Микроорганизмдер генетикасы: бактериялар мен вирустардың құрылымы, көбеюі және генетикалық механизмдері

1 МИКHООРГАНИЗМДЕР ГЕНЕТИКАСЫ
Адамның көзі ең алғаш бактерияларды көргеннен бері үш ғасыр уақыт өтті. Ал микробиологияның өз алдына дербсс ғылым болып бөлінуі үшін тағы екі жүз жыл қажет болды.
Аса ұсақ тіршілік иелеріне биохимия, биофизика, микробиологиямен қатар генетика да ерекше көңіл бөліп отыр. Молекулалық биология молекулалардың өзара қарым-қатынасын зерртеу үшін, вирустар мен бактериялардың өте жақсы модель болып табылатынын анықтады. Кейбір вирустардан және бактериофагтардан басқа барлық микробтар клеткалы организмдер болып табылады. Егер жануарлар мен өсімдіктердің басым көпшілігі көп клеткалы организмдер болса микробтар әлемі әдетте бір клеткалы болып келеді. Бактериялар, ашытқы саңырауқұлақтар (дрожжи), кейбір микросаңырауқұлақтар, қарапайым организмдер және көптеген балдырлар - осылардың бәрі микроорганизмдер әлемінің бір клеткалы өкілі - прокариоттар деп аталады. Олардың клеткасынын, дамуы ең қарапайым эуукариоттардан да (жоғары сатыдағылар) төмен ұйымдасқан. Мик-роорганизмдерде миточ да мейоз да болмайды, ал клетка құрылысында ешқандай компартментлизация құбылысы (клетканың ядроға, ядрошыққа және цитоплазма органоидтарына бөлінуі) жоқ, Электрондық мнкрофотографияларда прокариоттар клеткасының екі түрлі құрылымдылық бөліктеп тұратынын көруге болады. Олар цитоплазма, нук-леоплазма немесе нуклеоид. Цитоплазмамен нуклеоплазма арасында мембрана болмайды.
Бактериялар өте тез көбейеді. Мысалы оның аналық клеткасы ор 20 минут сайын екі жас клеткаға бөлінеді. Аз уақыттың ішінде олардан миллиондаған ұрпақ алуға болады. Сонымен қатар хромосомалар жиынтығы гаплоидты (дараланған) түрде болғандықтан олар әрі гаметаның әр дербес организм міндетін қоса атқарады. Міне осының бәрі вирустар мен бактерияларды геннің нәзік құрылымын зерттеуде табылмайтын генетикалық объект ретінде қолдануға мүмкіндік береді.
Генегикалық зерттеулерде қолданылатын классикалық обьектілерге бактериялардан ішек таяқшасы (Escherichia coli, Salmonella) туыстас бактериялар, нейроспора, ал вирустардың ішінде, осы аталған бактерияларды зақымдандыратын, бактериофаттар мен темекі теңбілінің шарусы (қысқаша ВТМ - вирус табачной мозаики) .
Бұлардың қызметін генетиктер 40-жылдардан гапа бастап пайдаланды. 1944 жылдан 1952 жылға дейін бактерияларда генетикалық материалдың қосылуын және рекомбинациясын тудыратын үш маңызды процестер ашылды, олар - трансформация, конъюғация және трансдукция.
БАКТЕРИЯЛАР МЕН ВИРУСТАРДЫҢ ГЕНКТИКЛЛЫҚ ЕРЕКШЕЛІП, ҚҰРЫЛЫСЫ ЖӘНІЕ ӨСІП-ӨНУІ
Кейбір вирустардан және бактериофагтардан басқа барлық микробтар бір клеткалы организмдср болып табылады. Сөйтіп микробтар жайында да олардың аталық аналық, ұрпағы жөнінде айтуға болады (бактерияларда "аналық" клеткалар және жас клеткалар деп айту қабылданған) .
Өсімдіктер, жануарлар және микроорганизімдердің бәріне бірдей ортақ өте маңызды белгі - олардың клеткалық құрылысы. Клетка дегеніміз тірі материяның ең ұсақ және лайықты белгілі орта жағдайында өз бетімен тіршілік ете алатын бөлігі.
Микробтар әлемінде жеке клеткалардың қозғалыс, зат алмасу, көбею, сыртқы әсерге жауап беру сияқты негізгі тіршілік әрекеттерін дербес түрде атқара алу қабілеті бар екендігі жайлы көптеген фактілерді кездестіруге болады.
Орташа бактериялар клеткасының ұйымдығы миллиметрдің мыңнан бір болігіндей ғана болады. Сондықтан олар үшін өлшем бірлігінің өте ұсақ түрін: микрометр (мкм) немесе миллиметрдің мыңнан бір бөлігін, наиометрді (нм) миллиметрдің миллиондық бөлігін пайдаланады. Ал электрондық микроскоппен жұмыс істеген кезде бұлардан да ұсақ өлшемді - ангетремді (А) немссе миллиметрдің он миллиондық бөлігін қолданады.
Сонымен: 1 мм = 1000 мкм = 1 нм = 1 А°. Бұларда барлық басқа клеткалар сияқты протоплазмасы Г>ар, ол цитоплазма мен ядродан (бактерияда көбінесс ядро аймағы деген айтылады) құралған. Цитоплазманың өзі цитоплазмалық мембранамен қоршаулы болады, мембрананың сырт жағы клетка қабысымен түйеседі, клетка осы клетка қабығына байланысты болады. Бактерия клеткасына пенициллинмен әсер еткен кезде әдетте оның клетка қабығының құрылысы өзгереді де цитоилазмалық жұка мембрана ғана калады.
Клетка қабығының қорғаныш қызмет атқаратыны 1-ші тараудан белгілі, сонымен бірге ол әр түрлі бактериялардың өздеріне ғана тон тұрақты пішін береді. Егер бактерия таяқшасының қабығынан оның ішіндегі затын бөліп ажыратса, онда иротопласт (протоаласт - цитоплазмалық мембранамен шектеген протоплазма) таяқша пішінінен айырылады да шар пішінде болып қалады, бұл бірақ өзінің барлық тіршілік қызметін сақтап қалады. Вирустар мен фагтар бактерияларда (бактериофагтар), өсімдіктер мен жануарларда (вирустар) паразиттік тіршілік ететін клеткасыз формалар болып табылады. Оларда хромосоманың ролін ДНҚ немесе РНҚ молекуласы атқарады. Бактериялар (прокариот) аралық орын алады. Вирустарға қарағанда бактериялар едәуір күрделі құрылымды келеді, бірақ олардың ядросы цитоп-лазмадан бөлінбеген, ал хромосоманың қызметін мұнда да ДНҚ жіпшелері атқарады. Клетка ұршық түзбестен бөлінеді. Алайда микроорганизмдерде молекулалық генетиканың объектісі болатын жалпы ерекшеліктері бар. Тіршілік циклінің қысқалығы негізгі ерекшелігі болып табылады. Мысалы, көптеген бактериофагтарда, вирустар мен бактери-яларда бір бөлінуден екінші бөлінуге дейінгі уақыт 20-30 минут, саңырауқұлақтарда 1-2 сағат, ал хлорелла балдырында 1 тәулік болады. Олардың мұндай ерекшелігі қысқа мерзім ішінде орасан көп ұрпақ алуды қамтамасыз етеді.
Микроорганизмдердің келесі ерекшелігі - өте жылдам көбейетіндігі - бір мезгілде орасан көп мөлшерде особьтар алуға болады. Бұл миллионға біреу және одан сирек кездесетін генетикалық құбылыстарды (мутацияларды) та-буға мүмкіндік береді. Көптеген микроорганизмдерде көбеюдің екі әдісінің (жыныссыз және жынысты) болуы - үшінші ерекшелік. Бұл жынысты жолмен көбею барысында рекомбинациялар алуға, ал жыныссыз көбеюде мейоздан соң, гаплофазада тікелей рекомбинация өнімдерін зерттеуге мүмкіндік туғызады.
Мұның бәрі генетикалық талдаудың шешуші қабілетін айтарлықтай арттырады.
ВИРУСТАРДЫҢ СИПАТТАМАСЫ. Өзінің көлеміне қарай вирустың бөлшектері ең кішкене тірі клеткалар мен химиялық қосылыстардың ірі молекулалары арасынан орын алады.
Бактериофагтар формасының ерекшелігімен көзге түседі. Мысалы, фаг Т2 - ішек бактериялары паразитінің формасы шокиарга ұқсайды. Клеткаға шабуыл жасағанда өзінің құйрық тәрізді қосалқысымен жанасады.
Вирустардың белок пен нуклеин қышқылдарынан (нуклеопротеид) тұратыны дәлелденді. Нуклин қышқылдарының екі типі табылды. Бірқатар вирустарда тек қана РНҚ, басқа біреулерінде - тек қана ДНҚ.
Тіршілік құбылысы қандай деп сұраққа Пиолопар былай жауап қайтарады: әрбір тірі организімнің өзіне тән формасы, шамасы, ішкі және сыртқы қасиеті Гюлады органдардың бейімделуі осыларға байланысты тірі организімге қозғалыс, сыртқы тітіркеніске әсерлему, өcу, зат алмасу процесі, өсіп-көбею қабілеттілігі сиаяқты аса маңызды ерекшелік тән. Өсіп-көбею арқылы тұқым куалау өзгерісі болады. Осы тұрғыдан вирустарлы алып қарап бұл жәндіктердің тірі, я тірі еместігін түсінуге әрекет жасап көрелік. Химиктерде олар - кристалка айналуға қабілетті ірі молекулалар. Олардың тірі организмдерге топ белгілері бар, өсіп-көбейе алады (бірақ тек қана тірі клеткалар арасында) және соңғы уақытта дәлелденгендей, тұқым қуалау өзгерістеріне ұшырайды. Вирустардың тек қана белгілі бір формасы мен мөлшері ғана емес, сонымен бірге белок пен нуклеин қышкылының белгілі бір құрылысын байқататын ішкі қасиеті де болады. Дегенмен клеткалармен салыстырғанда ол анағұрлым қарапайым. Сондай-ақ ең ірі вирустар мен ең ұсақ бактериялардың арасында көлемі жағынан бірқатар айырмашылық бар.
Сонымен вирустар жанды зат па, әлде жансыз ба? Америка биохимигі У. М. Стэнли бұл сұраққа былай деп жауап қайтарды: "Олар жанды ма, я жансыз ба - бұл жөнінде қойылған сұраққа шын мәнінде қанағаттанарлық жауап ала алмастан шексіз таласуға болады. Бір жағынан алғанда вирустар тірі организмдерге, екінші жағынан - әдеттегі химиялық молекулаларға ұқсас, бірақ біріншіден де, сондай-ақ екіншіден де айырмашылығы бар. Олардың екі жақтылық бейнесі және біздің толық зерттеуімізге боларлық бір сылырғы қарапайым құрылымы, бізге бір жағынан - осім-кобеюге қабілетті химиялық молекулалар тұрғысынан қарастыруға мүмкіндік береді. Сөйтіп біз басқа да барлық тірі организмдерде өтіп жататын өсіп-көбею процесінің химиялық мәнін түсінуге жуықтап келеміз".
Вирустардың аса ірі молекулаларының, сонымен қабат тірі материяның ең кішкене формасының бір ғана жалпы ерекшелік белгісі бар: олардың бәрі де тірі оргамизмдер клеткаларында тіршілік ететін жәндік болып табылады. Вирустардың, бактериялардың паразиттерінің ішінен бактерио-фагтардың бірнеше типтері белгілі.
Темекі теңбілінің вирусы (ВТМ) өзге вирустардан гөрі мұқыят зерттелген.
Рибонуклеин қышқылы вирус бөлшегінде қарапайым шиыршық түрінде соңғы белоктың макромолекуласы болып саналатын цилиндрде тасаланған. Бөлшектің ішінде қуыс "капал" бар. Бұл камандың диаметрі 4 нм. Белоктың макро-молекуласы ұсақ пептид тізбектерінен (суббірліктерден) тұрады, олар ВТМ-ның әр бөлшегінде орта есеппен 2200-ге жуықтайды. РНҚ спиралінің үстінен жабатын "жамылғыш' тәрізденген осы тізбектер неғұрлым ірі спираль болып табылады, оның сыртқы диаметрі 17-18 нм тең. Спираль 360 градусқа айналған сайын белоктық "жамылғышқа" 16 1/3 жаңа пептид тізбектері қосылып, оның ұзындығы 2, 3 нм ұзарады. Орамдардың жалпы саны 130-га жуық. 23123131313131
Белоктар құратын тізбектер химиялық түрғыдан әр-дайым біртскті. Олардың қүрылымының орбір суббірліктері нсигидтік байланыстармсн нолипеіітидтерге біріктірілген амип қышқылдарының тізбсктерінен түрады. Осындай бір суббірліктс амин қышқылдарының 168 молекуласы кірсді.
Әрбір суббірлікте ортүрлі 16 амин қышқылы бар. Олардын, атаулары, қысқартылған символдары және суббірліктегі молекулалардың саны төмендегі кестеде келтірілген.
РНҚ-ның негізгі суббірліктері нуклеотидтер болып та-былады. Осы қосылыстың төрт типі бар: аденил, гуанил, цистидил және уридил қышқылдары. РНҚ-ның бүкіл ши-
6-кесте
ыршығьшда 6500 иуклеотид болады, бірак. опып калаіі ор-наласатыны олі дс болса бслгісіз.
ВТМ-ныц бүкіл молекуласы мі. іиіід; ііі Гніоіүіідік элемеиттсрдіц: коміртегі, сутсчі, огісіі, ; ііоі. фоіфяр ж; ліс кукірттің шамамен 525 атомыікпі турады. 5 млн 250 мың атом РНҚ молекуласындаш 6500 пуклсотпд іісіі бело-ктардың 2200 суббірліктсрі арасыида болііпсн, Оүлардыц оркайсысыида 158 амин к. ышкылі>ііп>щ калдыкгары Gap. Құрамы мен қүрылымыиың осыидаіі күрдсліліі іпс қарама-стан, ВТМ-ның үзындығы неборі 0, 0003 милліімстріе, ал ені - 0, 18 миллиметрге тең.
ВТМ-re үксас шиыршықты күрылым копісісп one вирустарда да бар. Әзірге белгілі иирустардыц ішііідегі сң үсағы - бактериофаг ф + 174, оныц тек қапа 12 үстіцгі суббірлігі бар.
Жеке вирус бөлшегі - осы вирустыц нсгізгі инфекци-ялық бірлігі. Ол белоктық "қабыктан" жоне омыц ішіндс жасырын жатқан нуклеин қышқылы молекуласыиан түра-ды. Вирустар бөлшектерінің клеткалардан тагы бір айырма-шылығы - оларда ордайым нуклеин қышқылыныц (ДНК, РНК) екеуі де бактериялардың жонс басқа организмдсрдіц клеткаларындағы сияқты әрбір вирус бөлшсгіндс мацызды "басқару" қызметін атқарады.
БАКТЕРИОФАГТЫҢ ӘРЕКЕТІ. Вирустың клеткапі қалай отетіні жоне онда бүлдіру озгсрістеріп қалаіі жүргізетіні женіндегі моліметгср алгаш рет бактериофапа-
рга - ішек бактермисыііыа (Е. соіі) жауларыпа бакылау жүргізілгсн
ксідс
алыпды. Элсктромдық микроскоп Т4 бактсриофаі ыныц сыр-ткы күрылымыпыц сырілп ашуга жордем тигізді. Фаі бөлшсгінің басы боладі. і, ол екі жагынан қакмақшалары бар алты қырлы призма тәрізді. Басынан үзынша келген қүйрық қосалк. ысы кетеді, оның үшында жіп тәрізді бірнеше "аяқшалар"
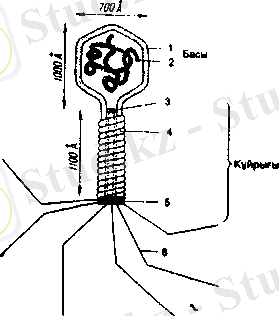 бар. Қосалк. ының каналы
30-сурет.
Т-жүп фагының күрылысы. оны баспсн жалгастырады 1 - белок; 2 - ДНК; 3 - куыс; 4 - кдбыгы; ЖӘНС СОЛ Каналда ДНҚ-5 - гексогональдык табакша; б - күйрық
Н
ЫҢ МакрОМОЛСКуласы бо-жіпшелері.
бар. Қосалк. ының каналы
30-сурет.
Т-жүп фагының күрылысы. оны баспсн жалгастырады 1 - белок; 2 - ДНК; 3 - куыс; 4 - кдбыгы; ЖӘНС СОЛ Каналда ДНҚ-5 - гексогональдык табакша; б - күйрық
Н
ЫҢ МакрОМОЛСКуласы бо-жіпшелері.
лады. Фаг к, абыгыііын калгаи болігі бслоктық жаратылысқа жагалы (30-сурет) .
Г2 бактсриофагыныц імым монінде к. озгалыс органда-рі. і Аоііс оргадаіы молекулалармсн сок. гыгысуы салдарыиан сүііык. оргада орец жылжиды. Соіігіп ол бактериямен жана-сады. Фаі' оныц сыргына "жіпше аяк, гарымсн" жабысады. Қүйрык. к. осалк. ысьшым үшыида бактерияның қабығын за-қымдаіііьіп жоис оны "ксміріп" шагын тесік жасайтын фер-мснт болады. Осы операциядан ксйін қосалқы фаг дснесіне таргылады (гармон тәрізді созылып жиырылады) .
ДНҚ бактериофагтың басынан қүйрық қосалқысының каиалы арқылы жөне бактсрия ьсабыргасында пайда болған тесік арқылы клеткаға енеді. Осының бәрі шприц арқылы дорі жіберу процедурасын еске түсіреді. Бас пен қосалқы-пың бслоктық қабыгы сыртында қалады, ал бактерияның клсткасына ДНҚ ғана енеді. Сөйтіп фаг ДНҚ-ның оздігінен осіп-көбеюі басталады. ДНҚ-ның макромолекула-сы бактсрияның "шикізаты" мен бүкіл ферменттік аппара-тын пайдалана отырып, өзінің есіп-көбею процесіне қаты-» суға можбүр стсді. ДНҚ-ныц бастапқы макромолекуласы-нан екі туынды пайда болады және тез арада-ақ олардың саны 8-Іб-ға дейін т. т. көбейс түседі. Жарты сағат өткен сон фаг ДНҚ-сының бастапқы молекулаларының бәрінен клсткада опың '50-300 үрпағы болады, бүлар бактериофаг-тыц оасі, і мсн күйрық қосалк, ысы үшін бактериянын, белок жимактауына можбүр етеді. Клетка әуе шары тәрізді жары-лып кстеді, сол кезде қоршаған ортаға 150-300 бактериофаг гүссді, сойтіи олар дсреу кслссі бактсрияларға шабуыл жа-саііды. Ьүл процссс қайталапады жоне тағы бір жарты сагат откси еоң ортада ондаган мыц фаг пайда болады. Біраз уак, ыг откеннен кейін онда бірде-бір тірі бактерия қалмай-ды тск жсціскс жеткем бактсриофаггар қаптап кетеді. Элек-троидык. микроскоппен қараганда бактериофап-ың бактсри-ямси жаиасуыи, фагтың қандай бөлігі клеткаға енетінін жоие сыртқы жағында нс қалатынын айыру қиын. Осыған толмқ коз жеткізу үшін дол зерттеулер жүргізу керек.
1952 жылы екі американдық зерттеушілер А. Херши мсн М. Чейз радиоактивті изотоптарды (фосфор 32 Р мен күкірт 3<і 5 бір мезетте қолданып тәжірибе жүргізді. Фосфор тск қана бактериофагтардың ДНҚ-сында кездеседі, ад күкірт олардың белоктарының қүрамына ғана ене алады.
Бактсриялар жиыны биогсидік фосфордың көзі ретінде тск қана І5 Р болған қоректік ортада өсірілді. Іс жүзінде бак-тсриялар радиоактивті фосфорды жай фосфордан айыра ал-
майды жоне І2 Р бар ортаны пайдаланады. Осыилаіі жолмеп атомдар 32 / > -дан бактериялық клсткаларга түссді, бүдап кейін олар бактериофагтыц олжасына аііналуі. і мүмкіп жонс Ъ Р бактериофагтар ДИК-сыныц күрамыпа иіеді. Осыгам үқсас тәжірибе радиоактивтік қосмлыстарм күкірт ! \У Сю- лып саналатын басқа пробиркада қайталаііып жопс бслоіы-на 33 5 күкірті бар бактериофагтар алынды. Соңыиап осы фагтар радиоактивті қосылыстары жоқ ортада осіріліен бак-терияларға жұқтырылды. Алғашқыда радиоактивті күкірт бар бактериофагтар пайдаланылды. Жүқтыру процссі бірнеше минуттың ішінде аяқталды, одан ксйін иситрифуга арқылы зақымданган бактерияларды бактериофаггьщ қал-дықтарынан бөліп алып, олардың радиоактивтігін анықта-ды. Бактерияларда ешбір радиоактивті белгі болгаи жоқ, ал "талқанданған" фагтарда (бастары мен құйрык қосалқыла-ры) радиоактивті белгі анағүрлым котеріңкі болды. Демек, бактерияға фагтан тек қана ДНҚ (күкірті болмайтын) снді, ал күрамына 35 S кірген белок клеткалардап тыс қалды. Осыдан кейін өзініц ДНҚ-сында радиоактпнті фосфор (фагтардын белоктарында фосфор болмайды!) Полатым фаг-тармен тожірибе жасалды. Бактсриялар зақилідангашіап кейін бірнешс минут өткен соң фагтардыц қаллмқтарі. і мсн закымданган бактериялар тап сондай одісисн поліиіи алы-нып, радиоактивтігі зерттелді. Бүл жолы ол бактсриялардын клеткаларынан табылды! Сонымен бактсриялардм фагтар-мен зақымдандырғанда, бактсриялық клеткалардын ішінде фагтың ДНҚ-сы енетіні (онда фосфор болады), ал сыртқы жағында онын белоктық қабыгы қалатыны (оида күкірт болады) айкын дәлелдснді.
Бірқатар жагдайларда бактериофаг микроб клеткасында пайда болғанда, клетканың қүрып кетуі міндет смес. Фаг-тың ДНҚ-сы клеткаға енсді, бірак. ешнорсс болмагандай, клетка бүрынғысынша тіршілік ете береді. Закымданған бактерия былай қарағанда ешбір кедергісіз өсс бсріп, қолай-лы жағдайларда екі жаңа клеткаға бөлінеді. Бірак. оныи касиеттерінде өзгерістер болады - бактерия бслгілі бір типтегі фагтарға төзімділік көрсетеді. Осыныц өзі оның клеткасында фаг ДНҚ-сының болатындығына бірдсн-бір куә болып табылады. Өзінің хромосома қүрамында профаг кіретін бактерид клеткаларын лизогенді, ал бактерия мен профаг ДНҚ-сының бірігіп тіршілік етуін лизогения деп атайды (грекше "лизис" - еріту, бактерия клсткасыиың күйреуі) . Лизогенді деп айтылатын себебі, бслгілі бір ксчсң-де белгісіз себептен миллиард клеткадаи 2-3 клстка оііиіп
мсймаидостык. корсеткені үшін күтпеген жерден жазасын іаргып, күрыіі кетсді. Ксйбір клсткалар бактериофаітардың шабуылыпа шыдайды. Бірақ алғашкы соққыға төзген бұл "галаііқорлар" озгсріп, жана қасиеттергс ие болады жоне оларды келесі үрпақтарға ауыстырады.
Ьүл к. ү6ылысты алгашқы рст 1952 жылы байқаған Дж. Ледеберг оыы былай түсіндірді: Фагтың ДНҚ-сы лизогеңді бактсрияга сигсннен кейін фагтар одеттегідей өсіп-көбсймейді және клеткалар да қүрымайды. Бүл жағдайда фагтың ДНҚ-сы клетканың хромосомасындағы бактерия-лык ДНҚ-ға жалғасады. Осы клетка екі туынды клеткауа болінс бастағанда, хромосома бөлініп, фагтың саны екі есе көбейеді жоне оның жартысы жаңа хромосомалардың әр^-айсысына түседі. Сөйтіп, клеткалар одан әрі бөлінген сайіцң фагтыц ДНҚ-сы орбір туынды клеткаға ауысады. Әриңе, миллиардтаған клеткалардың ішінде фагтың ДНҚ-сы хро- мосомадан босап, қаркынды күйде өсіп-көбейетіндері де болады, ал клетка қүрып кетеді. Олардың кейбіреуіндеи фагтың ДНҚ-сы клеткалы хромосоманың бір бөлшегін озімен бірге алып кетіп, фагтың жаңадан пайда болған болшсктсрінде, оз ДНҚ-сынаи баска бактсриялық хромосо-маның "кішкене бір кесегі" болады. Мүндай фаг басқа бір беііімделгіш клеткамен ксздссксндс, фагтың ДНҚ-сымен біріс оган бактериялық ДНҚ-ның бөлшегі түседі және олар бірлссіп жаңа клетка-иеніц формасына жалғасады. Бөтен хромосоманың фрагмснті осы клетканың хромосомасымен араласып соңғы жаңа гендермен толығады. Сырттай қара-ганда бүл клстканың ойтеуір бір жаңа қасиетке ие болғаны байқалады. Айталық, ботсн ген бүған дейін "толыққан" клсгка түзс алмаған заттың жинақталуын анықтаса, онда бүл қабілеттілікті оның езі ғана емес, сонымен бірге оның үрііагы да алады. Түқым қуалаушылык. өзгерістің мүндай формасын Ледеберг трансдукция деп атады.
ТРАНСФОРМАЦИЯ
Трансформация қүбылысы женінде бүдан бүрынғы та-рауда соз к. озғалған. Онда біз пневмококктар колониялары-ның диссоциацияға душар болатынын, яғни олардың қасиеттерінің өзгеретінін байқадық. 5 формалы пневмокок-ктар клеткалары өте жүқпалы (екпе қабынуының типті формасын туғызады) . Олар әдетте жүп клеткалар бүркей^ кілегейлі капсулалар түзеді. Бүл капсулалар күрделі поли- сахаридтсн түрады. Керісінше S формасының өзгеруі іютижссіндс пайда болған R формалы пневмококктар
жүқіталы емес, капсулалар гүзбейді жоис жүптасып орла-ласпайды. Бұл айырмашылықтар жапурларги жасалган тожірибелер кезінде (жүқпалылығын айыру), микроскоппен зерттегенде және коректік ортада - агарда осіргсыде коло-ниялардың формасы мен түсінен байқалады.
... жалғасыСіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz