Генетика және селекция негіздері: тұқым қуалаушылықтың заңдылықтары, дамуы және моногибридті будандастырудың цитологиялық негіздері

ГЕНЕТИКА ЖӘНЕ СЕЛЕКЦИЯ НЕГІЗДЕРІ ЛЕКЦИЯЛАР
- Тұқым қуалаушылық пен өзгергіштіктің негізгі заңдылықтары
- Генетиканың даму тарихы
- Моногибридті будандастырудағы тұқым қуалау заңдылықтары
- Моногибридті будандастырудың цитологиялық негіздері
- Дигибридті будандастыру және белгілердің тәуелсіз ажырау заңдылығы
- Дигибридті будандастырудың цитологиялық негіздері
- Тұқым қуалаушылықтың хромосомалық теориясы
- Цитоплазмалық тұқым қуалау
- Генетикалық карталар
- Гендердің өзара әрекеттесуі
- Адам генетикасы
- Медициналық генетика және кейбір тұқым қуалайтын аурулардың алдын алу мен емдеу
Тұқым қуалаушылық пен өзгергіштіктің негізгі заңдылықтары
Генетиканың даму тарихы
Генетика - бүкіл тірі организмдерге тән қуалаушылық пенөзгергіштіктің зерттейтін биология ғылымының бір саласы. Тұқым қуалаушылық пен өзгергіштіктің заңдылықтарын ашып, оларды қоғамды дамыту үшін пайдаланудың жолдарын шешуде генетика басқа салаларынынң арасында маңызды орнын алады.
Жер бетіедегі тірі материяның дамуы олардың үздіксіз ұрпақ алмастырумен қатар жүріп отырылады. Тіршілік органимдердің көбеюімен тікелей байланысты. Сол арқылы белгілі бір биологиялық түрге тән белгілер мен қасиеттер ұрпақтан-ұрпаққа беріліп отырылады. Басқаша айтқанда, ұрпақтар белгілі дәрежеде өзінің ата-ансына ұқсас болып туады. Мұны тұқым қуалаушылық дейді. Көп жағдайда берілетірндіктен, ұрпағы ата-аналарына ұқсас болып келеді. Бірақ олардың арасында толық ұқсастық болмайды. Бір ата-анадан тарайтын ұрпақтың бір-бірінен қандай да бір белгісі жөнінде айырмашылығы болады.
Организмнің тұқым қуалаушылық қасиеті сыртқы орта факторларының әсерінен үнемі өзгеріп отырады. Оны өзгергіштік дейді. Көбею барысында қаңтар екінші біреуі өзгеріске ұшырайды. Осыған байланысты олар жаңарып, түрлене түседі.
Тұқым қуалаушылық пен өзгергіштік - бірімен-бірі қатар жүретін, бір жағынан, бір-біріне қарама-қарсы, өзара тығыз байланысты процестер.
Организмдердің тұқым қуалаушылығы мен өзгергіштігі туралы ғылымды генетика деп атайды (грекше «ganesis» тегі) . Бұл терминді 1906 жылы ағылшын бтологі У. Бэтсон ұсынады.
Тұқым қуалаушылық туралы алғашқы түсініктер ежелгі дәуірде Демокрит, Гипократ, Платон және Аристотель еңбектерінде кездеседі. Гиппократ: «Жұмыртқа жасушасы мен спермий организмнің барлық бөліктерінінің қатысуымен қалыптасады және ата-ананың бойындағы белгі-қасиеттері ұрпағына тікелей беріледі» деп есептеді. Ал Аристотельдің көзқарасы бойынша белгі-қасиеттердің тұқым қуалауы тікелей жүргізілмейді. Яғни, тұқым қуалайтын материал дененің барлық бөліктерінен келіп түспейді, керісінше, оның әр түрлі бөлшектерінің құрастырылуға арналған қоректік заттардан жасалады.
Бұдан кейін Ч. Дарвиннің пангенезис теориясы маңызды орын алады. Бұл теория бойынша өсімдіктер мен жануарлардың барлық жасушалары (клетка) өзінен ұқсас бөлшектер - геммулалар бөліп шығарады. Олар жыныс органдарына өтеді де сол арқылы белгілер мен қасиеттер ұрпаққа беріледі. Геммулалар кейде «мүлгіген жағдайда» болып, бірнеше ұрпақтан кейін білінуі мүмкін. Соған байланысты ұрпақтарда арғы ата-ана тектерінің белгі-қасиеттері қайталана алады деп есептелінген.
ХІХ ғасырдың 80-жылдарында «пангенезис» теориясын неміс зоологі А. Вейсман өткір сынға алады. А. Вейсман «ұрық плазмасы» туралы болжам ұсынады. Бұл болжамында тек жыныс жасушаларында кездесетін, тұқым қуалайтын заттың болатындығын айтты.
Генетиканың биология ғылымының жеке бір саласы ретінде қалыптасуына ХІХ ғасырдың екінші жартысында ашылған ірі ғылыми жаңалықтар себепші болды. 1865 жылы чех ғылымы Г. Мендельдің «Өсімдік будандарымен жүргізілген тәжірибелер» деген еңбегі жарық көрді. Ол тәжірибелері арқылы тұқым қуалушылықтың негізгі заңдылықтарын қалыптастырды. Сөйтіп, Мендель генетиканың негізін қалады. Бірақоның еңбегі 1865 жылдан бастап 35 жыл бойы көпшілік биологтерге, соның ішінде Ч. Дарвинге де белгісіз күйде қалды.
Г. Мендель ашқан тұқым қуалау заңдылықтары тек 1900 жылы ғана өзінің тиісті бағасын алды. Себебі үш елдің ғылымдары: голландиялық Г. Де Фриз, неміс ғалымы К. Корренс және австриялық генетик Э. Чермак-Зейзенгг әр түрлі объектілермен тәжірибелер жүргізіп, нәтижесінде, Мендель заңдарының дұрыстығын дәлелдеді. Көп кешікпей бұл заңдылықтардың жануарларға да тән екендігі анықталды. Оны 1902 жылы ағылшын биологі У. Бэтсон тауықтардың айдарларының пішіндерінің, ал Кюэно үй тышқандары жүндерінің ақ және сұр түстерінің тұқым қуалауы мысалында көрсетеді. Кейінірек 1909 жылы У. Бэтсон өсімдіктер мен жануарлардың әрқайсысының 100 шақты белгілерінің тұқым қуалауы Мендель заңдарына сәйкес жүретіндігін дәлелдейтін ғылыми деректерді жариялады. Сөйтіп, Мендель ілімі ғылымнан берік орын алды.
1909 жылы дат биологі В. Иогансен (1857-1927) биологияда аса маңызды болып есептелетін ген ( герекше “genos” - шығу тегі), генотип және фенотип деген ұғымдарды қалыптастырды.
Генетика тарихының бұл кезеңінде организмдердің жекелеген белгілерінің ұрпақтан-ұрпаққа берілуіне жауапты тұқым қуалаушылықтың материалдық бірлігі-ген туралы ұғым қалыптасып, Мендель ілімінің өзі қарай дамуына мүмкіндік туды. Дәл сол кездегі (1901 жыл) голландиялық ботаник Х. де Фриздің организмнің тұқым қуалацтын қасиеттерінің өзгеретіндігін көрсететін мутация теориясының ұысынылуы генетика ғылымының дамуында ерекше орын алады.
Генетика тарихындағы шешуші бір кезең америкалық генетик әрі эмбриолог Т. Морганның және оның ғылыми мектебінің тұқым қуалаушылықтың хромосомалық теориясын ашуымен тығыз байланысты. М. Морган және оның шәкірттері жеміс шыбыны-дрозофилаға тәжірибе жасаудың нәтижесінде тұқым қуалаушылықтың көптеген заңдылықтарын ашты.
Тұқым қуалайтын өзгергіштік туралы ілімді дамытуда орыс ғалымы Н. И. Вавилов зор үлес қосты. Ро 1920 жылы тұқым қуалайтын өзгергіштіктің ұқсас (гомологиялық) қатарлары заңын қалыптастырады. Бұл заң бір-біріне жақын туыстар мен түрлерде болатын тұқым қуалайтын өзгерістердің ұқсас болып келетіндігін дәлелдейді.
Ғалымға енгізілген жаңалықтың бірі - 1927 жылы орыс ғалымдары Г. А. Надсон мен Г. С. Филипповтің радиоктивті сәулелердің төменгі сатындағы саңырауқұлақтарда мутация алатындығын дәлелдеуі еді.
Ген теориясы дамытуда орыс биологтері А. С. Серебровский мен Н. П: Дубиннинің эксперименттік және тоериялық жұмыстарының үлкен маңызы болды. Сол сияқты популяциялық генетика мен эволюциялық генетиканың негізін қалауда орыс генетигі С. С. Четвериковтің алатын орны ерекше.
Генетиканың даму тарихы үш кезеңге бөлінеді. Оның алғашқы екеуі 1965-1958 жылдар аралығын, яғни классикалық генетика дәуірін қамтиды. Генетика тарихяндағы үшінші кезең 1953 жылдан басталды. Ол-химия, физика, математика, кибернетика сияқты нақты ғылымдардың зерттеу әдістері мен электрондық микроскоп, рентгенқұрылымдық анализ, т. б. қолданудың нәтижесінде молекулалық генетика негізінің қалауы.
1944 жылы америкалық микробиолог әрі генетик О. Эври тұқым қуалаушылықтың материалдық негізі - ДНҚ екендігін дәлелдеді. 1953 жылы америкалық биохимик әрі генетик Дж. Уотсон мен ағылшын биофизигі Ф. Крик ДНҚ-ның молекулалық құрылымының моделін жасады.
Қазіргі кездегі генетиканың дамуы тұқым қуалаушылық пен өзгергіштік туралы ілімнің барлық салаларында зерттеу жұмыстарының молекулалық деңгейде жүргізілетіндігімен ерекшеленеді. Мысалы, генді организмнен тыс қолдан синтездеу, дене жасушаларын будандастыру, генетикалық келуі (реперация), биополимерлерді қолдан синтездеу, ген инженериясы сияқты проблемаларды зеттеу кеңінен таралып отыр.
Генетика мен селекцияның дамуына Қазақстан ғылымдарының қосқан үлесі ерекше. Алшақ будандастыру, мутагенез, полиплоидия, гетерозис, т. б. мәселелерді қамтитын генетикалық зерттеулер тұрарарлық будандастырудың нәтижесінде бидайдың,, арпаның, будандары мен сорттарын алуда К. Мыңбаев, А. Ғаббасов, Ғ. Бияшев, Н. Б. Ахматуллина микроорганиздер генетикасының дамуына айтарлықтай үлес қосты.
Н. С. Бутарин, Ә. Е. Есенжолов, А. Ы. Жандеркин алшақ будандастыру әдісімен қойдың арқар-меринос тұқымын алды. М. А. Ермеков, Ә. Е. Еламанов, В. А. Бальмонт, т. б. қазақтың ақбас сиырынғ алатау сиырын және қостанай жылқысын, т. б. асыл тұқымдарды шығарды.
Қазақстанда тұңғыш рет М. Ә. Айтхожиннің басқаруымен молекулалық биология және ген инженериясы саласында көптеген зерттеулер жүргізіліп, алымға айтарлықтай жаңалықтар қосылды.
Қазақстан тұңғыш рет М. Ә. Айтхожиннің басқаруымен молекулалық биология және ген инженериясы саласында көптеген зерттеулер жүргізіліп, ғылымға айтарлықтай жаңалықтар қосылды.
Соңғы жылдары елімізге генетиканың аса маңызды салалары: молекулалық генетика, экологиялық генетика және радиациялық генетика бойынша ғылыми-зерттеу жұмыстары жүргізілуде.
Моногибридті будандастырудағы тұқым қуалау заңдылықтары
Гибридологиялық әдіс. Тұқым қуалаушылықтың заңдылықтарын зерттеудің ғылыми негізін Грегор Мендель қалады. Грегор Мендель 1822 жылы Гинчица шағын қыстағында, кедей шаруаның отбасында дүниеге келді. 1843 жылы Чехословакияның Брно қаласындағы Августин монастырында тыңдаушы болып орналасқан. Мендель оны бітіріп, діни атақ алғаннай кейін Вена университетіне түсіп, екі жыл жаратылыстану және математиканы оқып-үйренді. Оның бұл алған білімі бұршақ өсімдігіне жүргізген тәжірибелері нәтижесінде тұқым қуалау заңдылықтарын ашуға үлкен ықпалын тигізді. Ол 1856 жылы монастырьға қайтып келіп, өмірінің соңына дейін монах қызметін атқарады.
Г. Мендель Венаға оқып жүрген кезден-ақ өсімдіктерді будандастырумен шұғылданып, түрлі будандық ұрпақ типтерінің статистткалық арақатынасына назар аударып, есептеулер жүргізген.
Мендельдің бұл еңбектері 1856 жылы жазда басталған ғылыми-зерттеу жұмыстарына негіз болды. Ол өз тәжірибелеріне қолайлы объекті ретінде бұршақты (Pisum sativum) алды. Себебі, басқа өсімдіктермен салыстырғанда бұршақтың мынадай айрықша қасиеттері бар: 1) бірнеше белгілері бойынша бір-бірінен айқын ажыратылатын көптеген сорттары бар; 2) өсіруге қолайлы; 3) гүліндегі жыныс мүшелері күлте жапырақшаларымен толық қалқаланып тұратындықтан, өсімдік өздігінен тозанданады. Сондықтан әр сорт өзінше таза дамып-жетілетіндіктен, белгілері ұрпақтан-ұрпаққа өзгеріссіз беріледі; 4) бұл өсімдіктің сорттарын қолдан тозаңдандыру арқылы өсімтал будандар алуға болады.
Міне, сондықтан Мендель бұршақтың 34 сортынан белгілері айқын ажыратылатын 22 сортты таңдап алып, өз тәжірибелерін пайдаланды. Ол өсімдіктің негізгі жеті белгісіне көңіл аударды, сабағьның ұзындығы, тұкымының пішіні мен түсі, жетістіктерінің пішіні мен түсі, гүлдерінің түсі мен орналасуы. Мендель тәжірибелерін келесі бір ерекшелігі - белгілердің тұқым қуалауын зерттеуге гибридологиялық әдісті қолдану арқылы дәл және тиянақты не алуында болды. Бұл әдістің негізгі жағдайлары мыналар 1) будандастыру үшін бір-бірінен жұп белгілері бойынша айы ажыратылатын бір түрдің даралары алынады; 2) зерттелетін белгіге тұрақты, яғни бірнеше ұрпак бойы қайталанып отырады; 3) ұрпақтағы алынған будандарға жеке талдау жасалып, нақты сандық есептеулер жүргізілді.
Мендель өзінің тәжірибелерін 8 жыл бойы (1856-1864) Берн қаласындағы Августин монастырының бағында жүргізді. Ол зерттеулерінің нәтижесі туралы 1865 жылы 8 ақпанда сол Берн қаласындағы Табиғат зерттеушілер қоғамының кеңесінде Соның негізінде «Өсімдік будандарымен жүргізілген тәжірибелі атты еңбегін жариялады.
Қандай да болсын белгі-қасиеттерінде тұқым қуалайтын өзге кемшіліктері бар организмдерді будандастырғанда, «гибридті тор» алынады. Бір ғана жұп белгілерінде айырмашылығы бар ата-ана формалар будандастырылса моногибридті, екі жұп белгісі дигибридті, ал белгілердің саны көп болса полигибридті будандастыру деп атайды.
Моногибридті будандастыру. Тұқым қуалаушылықты; Мендель ең қарапайым моногибридті будандастырудан бастап, қарай біртіндеп күрделендіре түскен. Мендельдің ұсынысы «бок» гендер латын әріптерімен белгіленді. Бір аллельді жұптың гендері бірдей әріппен, яғни доминантты (басым) генді үлкен әріппен, ал рецессивті (басылыңқы) генді қіші әріппен белгілеу ұсынылған. Олай болса, доминантты қасиет көрсететін бұршақтың гүлінің қызыл түсі мен тұқымының сары түсін - А, рецессивті - гүлдің ақ тұқымының жасыл түсін - а деп, тұқымының доминантты тегіс пішінін - В, рецессивті кедір-бұдырлы әрібін - b деп белгілейміз. Ата-аналық формалар Р әріпімен белгіленеді (латынша «parent’s» ата-ана) . Аналықты 2 белгісімен, аталықты о* белгісімен, будандастыруды х, ұрпақтарын Ғ әріпімен (латынша «Ғilius» - ұрпақ) гаметаларды Г, ал бірінші, екінші және үшінші ұрпақтарын Ғ1, Ғ2, Ғ3 және т. б. деп белгілейді.
Бірінші ұрпақтың біркелкі болуы. Мендельдің бірінші заңы. . Аналары бір-бірінен бір жұп белгі бойынша ажыратылатьын даралар будандастыруды моногибридті деп атайды. Мендель өз тәжірибелерін
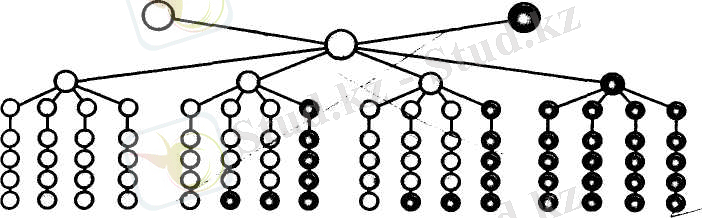
Моногибридті будандастыру барысы. Сары түсті дөңгелектер - доминантты белгісі бар, жасыл түсті дөңгелектер - рецессивті белгісі бар организмдер бірінде бұршақтың тұкымдары сары және жасыл түсті екі сортын алып будандастырған. Сонда бірінші ұрпақтан алынған будандардың барлығы сары тұқымды болып, жасыл түс көрінбеген (98-сурет) .
Дәл осындай нәтиже бұршактың қызыл гүлді және ақ гүлді формаларын алып будандастырғанда да көрініс берді. Яғни, қызыл гүлді және ақ гүлді бұршақты будандастырғанда, бірінші будандық ұрпақтың барлығы қызыл гүлді болып, ақ түс байқалмады. Осы зерттеулердің нәтижесінде, Мендельдің бірінші заңы - бірінші будандық ұрпақтың белгілерінің біркелкі болу заңы қалыптасты. Мұны бірінші ұрпақ будандарының біркелкілік заңы немесе толық доминанттылық заңы деп те атайды (98-сурет) .
Бірінші ұрпақта басымдық касиет көрсетіліп, бірден жарыққа шығатын белгіні доминантты, көрінбей қалған белгіні рецессивті деп атайды. Қарама-қарсы (альтернативті) белгілерді анықтайтын жұп гендер аллельді гендер деп аталады. Мысалы, тұқымның сары түсі мен жасыл түсін, гүлдің кызыл түсі мен ақ түсін анықтайтын гендерді аллелъді дейді. Сонымен зиготада әр уақытта ата-аналарынан қабылдаған екі аллель (ген) бар деп есептеп, кез келген белгі бойынша оның генотиптік формуласын екі әріппен белгілейді. Генотип деп ата-аналардан алынатын гендердің толық жиынтығын айтады. Генотипіне Қарай организм гомозиготалы не гетерозиготалы болуы мүмкін. Гомозиготалы деп тек бірыңғай доминантты (АА) немесе рецессивті (аа) аллельдерден тұратын организмді айтады. Гетерозиготалы о рганизм керісінше, әр түрлі аллельдерден тұрады (Аа) . Организмге тән ішкі және сыртқы белгілердің жиынтығын фенотип деп атайды. Мысалы, тұкымның түсі, пішіні, сабақтары, биіктігі, көздің қара немесе ақ болуы және т. б.
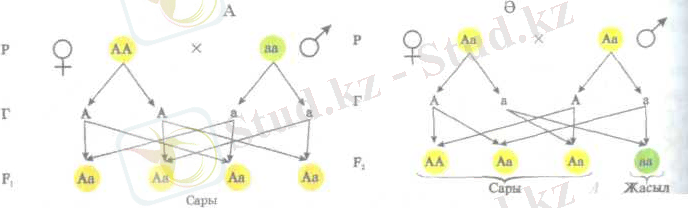
Меңдельдің жүргізген тәжірибелерін жоғарыда келтірілген белгілерді пайдалана отырып, сызбанұсқа түрінде былай көрсетуге болады.
бұршак тұқымының сары түсін - «А», жасыл түсін «а» деп
белгілейік (99-сурет, А) . Бірінші үрпақтағы (Ғ1) барлық даралары біркелкі генотипі бойынша гетерозиготалы, фенотипі бойынша бәрі
99-сурет. Моногибридті будандастыру барысының сызбанұсқасы
сары тұқымды болып шығады. Осыдан келіп біркелкілік ережесін былай тұжырымдайды:
Бір-бірінен айқын бір жұп белгі арқылы ажыратылатын гомозиготалы дараларды будандастырса, бірінші ұрпақтан генотипі де, фенотипі де біркелкі будандар алынады. Мынаны бірінші ұрпақ будандарының біркелкілік заңы деп атайды.
Белгілердің ажырау заңы. Мендель бұл тәжірибені одан әрі жалғастырып, бірінші ұрпақтағы будандарды өздігінен тозаңдан-дырғанда - екінші ұрпақта сары тұқымды да, жасыл тұқымды да даралар алынған. Екінші ұрпақта ата-аналарының екеуіне де тән белгілердің көрініс беру заңдылығын ажырау заңы деп атайды. Белгілердің ажырауы кездейсоқ болмай, белгілі бір сандық ара-қатынаста жүреді. Атап айтқанда, екінші ұрпақта алынған барлық өсімдіктердің ірісі сары тұқымды, ірісі жасыл тұқымды болып шығады.
Олай болса, фенотипі жағынан доминантты және рецессивті белгілердің сандық арақатынасы 3:1-ге тең болып өзгереді (99-сурет, Ә) .
Бұл сызбанұсқадан екінші ұрпақтағы даралардың бір бөлігі, яғни 25-і генотипі жағынан доминантты белгі бойынша гомозиготалы (АА), екі бөлігі - 50-і гетерозиготалы (Аа) және бір бөлігі - 25-і рецессивті белгі бойынша гомозиготалы (аа) екенін оңай түсінуге болады. Осы айтылғандарды қорыта келе, ажырау заңын былай тұжырымдайды:
Бірінші ұрпақтағы алынған гибридтерді өзара будандастырған жағдайда, екінші ұрпақта (Ғ 2 ) белгілердің ажырауы жүреді. Ажырау арақатынасы фенотипі бойынша З:1, ал генотипі бойынша 1:2:1 -ге тең болады, бірнеше арақатынастар жоғарыдағы сызбанұсқада бейнеленген.
Тақырыптың түйіні
1. Тұқым қуалаушылық зандылықтарын тұңғыш рет Чех ғалымы
Грегор Мендель ғылыми тұрғыда зерттеген.
2. Г. Мендель өз тәжірибелерінің қорытындысын 1865 жылы
«Өсімдік будандарымен жүргізілген тәжірибелер» деген еңбегінде
жариялаған.
3. Г. Мендель бұршаққа тәжірибе жасады. Ол тәжірибе жүргізуге өте қолайлы объекті болды. Себебі басқа өсімдіктермен салыстырғанда, бұршақтың мынадай ерекше қасиеттері бар: 1) бірнеше белгілері бойынша бір-бірінен айқын ажыратылатын көптеген сорттары бар; 2) өсіруге қолайлы, күй талғамайды; 3) гүліндегі жыныс мүшелері күлте жапыракшалармен толық қалқаланып тұратындықтан, өздігінен де, айқас та тозаңданады; 4) бұршақтың 34 сорты болған, соның 22 сортына тәжірибе жасады.
Г. Мендель бір белгісі бойынша тәжірибе жасау арқылы екі заңдылық ашты. Атап айтқанда, «бірінші ұрпақтың біркелкілігі» және «белгілердің ажырауы» заңдары.
Моногибридті будандастырудың цитологиялық негіздері
Гаметалар тазалығы ережесі. Бірінші буында алынатын будандардың біркелкі болуы мен екінші буын ұрпақтарында белгілердің ажырау құбылысын түсіндіру үшін, Мендель гамета тазалығы болжамын ұсынды. Оның мәні - организмнің кез келген белгі-қасиетінің дамуын тұқым қуалау факторы, яғни генді анықтайды. Мысалы, раушан гүлдің қызыл гүлділері мен ақ гүлділерін алып будандастырғанда, бірінші будандық ұрпақтың барлығы қызыл гүлді болған (101-сурет) . Ол бірінші будан ұрпақта қызыл гүлді өсімдіктің Доминантты «А» гені бар гаметасы мен ақ гүлдінің рецессивті «а» гені бар гаметаларының қосылуының нәтижесі болып есептеледі. Соңдықтан олардың генотипінде гүлдің қызыл түсін де, ақ түсін де анықтайтын гендер болады. Бірақ қызыл түстің гені доминантты
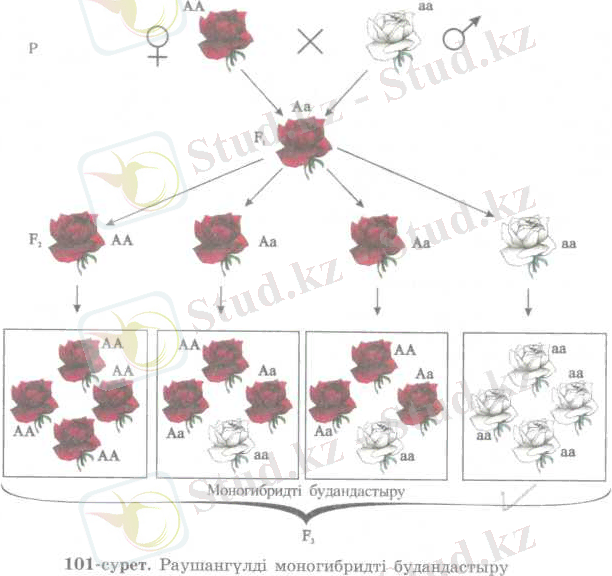
болғандықтан, бірінші ұқрпақтың барлығы да қызыл гүлді болады. Сонда олардың фенотипі бірдей болғанымен, генотипінде екі түрлі ген болғаны. Ал ондай будан организмнен гамета түзілгенде, оған тек бір ғана доминантты «А» гені немесе рецессивті «а» гені беріледі. Бұл жағдайда будан организмнің гаметасында аллельді (жұп) гендер бір-бірімен араласып кетпей, таза күйінде сақталады. Мұны гамета талшыгы дейді.
Әрине, Мендель будан организмнен гамета түзілу кезінде, геннін таралу процесін жасушаның нақты бір материалдық құрылымдарымен және жасушаның бөліну механизмімен байланыстыра алмады.
Алайда Г. Мендель хромосомалық теория қалыптаспай тұрып-ақ, мейоздық бөлінудің механизмі мен гендердің әрекеттері туралы күні бұрын дұрыс болжам жасап, гамета тазалығы туралы ережесін ұсынады.
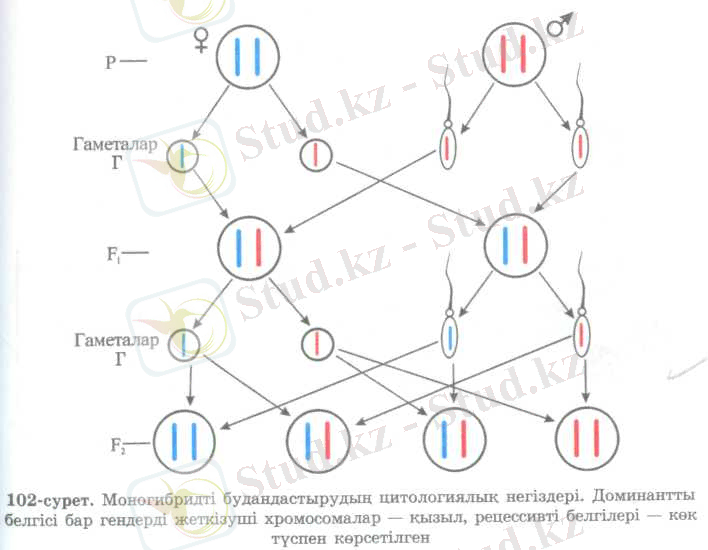
Моногибридті будандастырудың цитологиялық негіздері. Мендельдің әлемге әйгілі тұқым қуалау заңдылыктары 1865 жылы жарияланғанымен, сол кезде толық колдау таппай, 1900 жылға дейін белгісіз болып калды. Өз тәжірибелері негізінде дұрыс нәтиже алғанымен бірінші ұрпақтағы будандардың біркелкі болуы және екінші ұрпақта белгілердің ажырау себептері, гаметалар тазалығының механизмдері Мендельге белгісіз болды. Себебі ол кезде жасуша туралы мәліметтер жеткіліксіз еді. Атап айтканда, жасушаның тұқым қуалау ақпараты, митозды бөліну, мейоз, гаметалардың түзілуі мен ұрықтануы, т. б. Ал қазір цитология ғылымы жан-жақты зерттеулер нәтижесінде, орасан зор табыстарға жетті. Сондықтан Мендель заңдарына цитологиялық тұрғыдан негіздеме беруге толық мүмкіндік бар.
Организмнің кез келген дене жасушаларында хромосомалардың диплоидті жиынтығы бар екенін еске түсірейік. Бұршактың хромосомалар саны - 14, яғни 7 жұп. Алуан түрлі белгілерді анықтайтын аллельді гендер түрлі жұп хромосомалардың бірдей үйлескілерінде орналасқан. Мысалы, тұқымның түсін анықтайтын аллельді гендері бар бір ғана жұп хромосоманы алайық. Оның әр сыңарында: А - тұқымның сары түсін анықтайтын доминантты және а - жасыл түстің рецессивті гендері болады. Мейозды бөлінудің нәтижесінде пайда болған бұл сыңар хромосомалар қайтадан жұптасып, ондағы гендер де аллельді жұп құрайды (102-сурет) .
Ендеше, Мендель тәжірибесіндегі алғашқы доминантты және рецессивті белгі бойынша гомозиготалы даралар будандасқанда, гендері әр түрлі гаметалардың қосылуына байланысты бірінші ұрпақта біркелкі гетерозиготалы будандар (Аа) алынған. Бүл будандардан екі түрлі гендері бар ұқсас хромосомалар мейозды бөліну кезінде екі гаметаға ажырап кетеді. Осындай екі түрлі, бірінде - доминантты, ал екіншісінде рецессивті гені бар гаметалардың қосылуына байланысты келесі ұрпақта (Ғ 2 ) ажырау жүрген. Ажыраудың арақатынасы ІАА: 2Аа : Іаа сипатта болатыны сондықтан.
Сіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz