабілетті. Әртүрлі өсімдіктерде олардың диaметрі және ұзындығы әртүрлі болады. Г. П. Белостокованың (1981) бақылаулары бойынша тамыр түктерінің формасы бір топқа және бір түрге кіретін өсімдіктерде әр түрлі болады. Автор цилиндр, түйреуіш, спиральды, дихотомды және саңырауқұлақ тәрізді тамыр түктері болатындығын анықтады. Тамыр түктері тамыр бетінің эпидермис клеткаларынан өсіп шығады (сурет 1) [12] .
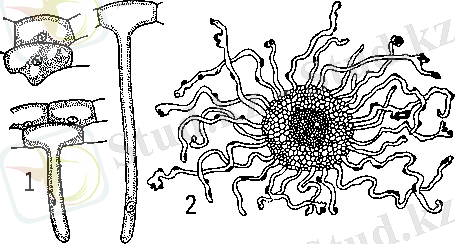
Сурет 1 - Тамыр түктері: 1 - тамыр түктерінің түзілу кезеңдері; 2 - тамырдың тамыр түктерімен көлденең кесіндісі
Арпаның «Lux» сортының дәнін бір тәулік бойы суға бөктіріп, онан соң 1, 5 мМ, pH 3, натрий азийдінде екі жарым сағатқа қойған. Одан кейін суын құрғатып, далалы жағдайда егістікке отырғызған. Зерттеу нәтижесінде 1, 5 мМ натрий азийді бар ортада өскен мутантты арпаның тамыр түгі мен жабайы арпаның тамыр түгі арасында айырмашылықтар болатыны анықталды (2-сурет) [14] .

А
Б
а - жабайы түрінің тамыр түктері; б - 1, 5 мМ натрий азийді бар ортада өсірілген тамыр түктері
Сурет 2 - Арпаның тамыр түктерінің жалпы көрінісі
Тамыр түктері - тамыр эпидермис (ризодермис) клеткасынан туындап шыққан ұзын түтікшелі формада өскен дене. Arabidopsis өсімдігінің тамыр түктерінің диаметрі 1-10 мкм-ге жуық немесе 1 мм-ге дейін өсе алады. Тамыр түктері тамырдың беткі аймағын және тамыр диаметрін кеңейтеді, бұл өсімдіктердің қоректік заттарды қабылдауына және микробтардың өзара әсеріне көмектеседі [19] .
Леваковский Н. тамыр түктері сіңіруші мүше екендігін айтады және тамыр бетінің су және басқа төмен-молекулалық қоректік заттардың өсімдікке тасымалдануын ұлғайтады. Тамырдың 1мм 2 ауданында 1900 данаға жуық түктер болады, олар тамырдың сіңіруші ауданын 5-80мм 2 -қа ұлғайтады. Згуровской Л. Н (1958) ағаштың түріне бақылауы бойынша қайың тамырының ( Pinus silvestris ) 1мм 2 ауданында 272 дана тамыр түктері, ал қылқан жапырақты үйеңкіде ( Acer platanoides ) 1220 дана тамыр түктері болған. Муромцева А. И. (1948) мәліметтері бойынша егілгеніне бір жыл болған Анис ( Malus ) алмасының тамырында жылдың соңында тамыр түктері 3 км-ге дейін тараған. Жүгері тамырының беткі 1см 2 ауданында 425 дана тамыр түгі, ал үрме бұршақта ( Pisum ) 230 дана, ал бидайда ( Tritiсum ) 1000 данаға дейін болады.
Тамыр түктерінің тығыздығы көп жағдайда қоршаған орта жағдайына және қолданылатын агротехникаға байланысты. Debraux G. зерттеулері бойынша егер тамыр түктері жоғары агротехникалық қорда болса, онда олар қант қызышасының ( Beta vilgaris ) тамырының беткейін 10 есе, ал тыңайтпаған жерлерді 40 есе ұлғайта алады. Тамыр түктері эфемерлі түзілімдер, олар бірнеше күн кейде бірнеше апта ғана сақталады. Бірақ, Whitaver E. S (1923) зерттеуі көрсеткендей кейбір Asteraceae түрлері 1, 2 кейде 3 жылға дейін сақталады.
Негізінде тамыр түктері жас, бірақ жақсы дамыған қабықта болады және тамыр ұшында олар болмайды, бірақ эпидермальды (ризодермис) клеткалар жақсы бөліну кезінде болады. Қанағаттанарлық сулы режимде бидайдың, қарабидайдың, арпаның және жоңышқаның ( Triticum, Secale, Hordeum, Trifolium ) тамыр түктері жақсы сақталады және гүлдену кезеңіне дейін сақталады.
Көптеген ағаштарда көп уақыт бойы тамыр түктерімен қатар біріншілік қабаттар сақталады. Ағаштардың көп түрінде тамыр түктері болмайды, бірақ тамыр талшықтарынан алынған эпиблема суды түктері бар эпиблема сияқты сондай жылдамдықпен сіңіреді. Сонымен қатар балшық өсімдіктерінде ( Butoms umbellatus, Caltha palustris, Nymphaea, hippuris vilgaris, Myriophyllum spicatum ; тамыр түктері болмайды, аз мөлшерде Ranunculus repens, Polygonatum odoratum, Allium түрлерінде кездеседі .
Кейінен белгілі болғандай су өсімдіктерінде түктер болмайды, бірақ оларды субстратқа ексе бірден тамыр түктері пайда болады. Жерде өсетін өсімдіктерде тамыр түктері суға тигенде пайда болды .
Тамыр түктері өте кішкентай болады, ұзындығы алмада-0, 1мм, өрікте ұзындау, қарақатта-1мм, қияқта-2, 4мм, дәнді дақылдарда - 1, 2-1, 5мм, орхитде- 3 мм, кейбір өсімдікттерде 10 мм ге дейін жетеді; диаметірі негізінде 0, 05- 0, 17 мм ден аспайды.
Belford D. S, Preston 1961 жылы Sinapis alba өсімдігінің тамыр түгін электронды микроскоппен және рентгенскопиямен зерттегенде, олардың екі қабатты қабығы болатынын анықтаған. Сыртқы қабаты ретсіз орналасқан микрофибрилладан тұрады, құрамында жоғары көлемде аморфты компоненттер (пектин, гемицеллюлоза) болады, олар барлық түктерді қаптайды.
Ішкі қабат арнайы белгіленген целлюлоза микрофибринінен және тамыр түктерінің ұшына жетпейтін бөліктерден тұрады. Тамыр түктерінің қабырғасы шырышты қабаттан, кутиннен, целлюлоза- пектинді тордан тұрады, сонымен қатар тамыр түктерінде саңылаулар мен плазмодесмалар кездеседі. Плазмодесма арқылы топырақ бөлшектерімен байланыс жасалады (3- сурет) .
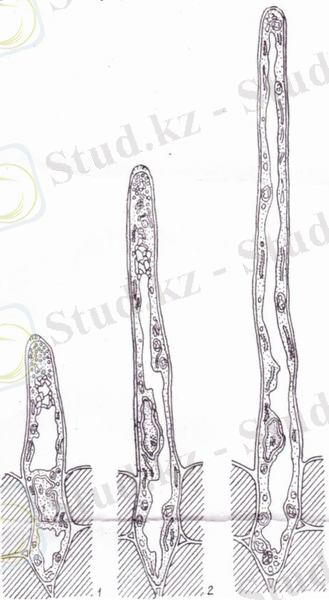
Сурет 3 - Әртүрлі сатылардағы тамыр түктері: 1, 2 - бастапқы, 3- тамыр түктерініің өсіуі тоқтайды.
Тамыр түктері қоршаған орта өзгерістеріне өте сезімтал. Ауа камераларында және ертінділерде жағдай өзгерсе, тамыр түктерінің формасы өзгеріп, өз формасын сақтамайды. Топырақта жағдай мүлдем басқа болады, мұнда тамырлар микроорганизмдермен, топырақ ауасымен, біркелкі емес химиялық және физикалық жағдайлармен бірге пайда болады, сондықтан тамыр түктеріне әр түрлі жағдайлар әсер етеді, олар жуандайды, беткі бөлігі үлкейеді, бағыты өзгереді, кейде бұтақтанады, бірақ әр түрлі бұтақтану және жуандау болса да, тамыр түктері бірклеткалы болып қалады және қайта түзіледі. Тамыр түктерінің протопластары өте вакульденген [12] .
1. 2 Тамыр түктерінің атқаратын қызметі
Тамыр түктерінің гомеостазы мен морфогенезін зерттеудегі соңғы жетістіктер жоғары сатылы өсімдіктердегі гормоналды элиситорлық және басқа жерлерді, сондай-ақ гүлдеу кезіндегі тозаң түтігінің даму теориясындағы, патогенді саңырауқұлақ клеткаларының пролиферециясының теориясында алынған нәтижелерді экстраполилерлеуге мүмкіндік береді. Осы зерттеу барысында алынған мәліметтер бидайдың тамыр түктері жапырақ пластинкаларының түткті клеткалармен бірге тұздану маңайындағы құрғату факторларының әсерін анықтау үшін әрі үлгі болып есептелетіндігін көрсетеді. Тамыр жүйесі түктену клеткасының физиологиялық, биохимиялық қасиеттерін зерттеу біздің экстремальді абиотикалық жағдайда әсер ету кезіндегі бидайдың тұрақтылығы мен өнімділігіне әр түрлі мүшелердің түктенгіш клеткаларының әсері мен әр түрлі түктену типтері туралы түсінігін едәуір кеңейтті. 2008 жылдың соңынан бері тәжірибелік аспектте, Claire Grierson et. al. тобының минаралды заттар мен ылғалды тимді сіңіруге қабілетті, ұзартылған түгі бар сортформаларын қолдану есебінен өнімділікті арттыруға бағыттылған зерттеу жұмыстары жылдам, қарқынды жүруде. Дәл осы әдіс әсіресе энергия мен иілгіш түктердің көзі болып есептелетін өсімдіктер- бидай, арпа. Жүгері дақылдары үшін болашағы зор деп күтеді [20] .
Жапырақ және тамыр түктерін трихобластар деп те атайды. Тамырда маманданған жоғарыда аталған клеткалардың рөлі аса маңызды болып табылады, өйткені олар тамырдың сыртқы ауданында қарқынды көбейіп, өсімдікті сумен және қоректік заттармен қамтамасыз етуге қабілетті. Клетка қабатының негізінде жатқан тамыр эпидермисінде (ризодермисінде) екі түрлі клетка типі маманданған. Эпидермис (ризодермис) клеткалары клетка кабатының бетінде орналасып, тамыр түктерін түзеді. Түктер эмбриогенездің қарқынды дамуының соңғы және жетілудің бастапқы кезеңдерінің аралығында толық қалыптасады.
Тамыр түк клеткалры - суды көп мөлшерде сіңіріп тоқтаусыз жапыраққа жеткізу қабілетіне ие. Ал бұл процесс диффузияға сүйенеді. Тамыр түктері тамырдың топырақпен жанасу беттік ауданын кеңейтетіндіктен диффузияның жылдамдығы артады, өйткені диффузияның жылдамдығы тікелей беттік ауданға байланысты.
Су және минералдар топырақтан тамыр түктеріне өту арқылы ксиломаға барып бүкіл өсімдік денесіне таралады. Тамыр түктерінен ксиломаға сұйықтықтар 2 жол арқылы өтеді. 1) апопласт 2) симпласт [21-25]
1. 3 Өсімдік тамырының құрлысы
Белгілі болғандай, тамырдың құрлысын жоғары сатыдағы өсімдіктерде зерттеу ыңғайлы. Тамырдың артықшылығы оның бөліну зонасы және клеткалардың таралу аймағы бөлек орналасқан. Жапырақтың өсіуін зерттеу қиын, себебі олардың бөлінуі бір уақытта басталады. Бұл қиындықтарды тамырмен жұмыс істегенде жеңу оңай. Олардың апикальды бөлімдері меристамада орналасқан, оның астында клеткалардың бөліну аймағы.
Тамыр аймағын екі әдіспен бөлуге болады:
- Анатомиялық құрылымы негізінде
- Клеткалардың ұзындығын өлшеу негізінде
Сіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz