Бұлшықеттің жиырылу физиологиясы және механизмі

БҰЛШЫҚЕТТЕРДІҢ ЖИЫРЫЛУЫ
Адам мен омыртқалы жануарларда кездесетін ет тіні құрылымына қарай көлденең жолақты қаңқа еті, көлденең жолақты жүрек еті және бірыңғай салалы ет болып үшке бөлінеді. Бұл тарауда қаңқа еті мен бірыңғай салалы бұлшықеттің қасиеттері қаралады.
Көлденең жолақты еттер қатарына қаңқа еттері, тіл, көз, жұтқыншақ, көмей, өңештің жоғарғы бөлігіндегі еттер жатады. Қаңқа еттерінің қызметі ми қыртысымен тығыз байланысты, сондықтан олар адам денесінде ерікті қимыл-әрекеттерді қамтамасыз етеді.
Бірыңғай салалы бұлшықеттің де адам (жануар) үшін маңызы өте зор. Ол теріде, ішкі ағзалардың, сөл мен несеп түтіктерінің, қан және лимфа тамырларының, өкпе, кеңірдек, жыныс ағзаларының қабырғаларында орналасқан. Бұл ет аталған қуысты ағзалар мен түтіктерді кеңейтіп тарылтады да бүкіл ішкі ағзалар қызметіне әсер етеді.
Бұлшықеттерінің физиологиялық қасиеттері
Бұлшық ет қозғыш тіндердің бірі, демек, басқа қозғыш тіндерге тән қасиеттер - козғыштық, қозуды өткізу бұған да тән. Мұнымен бірге бұлшық еттің ерекше қасиеті - жиырылу.
Ет тіннің қозғыштық, дәрежесі қозу табалдырығы арқылы анық-талады. Көлденең жолақты қаңқа еті жүрек етіне қарағанда қозғыштау келеді, яғни оның қозу табалдырығы әлдеқайда төмен болады. Бірыңғай салалы бұлшықеттің қозғыштық дәрежесі жүрек етінікінен де төмен, демек, ол бұлшыкеттің тітіркендіру табалдырығы бәріненде жоғары болады.
Ет талшықтарының қозуды өткізу шапшандығы сомалық жүйке талшықтарынікінен көп төмен. Қозу ет талшығы сарколеммасын бойлай әртүрлі жылдамдыкпен, атап айтқанда: қаңқа етінде секундіне 3, 5-14 м, жүрек етінде 0, 9-1 м, ал бірыңғай салалы етте 0, 5 мм-ден 5-10 см-дей жылдамдықпен тарайды.
Ет тіні созылғыш (серпімді) келеді. Оның созылғыштығы резеңкеге қарағанда шүбәсіз, яғни босатқанда ет талшығының ұзындығы тура бастапқы созбай тұрғандағы қалпына келеді.
Бұлшықеттің негізгі қызметі және физиологиялық қасиеті - жиырылып жазылу. Жиырылу белгілі бір тітіркендіргіш әсеріне берілетін жауап, яғни ет ұзындығының қысқарып қатаюы (тонусының жоғарылауы) .
Жүрек еті мен кейбір бірынғай салалы, яғни ішек, лимфа тамыры қабырғаларындағы еттердің автоматиялық қасиеті де бар. Қаңқа етінің мұндай қасиеті жоқ. Ол орталық жүйке жүйесінің әсерінсіз жиырылмайды. Қаңқа еті тұлғалық жүйке жүйесімен, оның ішінде ми қыртысымен байланысты болғандықтан олардың жиырылуы адамның еркіне байланысты. Жүрек еті мен бірыңғай салалы етгі адам (жануар) өз еркімен жиырылта алмайды, бұлардың жиырылып жазылуын және автоматиялық қасиетін вегетативтік жүйке жүйесі реттейді. Ет пен жүйке байланысы бұзылса немесе түрлі себептермен ет ұзақ уақыт жиырылмаса, ет талшықтары құрылымы өзгеріп, семіп қалады (атрофия) . Ал керісінше етті арнайы әдістермен әдейі шынықтырса, жеке ет талшықтары үлкейіп, бұлшыкеттің көлемі ұлғаяды (гипертрофия) болады.
Тәжірибе жүзінде етті тікелей өзіне не жүйке арқылы әсер ете отырып қоздыруға болады. Бүлшықеттің жиырылуы оның сырттан келіп түскен тітіркеніске жауабы. Бір эфференттік жүйке тармақталып көптеген ет талшықтарымен түйісіп соншама мионевралдық түйіспе құрады. Бұл бұлшықеттің қозғалтқыш бірлігі (моторлық бірлік) боп саналады. Қозғалтқыш бірлігіндегі ет талшықтарының жалпы саны 3-6-дан 2000-ға дейін жетеді. Мысалы, саусақ бүгетін бұлшықеттің қозғалтқыш бірлігінде 10-25, дене мен қол етінде 500, балтыр етінде 2000 ет талшығы болады. Қозғалтқыш бірлігіндегі ет талшықтары жүйке талшығынан келген серпініске түгелдей бір мезгілде жиырылады. Жылықандыларда тез және баяу жиырылатын моторлық бірліктер болады. Тез жиырылатын еттер ақ еттерде, ал баяу жиырылатындар қызыл еттерде кездеседі.
Бұлшықеттердің жиырылу түрлері
Бұлшықет жиырылған сәтте ет қысқарып, осыған орай, оның тонусы (ширығуы) күшейсе, мұны ауксотониялық жиырылу деп, қысқарғанмен тонусы өзгермесе, оны изотониялық жиырылу деп атайды. Ал ет қысқармастан тек қатаятын болса (тонусы жоғарыласа), оны изометриялық жиырылу дейді. Белгілі бір тітіркендіргіш жалғыз әсер етсе, оған ет бір рет жиырылып жазылады. Мұны жеке жиырылу деп атайды. Ал етгі жиі-жиі тітіркендірсе, сандаған жиырылу бірігіп жинақталады да, ет біраз уақыт сіресіп қалады (тетанус) . Осыған орай ет жиырылуы жеке және сіресіп жиырылуы (тетанус) болып бөлінеді.
Көлбақаның балтыр етінің жеке жиырылуы 0, 11-0, 12 секундқа со-зылады, ол жасырын (латенттік) жиырылу (0, 01 сек) анық жиырылу (қысқару - 0, 05 сек), босаңсу (0, 05-0, 06 сек) кезендерден тұрады. Тітіркендіргіш әсер етісімен ет бірден жиьфылмайды, әсер тигеннен бастап қысғсірғанша біраз уақыт өтеді. Ол уақыт жасырын кезен деп аталады. Жасырын кезең тітіркендіргіш әсерінен жиырылудың баста-лу арасындағы уақыт. Жеке жиырылу сызығында бүл кезең көлденең көлбеу сызық ретінде байқалады (13-суретД) . Ет талшығы қысқарып қатайған сәтте сызық жоғары қарай көтеріледі де, босаңсып солған кезде керісінше төмендейді.
Қаңқа етінің әрекет потенциалы, сондай-ақ қозғыштық қабілетінің абсолюттік және салыстырмалы рефрактерлік кезеңдері өте қысқа болады. Сондықтан ет жиырылған сәтте оны одан әрі жиі-жиі тітіркендірсе оларға жауап бере береді. Бірнеше жиырылу бірігіп жинақталады да жиырылу серпіні күшейеді (13-сурет, Б) . Мұндай жинақталу толық, шала болып екіге бөлінеді. Жиі тітіркендірген сәтте әрбір тітіркеніс етке жиырылған кезде тисе, ет босаңсымастан одан әрі қарай жиырылып, оның жиырылу дәрежесі күшейе түседі. Бұл - толық жинақталу. Келесі тітіркендіргіш ет босаңси бастаған кезде тисе, ет солған жерінен бастап қайта жиырьшып бұрынғыдан да күшті жиырылады. Мұны жиырылудың шала жинақталуы не өркеш тәрізді жи-нақталу дейді. Демек, жиі-жиі тітіркендірсе, ет ұзақ және күштірек жиырылады, яғни сіреседі. Мұны тетанус деп атайды (13-сурет, В) .
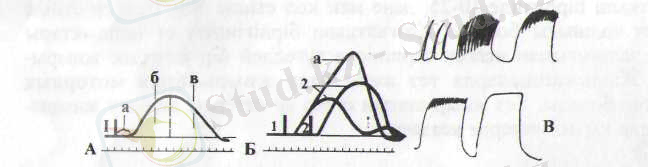
13-сурет.
Еттің жеке жиырылуы (А), жинақы жиырылуы (Б), тетанус (В)
- - тітіркендіргіштің бірінші әсері;
- - екінші әсері.
Тітіркендіру жиілігіне қарай тегіс тетанус және тісті (кертіш) тетанус болып бөлінеді. Тетанустың түрі тітіркендіргіштің жиілігіне байланысты. Тітіркендіру жиілігі секундіне 20-дан асса, жайпақ тетанус, ал 15-20 аралығында болса, тісті тетанус пайда болады.
Организмде жүрек етінен басқа еттер жеке жиырылмайды. Мұның себебі орталық жүйке жүйесі шеттегі ағзаларға әр қашан да көптеген
бірінен соң бірі тізбектелген серпіністер жібереді, жеке серпіністер болмайды. Бұл орталық жүйке жүйесінің ерекше бір қасиеті. Жүрек етінің жеке жиырылып босауы, оның рефрактерлік кезеңінің ұзақтығына байланысты. Орталық серпіністері жүрек етінің рефрактерлік кезеңіне тап болса, көбі жауапсыз қалады. Сондықтан бір серпініске жауап берген жүрек еті жиырылып онан соң босап үлгіреді.
Тез және баяу жиырылатын ет талшықтары болады. Тез жиырылатын талшық көбіне үлкен мотонейронның аксонымен бір түйіспе құрады. Мұнда жүйке серпінісінен әрекет потенциалы пайда болады да ол ет мембранасы бойымен тез таралып фазалық (кездік) қимылдар жасауға мүмкіндік береді. Ал баяу жиырылатын ет талшығы бірсыпыра мионевральдық түйіспелер құрады. Бұларда постсинапстық коздыру потенциалы әрекет потенциалын тудырмайды (олардың мембраналарында № арналары болмайды), сондықтан бұлшыкеттің тек қатаю қабілеті күшейеді, бұл тонустық жиырылу болып саналады. Демек, еттердің фазалық және тонустық жиырылу түрлері болады.
Бұлшыкеттің жиырылу және босаңсу механизмі
Көлденең жолақты бұлшықеттің жиырылу сырын білу үшін ет жиырылған кезде миофибрилдердің неліктен қысқаратының, ал солған кезде не себептен ұзарып, қалпына келетінін, сондай-ақ бұлшықеттің қозуы мен қысқаруы арасында қандай тәуелділік бар екенін білу керек. Ол үшін алдымен миоциттің, әсіресе миофибрилдердің құрылымына тоқтай кетейік. Қаяқа еті көлденең жолақты миоциттерден тұрады. Талшық үзын цилиндр тәрізді. Оның ұзындығы 10 смдей, кейде одан да көп, ені 12-70 мкм. Миоцитті қаптаған қабық саркөлемманың саркоплазмаға қарай шыққан көлденең Т - өсінділері болады. Олар миофибрилді саркомераға бөлетін 2 мембраналарының тұсында пайда болады. Сарколемманың іш жағында, миофибрилдер тобының сыртында олармен қатар жатқан саркоплазмалық ретикулум мембрананың Т - өсіндісіне жеткенде кеңіп Са 2 " иондарына толы қуыс (цистерна) құрады. Осы сарколеммалық бір көлденең Т - өсіндісімен оның екі жағындағы екі ретикулум цистернасының түйіскен жері (жасуша ішіндегі үш элементтен тұратын триада немесе үштік) - ішкі түйіспе деп аталады.
Бұлшықеттің жиырылып қысқаруы мен қайта босауы саркоплазмадағы миофибрилдердің ұзындығының өзгеруіне байланысты. Олар талшықтың бір ұшынан екінші ұшына дейін созылатын қатар-қатар орналасқан жіпшелер будасы (14-сурет, а, б) . Миофибрилдерді әрбір 2, 5 мкм-ден соң 2 мембрана көлденең 2000-дай бөлікке - саркомераға бөледі. Әр саркомераның қақ ортасында 2500-дей жуандығы 10 нм миозин және 2 мембрананың екі жағына тіркелген миозиннен екі есе жіңішке - диаметрі 5 нм актин орналасады. Актиннің ұшы екі жақтағы миозин (белок) талшықтары арасына жартылай кіріп тұрады. Миозин жіпшелерідің күңгірт (А) бөлігінде, ал актин ашық (I) бөлігінде болады. Бұл бөліктер миофибрил бойында бірінен соң бірі кезекпен орналасқан. Қатар-қатар жатқан миофибрилдердің күңгірт бөліктері бірыңғай бір қатарда, ашық бөліктері екінші қатарда орналасқандықтан, микроскоппен қарағанда жолақ-жолақ болып көрінеді.
Миозин шиыршықты 150 молекулалардан тұрады. Әр жіпшенің ұшында екі жұмыр (глобула) басы болады. Осы домалақ өсінділері арқылы (көлденең көпірше) миозин актинге жабысады. Миозиннің осы өсіндісінің АТФ қышқьшын ыдырататын ферменттік қасиеті бар. Бұл қасиеті актинмен байланысқанда 10 есе жоғарылайды.
Актин шиыршықты жіпшелерден тұрады, олардың ішінде әрбір 40 нм аралығында домалақ басы бар тропонин молекуласы және тропомиозин белогі кездеседі. Тропомиозин жіпшелері жиырьшмаған етте актиннің миозин өсінділері жабысатын жерін жауып, миозиннің актинге жабысуына кедергі жасайды.
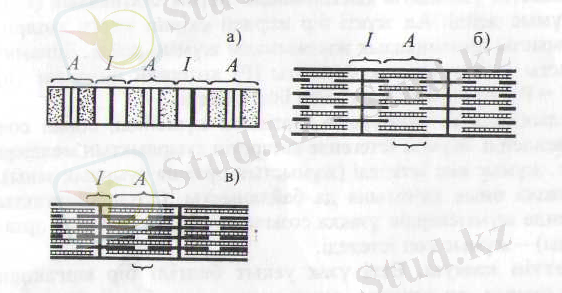
14- сурет. Миофибриллдердің құрылымы.
А - дискілері (маңғалдары) мен I және Н (а) белдеулері көрсетілген. Босансыған (б) және жиырылған (в) миофибрилдегі жуан (миозин), жінішке (актин) жіпшелері.
Ет жиырылған кезде талшықтар неліктен қысқаратынын түсіндіру үшін қазіргі З. А. Хаскельдің "белок талшықтарының жылжуы" теориясы айрықша қолдау тауып отыр. Бұл теория бойынша ет жиырылған сәтте актин талшықтары миозин бойымен сырғып толығынан олардың ара-арасына кіреді. Мұның салдарынан миофибрилдердің ашық бөлімі (I) қысқарады, тіпті жоғалып та кетеді, ал күңгірт бөлімі (А) оның қақ ортасындағы тек миозин жіпшелерінен тұратын ашыңқы (Н) тілімі жоғалып, одан әрі күңгірттене түседі және күңгірт (А) бөлімдері бір-біріне жақындайды (14-сурет, в) . Осы сәтте мнофибрилді жай микроскоппен қараса, оның көлденең жолақтығынан айырылғанын көруге болады. Актин және миозин жіпшелері ет жиырыл-ғанда қысқармайды. Актин жіпшелері миозин жіишелерінің арасына енгендіктен саркомера қысқарады. Бірнеше саркомераның қысқаруы миофибрилді әжептәуір қысқартады.
Актиннің миозиндер арасына кіруі миозиндердің көлденең өсінділері, актомиозиннің ферменттік қасиетінің жоғарылауы және саркоплазмада Са 2+ -иондары деңгейінің көтерілуімен байланыстырылады.
1939 жылы В. А. Энгальгард пен М. Н. Любимова ет жиырылған кезде миозиннің ферменттік белсенділігі өсетінін және ол еттегі энергия көзі АТФ қышқылын ыдырататынын анықтаған. Біраздан соң венгер биохимигі
Д. Сцент-Дюорди етте актиннің болатынын және актин мен миозин әрекеттесуінен актомиозин пайда болып, оның ферменттік қасиеті миозиннен 10 есе жоғары болатыньш дәлелдеді. Соңғы кезде бұл екі белоктың әрекеттесуі үшін саркоплазмада Са 2+ иондары мөлшері белгілі бір деңгейге жетуі (2-15х10 6 ) және иондарының болуы қажет екенін анықтаған. Мұнымен қатар актин оралымдары арасындағы тропонин мен тропомиозин молекулаларының керектігі де дәлелденді. Осы мәліметтерді пайдалана отырып, "белок талшықтарының жылжуы" теориясының мынадай моделі (үлгісі) қолданылады.
Ет талшығы жиырылуы үшін мотонейроннан мионевральдық түйіспеге импульс келіп жетуінен әрекет потенциалы туып, сарколеммада теріс заряд пайда болады. Ол мембрана бойымен Т өсінділері арқылы ішкі түйіспеге жетіп, ретикулум цистернасының Са 2+ ионына өтімділігін жоғарылатады. Мұның салдарынан кальций иондары цистернадан саркоплазмаға өтеді. Калыдий иондары тропонин молекулаларымен әрекеттесіп тропонин - тропомиозин комплексін құрады. Бұл тропониннің пішінін өзгертеді де, актин оралымдарының арасына тропомиозиннің тереңірек кіруіне жағдай жасайды. Сөйтіп актин тропомиозин кедергісінен құтылады, актинге миозиннің көлденең өсінділерінің жабысатын жері босайды. Бұл өсінділер актинге жабысып ондағы миозиннің ферменттік қасиетін жоғарылатады. Бұл үшін иондарының жеткілікті болуы да шарт. АТФ кышқылы ыдырай бастайды. Фосфор кышқылы бөлініп, аденозинді фосфор қышқылы пайда болады. Осыған орай миозин өсіндісінің (басының) пішіндік өзгеруіне байланысты олар қайық ескектерінің қимылын жасап (еңкейіп, жазьшып) актин жіпшелерін миозин жіпшелерінің ортасьша қарай тартады (енгізеді) . Бұған босаған энергия жұмсалады. Мұнымен қатар миозин өсіндісінде АТФ қышқылы қайта түзіледі және Са 2+ иондары саркоплазмада азая бастайды. Осыған орай актинмен миозин байланысы бұзылады. Әрекет потенциаларының реполяризациясы кезінде саркоплазма ретикулумының мембранасындағы кальцийдің тартқыштық белсенділігі күшейіп, Са 2+ иондары саркоплазмадан кері цистернаға өтеді, артығы кейде миоциттен сыртқа да шығарылады. Тропонин кешені (комплексі) ыдырап, тропомиозин алғашқы орнына келіп актиннің активті жерлерін жабады. Келесі серпіністе (ӘП-да) осы механикалық және химиялык үрдістер қайтадан қайталанады. Сонымен бұлшықеттің жиырылуы аяқталарда протоплазмадағы кальций иондары азаяды. Кальций мөлшері бастапқы калпына келерде жиырылған ет босайды да актин кері жылжиды, ет талшықтары ұзарып, бұрынғы қалпына келеді, осыған байланысты АТФ қышқылының мөлшері де бұрынғы қалпына келуге тиіс. Энергия өзегі бастапқы деңгейіне жетпесе ет толығынан босап шықпайды. Онда контрактура (құрысу) пайда болуы мүмкін.
Бұлшықеттің жиырылу күші
Бұлшыкеттің жиырылу күші тітіркендіргіштің күшіне, жиілігіне және бұлшықеттің құрылыс ерекшелігіне, функциялык жағдайына бай-ланысты. Бұлшықет мындаған ет талшыктары будағынан (топтары-нан) тұрады.
... жалғасыСіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz