Өсімдіктерді клондық микрокөбейту: әдістері, артықшылықтары және технологиялық кезеңдері

3. Өсімдіктерді клондық микрокөбейту
3. 1. Өсімдіктерді клондық микрокөбейтудің пайдасы
Маманданған үлпаның кез келген тірі клеткалары лайыкты қоректік ортада өсіргенде, өздерінің тотипотенттік касиетін жүзеге асырып, регенерация арқылы бүтін өсімдікке айнала алады. Жеке клеткалардан сол өсімдік түріне тән барлық белгілері мен касиеттері сакталған бүтін өсімдіктін тұзілуі клондық көбейту технологиясының негізін калайды. Клон деген жыныссыз жол-мен, яғни вегетативтік көбею жолымен, тұзілетін организм.
Өсімдікгердің клондық мшфокөбеюі деген өсімдіктердің іп үйго жағдайында жыныссыз жолмен көбеюі. Соңың нәтижесівде пайда болған өсімдіктер бастапқы өсімдікпен және өзара бір-бірімен генетикалық түрғыдан айнымастай бірдей болады.
Бұл биотехнологиялық әдістін дағдылы вегетативтік жолмен көбеюмен салыстырғанда бірталай артықшылықгары бар, атап айтканда:
1. Көбею коэффициенті өте жоғары. Мысалы, гербера, бүл-дірген, хризантема, раушанның бір өсімдігінен іп үііго жағдайывда бір жылдың ішінде 1 миллионнан астам клон өсімдіктер алуға болады. Алма ағашының бір бүршігінен 8 айдың ішінде 60 мыннан астам өркен шығады. Танкүрайдың бір бүтадағы меристемаларын бөліп алып өсіріп жылына 50 мьщға дейін өсімдік алуға болады. Соңымен, микрокөбеюдің коэффициенті баска вегетативтік көбею әдістерімен салыстырғанда мындаған есе артық.
2. Микрокөбеюмен катар өсімдіктер вирустар мен патогендік микроорганизмдерден сауыкгырылады.
3. Сұрыптау процесін жылдамдату. Жана сорттарды тез көбейтіп, оларды ауьш шаруашьшык өндірісінде пайдалану мерзімі едәуір кыскарады.
4. Вегететишікжолмен көбейе алмашын өсімдіктерді мысалы, пальманы тек іп үйго жағдайында көбейтуге болады. Осы эдіспен өнеркәсіп денгейінде біркатар өсімдіктерді көбейтеді.
5. Үнемділік. Арнайы бөлмеде стеллаждарда орналаскан пробиркаларда жыл бойы мындаған өсімдіктерді өсіру арқылы теплицалар аланы үнемделеді.
6. Жас өсімдіктерді алу, яғни кәрі даракгарды жасарту.
7. Өсу процесінжыл бойы үзбеуге болады, әсіресе Бұлдамуын-да тыңыштық кезені болатың өсімдіктерді көбейтуге тиімді.
3. 2. Клондық микрокөбейтудің әдістері
Клондық микрокөбейтудің түрлі әдістерін ғалымдар ол кезде өтетін морфогенездің өзгешеліктеріне карай жіктейді. Н. В. Катаева мен Р. Г. Бутенко былай жіктеуді үсынады: а) бүрыннан болған меристемалардан өскен өсімдікгер; б) жаңадан пайда болған меристемалардан өскен өсімдіктер (18-сурет) .
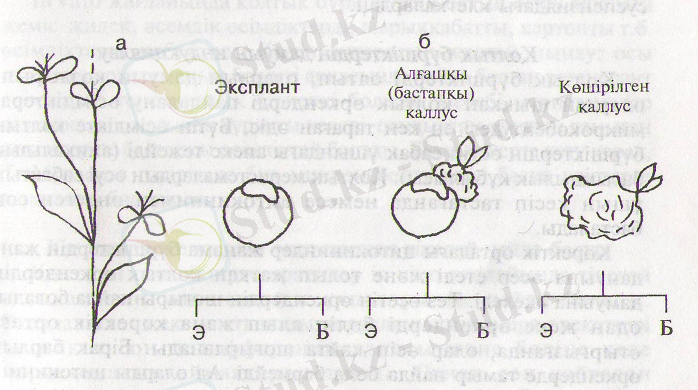
18-сурет. Өсімдіктерді клондық микрокөбейту әдістерінің жіктелуі:
а - бар меристемаларьш активтеу; б - бүршік (б) пен
эмбриоидтардың (э) пайда болуы (16) .
Бірінші типті өсімдіктер бүтін өсімдікте бүрыннан болған меристемаларды (сабактьщ апексі, колтық және бүйыккан бүршіктері) активтендіру жолымен пайда болады. Бұл меристемадан шыккан өсімдіктер генетикалық жағынан аналық өсімдікпен пара-пар, өйткені апекстерді in vitroжағдайында өсіргенде олар генетикалық тұрақтылығын сактайды.
Екінші типті өсімдіктер in vitroжағдайында пайда болған бүршіктер мен эмбриоидтардан алынады. Бұлөсімдіктерде маманданған және каллус клеткаларынан шыккандығына байланысты генетикалық өзгергіштіктер орын алуы мүмкін. Соңдықтан, шыккан клондар бастапқы өсімдіктен біршама ауыткып кете береді. Сөйтіп Бұләдісті тек каллустары түрақты немесе регенеранттарда пайда болған өзгерістер табиғи өзгергіштіктен аспайтың өсімдіктерге пайдалануға болады.
Бүршіктер мен эмбриоидтар былай пайда болады: 1) экспланттың маманданған клеткаларынан тікелей (көбейме мүшелерінің үлпаларынан, эпидермистан, субэпидермис үлпаларынан, жапырак мезофилінен т. б. ) ; 2) экспланттан шыккан алғашкы каллустан; 3) көшіріп отырғызған каллустан немесе суспензиядағы клеткалардан.
Колтық бүршіктердің дамуын индукциялау
Қолтық бүршіктерді оятып, олардың дамуын коздырьш, олардан шыккан колтық өркендерді пайдалану өсімдіктерді микрокөбейтудегі ен кен тараған әдіс. Бүтін өсімдікте колтық бүршіктердің өсуін сабак үшындағы апекс тежейді (апикальдық басымдылық құбылысы) . Колтық меристемалардың өсуі сабактың үшын кесіп тастағанда немесе цитокининмен өндеген соң басталады.
Қоректік ортадағы цитокининдер жанама бүршіктердің жан-дануына әсер етеді және толып жаткан колтық өркендердің дамуына әкеледі. Тез өсетін өркендердің шоғыры пайда болады, одан жеке өркендерді бөліп алып жана қоректік ортаға отырғызғанда, олар өсіп кайта шоғырланады. Бірақ барлық өркендерде тамыр пайда бола бермейді. Ал оларды цитокинині жок ортаға көшірсе, өзінен-өзі тамырлары өсіп шығады, әсіресе даражарнакты өсімдіктерде.
Колтық бүршіктерді ояту үшін қоректік ортадағы
фитогормондардың концентрациясын дүрыс аныктау керек. Цитокинин жоғары мөлшерде қолданылғанда колтық өркениіц дамуына мүмкіндік туғызады, Бірақ өсімдіктін морфологиясына теріс әсер етіп кемістіктері бар формалар пайда болады. Қоректік ортада ауксиннің мөлшері артық болса, каллус тұзілуі мүмкін. Каллус меристеманың өсуін тежейді немесе бастапқы өсімдіктен генетикалық айырмашылықтары бар косымша сабақ апекстерінің пайда болуына әкеледі. Ондай жағдайда ортада ауксиннің концентрациясын төмендетеді немесе тіпті оны коспайды.
Соңымен колтық бүршіктері бар сабак кесінділерін лайыкты қоректік ортада өсіріп, өркен алуға болады, ал оны күрамы баска ортада тамырландырып бүтін өсімдікті шығарады/^Көбеюді жылдамдату максатымен алғашкы өркенді 5-7 жапырак шыккан соң калемшелейді. Әрбір калемшеде бір жапырак болуы керек. Калемше биіктігі 1-1. 5 см болады. Оларды бір-бірден қоректік орта күйылған пробиркаларға отырғызады. Бұлтәсілді микрокалемшелеу деп атайды. Ол жүзім, картоп т. с. с. өсімдіктерге қолданылады. Микрокалемшелеуді 2-3 реттен артық пайдалануға болмайды, одан кейін өркендерді тамырландыру керек.
In vitroжағдайында колтық бүршіктерін оятып өсіру әдісімен жеміс-жидек, әсемдікөсімдіктерді, кырыккабатты, картоптыт. б. өсімдіктерді көбейтеді. Көніл аударатың жағдай мынау: осы әдіспен меристеманың бөлінуі аркасында пайда болған регенерант өсімдіктер генетикалық біркелкі болады. Себебі меристемалар генетикалықжағынан тұрақты келеді, совдықган олардан дамыған өркендер де бастапқы өсімдіктей болады. Бірақ ескерте кету керек, Т. Мурасиге пікірінше Бұлмикрокөбейту әдісі баска әдістерге карағанда бәсенірек.
Косалкы өркендердің экспланттан тікелей пайда болуы
Көптеген өсімдіктерде өркендер іп үііто жағдайында тікелей экспланттыи маманданған үлпаларынан пайда болады. Экспланттың жеке клеткалары корреляциялық өзара әрекетгері бүзылған соң дедифференцияланады және бөліне келе меристемалық, аймактарды тұзеді. Соңан соң кайтадан дедифференцияланып, өркен бүршіктері пайда болады. Сол бүршіктерді тамырландырып, бүтін өсімдікті алуға болады.
Бұл әдіс әсіресе шөптекті өсімдіктердің жапырактарын, пиязшықтарын, пиязшык түйнектерін, сабақтарын, тамырсабақтарын және түйнектерін пайдаланғанда жарамды келеді. Эксплантты ауксин мен цитокининнің шамалы мөлшері бар қоректік ортада өсіргенде бір шоғыр өркендер пайда болады. Оларды бір-бірінен ажыратып бөліп, әрқайсысын жеке ыдысқа отырғызып, тағы да бір топ шоғыр өркендерді алады. Одан кейін өркендерді тамырландырып, бүтін өсімдіктерді шығарады.
Пиязшықты өсімдіктердің көптеген түрлері табиғатта өте баяу көбейеді. Пиязшықтың түрлі ұлпаларынан іn vitro жағдайында қосалкы өркендерді өсіріп көбейту коэффициентін айтарлықтай аттыруға болады. Өте кішкентай экспланттарды қолданып, бір пиязшыктан көптеген біркелкі регенерант өсімдіктерді алуға болады. Осы әдіспен бірқатар өсімдіктерді микрокөбейтеді, атап айтқанда: шегіргүлді, фрезияны, амариллисты, лалагүлді, петунияны, шашыратқыны, каланхоэны, цикламенді т. б. Соңымен қатар, қосалқы өркендер жанама және қыстырмалы меристемалардан да дамуы мүмкін. Тегі меристема болғандықтан мұндай регенерант өсімдіктері бастапқы өсімдіктермен генетикалық біркелкі болады.
Регенерант өсімдіктердің каллустан пайда болуы
Микрокөбейтудің тағы бір көп таралған әдісі ол өсімдіктерді каллустан органогенез немесе эмбриоидогенез жолымен шығару. М орфогенездің басталуы қоректік ортадағы фитогормондардың қатысымен реттеледі. Органогенез арқылы регенерант өсімдіктерді алудың модельді жүйесі ретінде темекі бола алады (20-сурет) . Қоректік ортада фитогормондардың концентрациясын өзгертіп отырып, каллустан немесе тікелей экспланттан өркендерді өсіруге болады. Мысалы, темекінің Nicotina tabacum мен N. Оссіdentalis деген екі түрін бірі мен бірін будандастырғанда, будан ұрықтар тұзілген. Бірақ бұл ұрықтар табиғи жағдайда өне алмаған. Оларды in vitro- да өндіріп каллус алынды. Одан кейін каллуста сабақ бүршіктері мен тамырлардың пайда болуына жағдай туғызылды. Сөйтіп темекінің бағалы буданы көбейтілді.
Өкінішке орай, Скуг пен Миллердің анықтаған заңдылығы (86-бет) ең маңызды ауыл шаруашылық дақылдарына - дәнді дақылдар мен бұршақ тұқымдастарды қамти алмайды. Демек, каллустың регенерациялық активтілігіне генотиптін тигізетін әсері
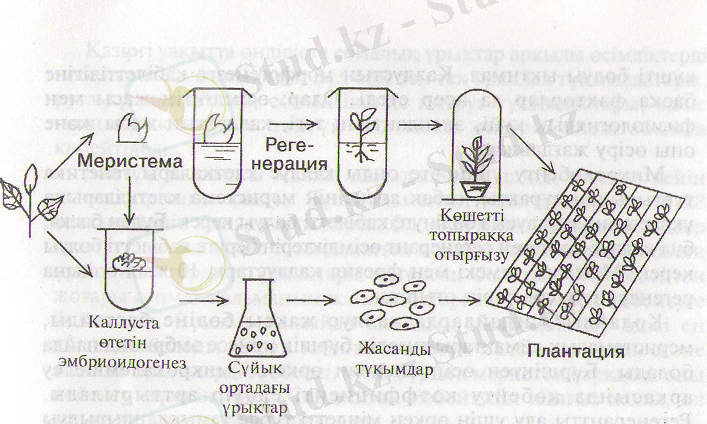
19-сурет. Клондық микрокөбейтудің технологиясы.
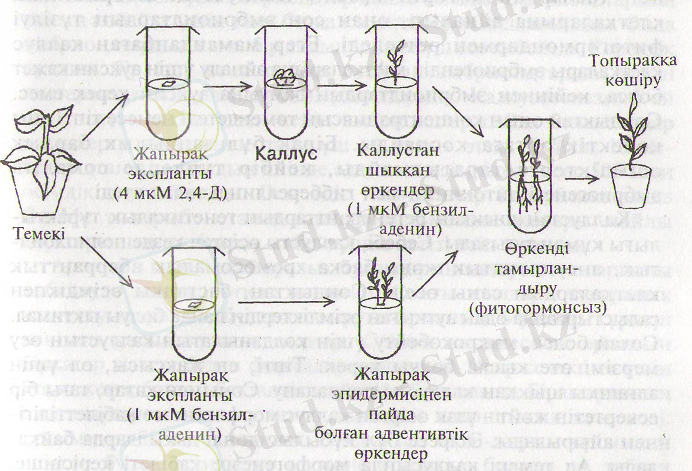
20-сурет. Темекі өсімдігінің регенерация жолдары (4) .
күшті болуы ықтимал. Каллустың морфогенезге қабілеттілігіне басқа факторлар да әсер етеді. Олар: өсімдіктін жасы мен физиологиялық күйі, экспланттың тегі, каллустың жасы және оны өсіру жағдайлары.
Микрокөбейту үшін өте оңды каллус клеткалары генетика тұрғысынан тұрақты, сабақ апексінің меристема клеткаларына ұқсас және тоқтаусыз бөлінуге қабілетті болуы керек. Бұдан басқа, бұл каллус көптеген регенерант өсімдіктерін беруге қабілетті болуы керек. Мысалы, темекі мен фрезия каллустары 10 жыл бойына регенеранттарды беріп отырған.
Қолайлы жағдайларда каллус жақсы бөліне бастайды, меристемалық аймактары тұзеліп бүршік немесе эмбриоид пайда болады. Бүршіктен өсіп шыққан өркенді микроқалемшелеу арқасында көбейту коэффициенті едәуір арттырылады. Регенерантты алу үшін өркен міндетті түрде тамырландырылуы қажет.
Ұрыққа ұқсас құрылымдар - эмбриоидтар - бір мезгілде сабақ пен тамыр апекстерін тұзеді. Каллустың эмбриондық клеткаларына айналып, онан соң эмбриоидтардың тұзілуі фитогормондармен реттеледі. Егер маманданбаған каллус клеткалары эмбриогендік клеткаларына айналу үшін ауксин қажет болса, кейіннен эмбриоидтардың дамуына ауксин керек емес. Соңдықтан оның концентрациясын төмендетеді немесе тіпті оны қоректік ортаға қоспайды. Бірақ бұл заңдылық барлық өсімдіктерге таратылмайды, кейбір түрлерде сомалық эмбриогенезі цитокинин мен гиббереллинді талап етеді.
Каллустан шыккан регенеранттардың генетикалық тұрақтылығы күмән туғызады. Себебі, каллусты өсірген кезде полиплоидтық, анеуплоидтық және басқа хромосомалық аберранттық клеткалардың саны өседі. Соңдықтан, бастапқы өсімдікпен салыстырғанда одан аутқыған өсімдіктердің пайда болуы ықтимал. Солай болса, микрокөбейту үшін қолданылатың каллустың өсу мерзімі өте қысқа болуы керек. Тіпті, ең жақсысы, ол үшін алғашқы шыққан каллусты пайдалану. Соңымен қатар, тағы бір ескертетін жәйт - ұзақ өсірген каллус морфогенезге қабілеттілігінен айырылады. Әсіресе бұл қүбылыс дәнді дакылдарда байқа-лады. Ал темекі каллусында морфогенезге қабілеті керісінше, бірнеше жылдап сақталады.
Қазіргі уакытта өндірісте сомалық ұрықтар арқылы өсімдіктерді көбейту аз қолданылады. Бұл әдіспен тек орхидея тұқымдастары, рута тұқымдастары, соңың ішінде цитрус тұқымдастары, және гвинеялық немесе зәйтүн пальмасы өндіріс масштабында көбейтіледі.
Зәйтүн пальмасы майлы дакылдардың ішінде соядан кейін екінші орынды алады. Бұл пальманың плантациялары 25-28 жылдан кейін жаңартылуы керек. Ол үшін миллиондаған өскіндер қажет. Пальма айқас тозаңданатың өсімдік болғандықтан, оның ұрығынан тараған ұрпақтары өзгергіш болады. Сол себептен жоғары өнімді палъманың жаңа сорттарын тұқыммен көбейтуге болмайды. Бұндай жағдайда клондық микрокөбейту әдісі ең қолайлы болады. Бірақ табиғатта зәйтүн пальмасы өркен және жанама өркендерді тұзбейді, яғни бүршік меристема арқылы оны көбейту мүмкін емес. Соңдықтан көбейтудің жалғыз жолы, ол каллустан регенеранттарды алу.
Каллустарды ағаштың ұшындағы ең жас жапырақтардан шығарады. Алғашқы каллусты қолайлы қоректік ортаға көшіру нәтижесінде оларда эмбриоидтар пайда болады. Эмбриоидтардың саның көбейту үшін тағы басқа қоректік орта қолданылады. Онда эмбриоидтардың саны бір айдың ішінде үш есе көбейеді. Қоректік ортаны бір ізді алмастырып отырып эмбриоидтардан өскін шығарып, кейін оларды тамырландырады. Каллусогенезге 3 ай уақыт жұмсалады, одан кейін эмбриоидтар пайда болып, олар өскіндерге жетілгенше тағы үш ай уақыт өтеді. Сөйтіп 10 эмбриоидтан бір жылдың ішінде алғашқы өсімдікке ұқсас 500 мың өсімдік алуға болады. Осы әдіс Кот-д'Ивуар Республикасында Ла-Ме тәжірибе станциясында пальманың жоғары майлы сорттарының өндірістік көбейтуінің негізі етіп алынған.
Арпа мен қара бидай аралық будандарын каллусогенез арқылы көбейтуде жақсы нәтиже алынған. Соңғы жылдары түраралық және туысаралық будандастыруға арпаның бірнеше түрлері кеніңен қатыстырылып жатыр. Соңың нәтижесінде арпаның бірқатар жақсы будан формалары алынған. Олар селекция үшін өте қажетті, бағалы бастапқы материал бола алады. Бірақ арпаның будандарымен айналысқанда мынадай қиыншылық кездеседі: тұқымның бітімі өте төмен, соңдықтан бірлі-жарым түқымнан өсірген будан өсімдіктер аз және ұрпақсыз болады да, табиғи
жолмен көбеймейді. Пісіп жетілмеген жас ұрықтарды in vitro жағдайында өсірсе, пайда болған каллустары жоғары тотипотенттік қасиет көрсетеді және оларды мерзімділікпен жаңа ортаға көшіріп отырса, регенерант өсімдіктерін шығаруға қабілетін бір жылға дейін сақтайды.
Осындай сомалық клеткалардан өсіп шыққан регенеранттардағы хромосомаларының саны зиготадан пайда болған ұрықтан шыққан будандармен бірдей болған. Бұл әрине микрокөбейтудің осы тәсілін қолданғанда, генетикалық тұрақтылықтың сақталатыңдығының белгісі. Сөйтіп, сомалық клеткалардан шыққан каллус ұлпаларын астық тұқымдастарды клондық көбейту үшін пайдалануға болады.
Соңғы жылдары астық тұқымдастардың бірнеше түрлерінде сомалық эмбриогенез процесінің өтуі жөніңде морфологиялық және гистологиялық дәлелдер алынған. Пісіп жетілмеген ұрықтарды, жас гүлшоғырларының және жапырақтардың төменгі жағындағы кесінділерін Мурасиге-Скугтың қоректік ортасында өсіргенде эмбриогендік каллус ұлпалары пайда болады. Осы эмбриогендік каллустарды ұзақ уақыт өсіруге болады және көптеген эмбриоид-тарды алып отыруға болады. Американ ғалымы И. Васил пікірінше, бұл әдіс астық тұқымдастарын микроклондап көбейту үшін өте қолайлы. Эмбриоидтардан тез уақытта көптеген өсімдік шығады, ол өсімдіктер бір клеткадан өсіп шығады, өйткені бір ғана эмбриогендік клетканың бөлінуі нәтижесінде пайда болады.
Бұдан басқа, эмбриоидтан шыққан регенеранттар әрдайым генетикалық тұрақтылықты көрсетеді. Клондық көбейтуде Бұлөте маңызды жағдай. Соңымен, редифференцияланбаған каллус клеткаларынан регенеранттардың пайда болатың екі жолы да (эмбриоидогенез бен органогенез) өте тиімді және кейбір жағдайларда олар микрокөбейтудің жалғыз ғана жолы болады.
Өсімдіктерді көбейту үшін эмбриоидтарды қолданатың тағы бір жол бар - ол жасанды тұқым. Жасанды тұқым деп полимер қабығымен қапталған эмбриоидтарды атайды. Бірақ практикада барлық эмбриоидтарды бірдей даму кезеніңе келтіру, яғни оларды синхрондау, өте қиын. Көлемі және тығыздығы бірдей эмбриоидтарды алу үшін оларды тесігі белгілі (400 - 500 мкм) илектен өткізеді немесе сахарозаның тығыздық градиентін әр түрлі етіп жасап центрифуга арқылы бөліп алады.
Қазіргі кезде сәбіздің, жоңышқаның, бидайдың, күріштін, балдыркөктін барлығы бірдей белгілі даму кезеңінде эмбриоидтардың жаппай шығарудың лабораториялық әдістері дайындалған. Оларды тұқым ретінде пайдалану үшін, әр түрлі қолайсыз жағдайлардан қорғайтың және ұзақ уақыт өміршендігін сақтауға себін тигізетін қабыкшаға салу керек. Қабыкша ретінде натрий альгинат қолданылады. Соңымен қатар, бұл қабық суды ұстап тұруы керек және капсуланың ішінде асептикалық жағдайды қамтамасыз ету қажет. Мақсатқа сәйкес капсуланың ішіне сомалық эмбриоидтың алғашқы дамуына қажет қоректік заттарды салуға болады мысалы, көмірсулар мен минералдық тұздарды. Әзірше бұндай «тұқымды» тек лабораторияларда шамалы мөлшерде ғылыми-зерттеу және селекциялық жұмыстары үшін алады. Ал биотехнологияның мақсаты оларды өндірісте мол мөлшерде шығару.
Өсімдіктер физиологиясы, генетикасы және биоинженериясы Институтыңың клеткалық инженерия лабораториясында Е. С. Сванбаев қызметтестерімен жоңышқаның жасанды тұқымын алудың технологиясын зерттеп дайындаған (21-сурет) . Жоңышқаның залалсыздандырылған өскіндерінің жапырағының сағағын Гамборгтың агарлы қоректік ортасында өсіріп, каллус алынады. Одан кейін каллус Гамборггың сұйық ортасына көшіріледі. Соңан соң суспензияда сомалық эмбриогенез басталады. Бір апта өткен соң суспензияны тесіктері әркелкі (200-300-400-500 мкм) торлар арқылы өткізіп, көлемі бірдей, яғни даму кезеңі біркелкі, эмбриоидтар іріктеліп алынады. Алынған эмбриоидтар гормондары жоқ қоректік ортаға көшіріліп, дамуы жалғасады. Ол үшін қоректік ортаға абсциз қышқылын енгізу керек, соңда эмбриоидтардың өсуі тежеледі, бірақ даму процесі тоқтамайды. Эмбриоидтардың алғашқы (тұқымжарнақ) жапырақтары өскен соң, оларды эксикаторда бөлменің температурасында кептіріп, қабық тұзілуі үшін полимермен өндейді. Полимерді ҚазМУ-дың химия факультетінің ғылыми қызметкерлері синтездеген.
Клондық микрокөбейту әдістерін сарапқа салғанда, мынаны атап өту керек: меристемадан шыққан регенерант өсімдіктері бастапқы өсімдіктерімен генетикалық жағынан біркелкі болғандықтан, каллус немесе экспланттың маманданған клеткаларынан
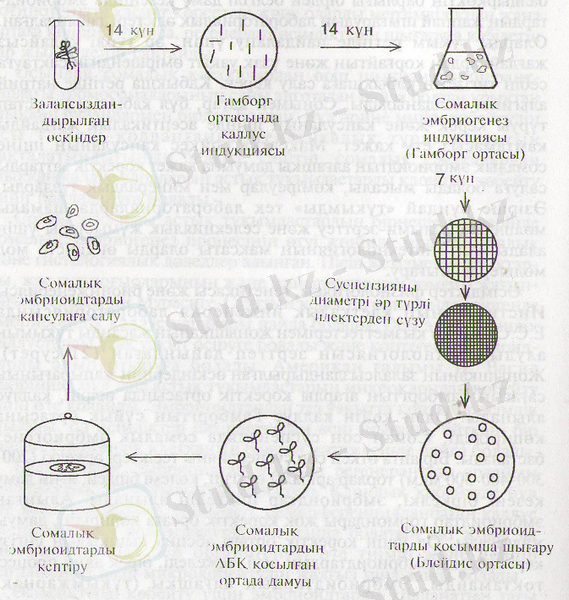
21-сурет. Жонышканың "жасанды ұрықтарын" шығару схемасы.
пайда болған өсімдіктерден едәуір айырмашылықтары болады. Шыққан өсімдіктерде біршама геномдық мутациялар болуы мүмкін.
Каллустан пайда болған өркендерде мутациялары бастапқы кезде көрінбейді, себебі олар жапырақтың формасына, гүлдердің түсіне, жемістін сапасына байланысты болуы мүмкін. Соңдықтан олар кейінірек өсімдік пісіп жетілген кезде ғана байқалады. Мысалы, қолтық меристемасынан пайда болған аспарагустың регенеранттарында генетикалық ауытқулық байқалмаған, ал каллустан шыққан өсімдіктердің 70% полиплоидтық болған. Егерде өсімдікті каллус арқылы көбейту қажет болса, ол үшін тек жас мүшелерінен алынған каллусты пайдалану керек.
Соңымен, генетикалық жағынан біркелкі өскіндерді шығарудың ең сенімді жолы, оларды қолтық өркендерден алу. Сабақтың ұшындағы бүршікті (апексті) in vitro өсіргенде жалғыз өркен тұзіледі, мұнымен бірге оның апикальдық басымдылығы болады. Қолтық бүршігінен шыққан өркеннен өсімдік шығару өте ұзақ уақытты және көп енбекті керек қылады, себебі әрдайым өскен өркендерді бөліп алып, оларды тамырландыру керек. Жылдам көбейту каллустағы эмбриоидогенез арқылы өтеді. Цитрустар мен зәйтун пальмасын көбейту жұмыстары көрсеткендей, бұнда генетикалық тұрақтылық сақталады. Эмбриоидтарды пайдалану қолайлы, соңдықтан олардан жасанды тұқым шығару технологиясының болашағы зор.
3. 3. Микрокөбейту процесінің кезендері
Клондық микрокөбейту жүмысы 4 этаппен өтеді (22-сурет) .
I ЭТАП. Эксплантты in vitro жағдайында өсіру. Бұл кезде толығымен залалсызданған эксплантты алып, лайықты қоректік ортаға отырғызып, оның жақсы өсуіне қолайлы жағдай туғызу керек. Бұл этаптың нәтижелілігі эксплантты дұрыс таңдап алуға байланысты. Ол үшін донор өсімдіктін әр даму кезеңінде ұлпаларынан немесе мүшелерінен экспланттар алынып, олардың in vitro жағдайында өсу және морфогенездік қабілеттері зерттеледі.
II ЭТАП. Микрокөбейту. Өркендердің пайда болуы және олардың саның көбейту.
III ЭТАП. Өркендерді тамырландыру және оларды сақтау. Жақсы тамыр жүйесі пайда болуына және дамуына жағдай туғызу. Ол үшін қоректік ортаға ризогендік факторы, яғни ауксин қосылады. Тамырлар өсіп жетілген соң өсімдіктерді топыраққа көшіруге дайындайды немесе салқын жерге қояды. Төмен температура өсімдіктердің дамуын тежеп, ұзақ уақыт сақтауға мүмкіншілік береді.
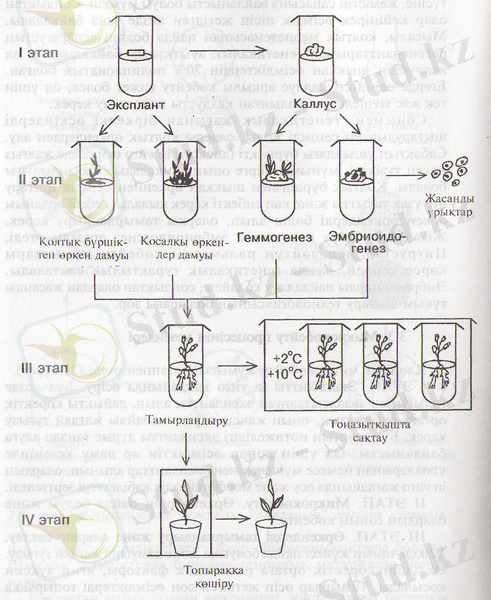
22-сурет. Клондық микрокөбейтудің кезеңцері (16) .
IV ЭТАП. Өсімдіктерді топыраққа отырғызу. Өсімдіктерді топыраққа отырғызу алдыңда оларды арнайы дайындайды. Сол үшін ауаның ылғалдылығын және жарықтың қарқындылығын арттырады. Өсімдіктер гетеротрофтық қоректенуден антотрофтық қоректенуге өтеді. Бұл ұқыптылықты талап ететін ең маңызды этап. Осы этапта өте сақ болу керек, себебі өсімдіктердің көбі бейімделе алмай, шығын болуы мүмкін.
Бірқатар өсімдіктерде технологияның I этапындағы негізгі қиындығы клеткалардың өздері ортаға бөліп шығаратың улы заттардың экспланттың өсуін тежеуі. Эксплантты антиоксидант-пен (мысалы, аскорбин қышқылы) жуып тастау, тіпті оны қоректік ортаға қосу да қажет. Кейбір ағаш өсімдіктерді көбейту технологиясында қиыншылықтар III этапқа байланысты. Онда Мурасиге-Скугтың қоректік ортасының негізгі құрамын өзгертеді (тұздардың мөлшерін екі есе азайтады немесе Уайттың ортасына ауыстырады), сахарозаның мөлшерін 0, 5-1 %-ке дейін төмендетеді, цитокининдерді шығарып тастайды, ауксиндерді қосады. Кеійде тамырдың тұзілуін ортаға қосқан хлороген және ферула қышқылдары қоздырады.
3. 4. Өсімдіктердің клондық микрокөбейуіне әсер ететін факторлар
Іn vitro жағдайында регенерацияның жылдамдығы өсімдіктер түріне байланысты болады, яғни генотиптін әсері күшті. Мысалы, қосжарнақты шөптекті өсімдіктер даражарнақты және ағаш өсімдіктерге қарағанда регенерацияға қабілеті едәуір жоғары. Өсімдіктік әр түріне ең тиімді регенерация әдісі іріктеліп алынады. Эксплант бөлініп алынатың өсімдіктер аурудан сау болуы керек. Теплицада өсіретін болса, жапырақтарына су шашыратпай өлшеулі суару керек, ауаның ылғалдығы төмен болу Қажет. Ең қолайлысы асептикалық ыдыстарда өсірілген залалсыздандырылған өскіндер.
... жалғасыСіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz