Теломерлер мен теломераза: құрылымы, репликациясы, ұзартылу механизмдері және қартаю мен онкогенездегі маңызы

Жоспар:
І. Кіріспе
ІІ. Негізгі бөлім:
- ДНҚ-ның теломерлік бөлімдерінің репликациялануы
- Буферлік теломердің қалыптасуы
- Теломеразаның көмегімен теломердің ұзаруы
- ALT механизмі
- Теломер және теломераза
- Теломераза және қартаю
- Қартаюдың теломерлік теориясы
- Теломераза және онкогенез
ІІІ. Қорытынды
Теломераза - қартаюды ақырындап тоқтатын қасиеті бар, жаңадан ашылған фермент. Олар клеткада синтезделіп, теломердің бірқалыптылығы мен күшіне жауапты. Теломера - ДНҚ жіпшелерінің ұштарында табылған құрылымды белок. Теломер (грек тілінен telos - ұшы, meros - бөлігі) - хромосома ауданының ұштарында орналасқан. Хромосомада екі теломер болады. Теломера хромосма репликациясын қамтамассыз етеді. Клеткалар бөлінгенде теломер жоғалуы немесе бұзылуы мүмкін, бұл клетканың бөлінуін зақымдайды. Клетканың репликация процесін немесе клетканың толық бөлінуін тоқтатады.
Теломера ДНҚ хромасомаларының ішінде «жоғарға бұралған» қалпында болады. Клетка хромасомаларының бөлінуі кезінде ДНҚ ұштары шатасып кетуі мүмкін. Клетканың бөлінуі кезінде хромасома тұтастығын сақтау үшін, олар ерекше кезектестікпен орналасып, көптеген рет қайталанады. Теломераза теломердің күшін қалпына келтіріп, клетканың дұрыс бөлінуінің мүмкіндігін арттырады.
Өкінішке орай, рак клеткалары да өз алдына теломераза қалыптастырып, клеткаларының үздіксіз және шексіз бөлінуін тудыруы мүмкін. Салыстыру үшін қылыпты клеткада репликация процесі белгілі бір бөлінуден кейін тоқтайды. Белгілі бір мағынада рак клеткалары мәңгілік. Оларды қоректендіретін ағзаны бұзғанша дейін өмңр сүреді. Рак клеткаларында теломеразалардың бөлінуін тоқтату үшін зерттеулер жүргізіліп жатыр.
Сонымен қатар зерттеушілер теломераза қызметінің қалыпты клеткаларда қосылуын анықтауда. Бұл белгілі болғанда, біз клеткалардың тежеуіне жңне сау клеткалардың қартаюын алдын ала аламыз. Теломердің ұзындығының қалпына келуі, биологиялық сағаттың орнын ауыстырады. Осының нәтижесі адам өмірінің жалғасы болуы мүмкін.
боялған адам хромосомалары мен олардың теломералары
Теломердің хромосомада орналасуы
ДНҚ молекулаларының толық , яғни теломерлік бөлімдерінің алғаш рет 1971 жылы А. М. Оловников айтқан болатын.
Мұның мәні мынады: ДНҚ - полимеразалық жүйесі аналық ДНҚ тізбегінің 3 ұшын репликацияландырмай қалдырады, яғни жаңадан синтезделетін ДНҚ тізбектері 5 ұшынан қысқарады. Себебі, әрбір жаңа тізбектің 5 ұшында орналасатын Оказаки фрагменті ерекше нуклеазалар әсерінен ұзаратын қысқа РНҚ - ұйытқыштан басталады. Әрбір ДНҚ - полимераза өз бетінше жаңа ДНҚ синтезін бастай алмайды, тек қана бар полинуклеотидтерді 3 ұшынан ұзартады. Сондықтан жаңа тізбек аналық тізбектен қысқа болады. ДНҚ молекуласының мұндай ұшын ( бір тізбегі екіншісіне қарағанда ұзын) үшкір ұшы немесе оверхенга деп аталады. ДНҚ - ның үшкір ұшы тұрақсыз болады. Себебі, эндонуклеазалар ұзын нуклеотидтерді бір - бірлеп алып тастап, ДНҚ ұшын тұйықтайды. Егерде жасушада теломераза болмаса, онда оның әр бөлінуінен кейін хромасоманың қысқаруына әкеледі.
Әрбір репликацияда ДНҚ молекуласы «РНҚ - ұйытқыш» ұзындығына сәйкес 10-15 нуклеотидке қысқаруы тиіс болғанымен, шындығында 50-65 нуклеотид жұбына қысқарады. Бұл ДНҚ - полимеразалық кешенінің қасиетіне байланысты болады.
Адамның ядролық ДНҚ - ның бір молекуласының орташа саны 120 миллион нуклеотид жұптарына тең. Теломераза болмаған жағдайда бір клетка бөлінуінде ғана ДНҚ 0, 5 % қысқарады. Бұл әрине өте аз. Бірақ табиғатта теломера ұзындығын қалпына келтіретін механизмдер болмаса, онда хромасоманың клеткалық бөлінуінің саны жоғалып кететін еді. Осылайша, хромасома ұшының репликацияланбауы үлкен бтологиялық маағынаға ие. Ол қартаю және карцерогенез сияқты тақырыптарымен тығыз байланысты.
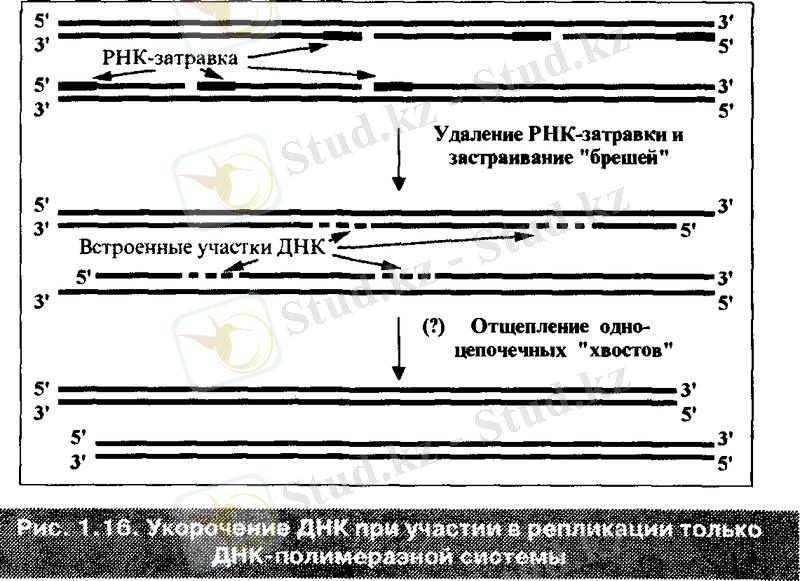
Клеткада ұштарының репликацияланбау проблемесы қалай шешіледі?
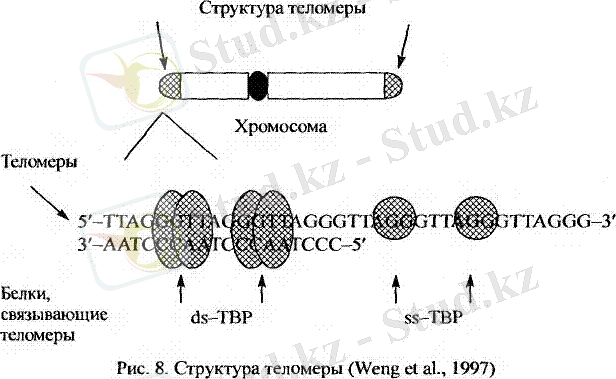
Ғылыми деректер бойынша хромасома ұштарында генетикалық ақпарат болмайтын арнайы гексонуклеотид ( алты нуклеотидтен тұратын ) бірізділіктер қайталанып орналасады.
(5) ЦТААЦЦ . . . ЦТААЦЦ . . . ГГТТАГ . . . ГГТТАГ (3)
(3) ГАТТГГ . . . ГАТТГГ . . . ЦЦААТЦ . . . ЦЦААТЦ (5)
Берілген схема бір гексонуклеотидтің құрылысын анықтайды.
(5) ГГТТАГ (3)
3 ұшынан басталатын ДНҚ - ның көптеген қайталамалары бар. Осы қайталамалардың әрбір құрамы бойынша теломер ауданыда ДНҚ G тізбегі және С тізбегі деп ажыратылады.
ДНҚ - ның бір тізбегі теломердің бірінші ауданында G тізбегі, ал екінші ауданында С тізбегі сияқты қатысады.
ДНҚ - ның теломерлік бөлімдерінде мыңдаған осындай гексонуклеотидтер қайталанады. Олардың жалпы ұзындығы адам эмбрион жасушаларында 10-15 мың нуклеотид жұптарына тең. Сонымен, хромасоманың екі теломерлік ұшы, адам ядролық ДНҚ молекуласының ұзындығының 0. 002 % құрайды.
Теломерлік қайталануларда ешқандай генетикалық ақпарат болмайды. Сондықтан ол теломеразасыз олардың біршама бөлігі түсіп қалған күннің өзінде де геном бірқалыпты қызмет етеді. Теломерлердің негізгі қызметінің өзі де болса керек, яғни олар геномның маңызды бөлігін толық репликацияланбаудан қорғап, буферлік қызмет атқарады.
Теломеразадан біржолата бас тартуға болмайды, себебі клетканың бөліну процесінде күндердің - күнінде ДНҚ - ның участкілері қысқайыр жойылуы мүмкін. Сонымен қатар, теломерлік участкілер ерекше, арнайы қызметтер де атқарып, белгілі бір шекке дейін ғана қысқарады.
Бұл проблеманы екінші жолмен шешу үшін - бүкіл және бастапқы клеткаларда репликацияланбаған ДНҚ участкілерін құрастырып бітіру. Осымен теломераза айналысады.
Теломераза жаңа қысқарған тізбекті емес, аналық ұзарған тізбекті ұзартады.
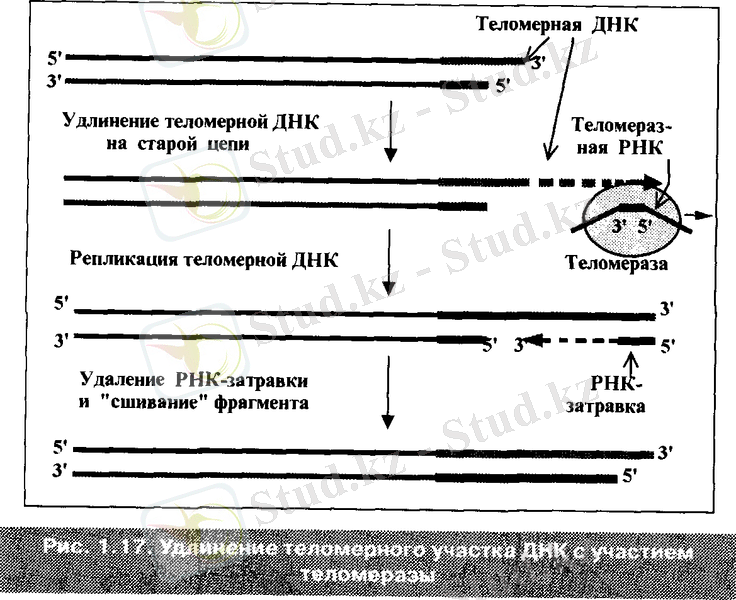
Теломераза аналық тізбектің 3 ұшында бірқалыпты жүйемен ондаған немесе жүздеген жоғраыда көретілгендей қайталамалардан - ГГТТАГ құрастырады. Сонымен теломераза G тізбектің бірінші немесе екінші теломерін ұзартады. Содан кейін ұзарған аналық тізбек матрицалық қалып ретінде тағы да бір Оказики фрагментінің жаңа С - тізбегін құрастырады. Ескі тізбектің 3 ұшының бастапқы ауданында праймаза «РНҚ - ұйытқысын» синтездейді. Содан кейін β ДНҚ - полимеразасы бірінен кейін бірі дезоксинуклеотидтік ұйытқыға қосылады (аналық тізбектің теломер қайталамаларына комплементарлы) .
Фрагменттің өсуі 5→3 бағытында жүреді және бұрынғы жаңа тізбектің 5 ұшының қосылуына жеткенде аяқталады. Фрагменттің тізбекпен тігілуі ДНҚ - лигаза ферментінің әсерінен жүзеге асады. Соңында эндонуклеаза жаңа тізбектің РНҚ - ұйытқысын алып тастайды.
Нәтижесінде ДНҚ - ның екі тізбегі бастапқы қалпына келеді, бірақ та теломерлік қайталамалардан ұзарады.
Зерттеу нәтижесінде барлық тірі ағзалардан табылған теломер ұзындығы осылайша қалпына келеді.
Кейде альтернативтік механикалық теломердің ұзаруы да кездесуі мүмкін (ALT - Alternative Lengthening of Telomeres) . Олар теломеразаның қатысуынсыз жүреді. ALT дрозафила мен опухольдық клеткеларда кездеседі.
Осы механиздердің бірі - теломерлер арасындағы әртүрлі хромасомалардың рекомбинациясы.

Бұл жағдайда екә ДНҚ молекуласы өздерінің теломерлік ұштарымен бірлесіп, гибридтік теломер құрайды. Соңғысында бір ДНҚ тізбегі басқа ДНҚ тізбегіне қарағанда ұзынырақ болуы керек.
Сонда ұзынырақ тізбек матрица қызметін атқарып, сол бойынша ДНҚ - полимераза жүйесі қысқа тізбекті құрастырады. Екі гибридті теломердің бірі өсіп келе жатқан тізбектің лидері ( жай ұзару есесінен жүреді ), ал екінші теломерде артта қалушы ( Оказаки фрагменті түрінде қарама - қарсы ұшынан өседі) болады.
Процестің аяқталуында гибридті емес теломер қалпына келеді немесе ДНҚ молекулалары теломер фрагменттерімен өздеріне сәйкес келетін тізбекпен ауысады. Осылайша, мейоттық емес клеткаларда мейозға тән құбылыстар жүреді.
Жалпы алғанда альтернативтік механизмнің ролі аса үлкен емес.
Теломерлік ДНҚ - ның өзіне сәйкес құрамы бар, ал оның ұзындығын бірқалыпты ұстау үшін арнайы теломераза ферменті қолданылады.
Теломера мен теломеразаға қатысты мәселелер:
А) Теломердің құрылымы
Теломерлік ДНҚ қайталамалардың нуклеотидтік структурасы белгілі. . Арнайы белоктардың арқасында, одан да жоғары дәрежеде ұйымдасады.
Бұл белоктар, гистонды белоктарға қарағанда нуклеосомалық глобулалар түзбейді. Нуклеосомалық құрылымдар қысқа теломерлерде табылмаған. Бірақ та тышқанның ұзын теломерлерінде ұйымдасу бар.
Теломерлік белоктар арасындағы ең әйгілісі - Rар1 белогы және оның аналогы TRF1 белогы (бауырмен жорғалаушыларда) . Осы белоктардың арқасында нығыз тығындалып, гетерохроматиннің құрамына кіреді.
Осындай құрылым теломерлерді бірқалымтылығын қамтамассыз етеді. Теломерлік қайталамалар клетка циклінің барысының көп бөлігінде теломера үшін жеткіліксіз. S - фазада белгілі бір белгіге жауап ретінде TRF1 белогы теломерадан диссоциацияланып, теломердің ұзаруы басталады. Содан кейін TRF1 белогы қайрадан теломерамен байланысып, оның шамадан тыс ұзаруын алдына алады. Осы себебпен ДНҚ теломерлік участкілері ДНҚ - метилаза және эндонуклеаза ферменттері үшін аз жеткілікті. Осының салдарынан мейоз процесінде теломер аудандарынада екі тізбектің айырылуы өте төмен жиілікте жүреді.
Теломерлік белоктардың көмегімен теломера ядроның матриксті компоненттерімен байланысады. Бүкіл клеткалардағы теломерлер ядро мембранасына жалғанғандығы толық дәлелденбеген.
Бірақ та, мейоз процесінің ерте және орта профаза кезеңдерінде мұндай байланыс бар екендігі дәлелденбеген.
Б) Теломераның функциялары
Теломералар төмендегідей маңызды қызмет атқарады:
- Механикалық қызмет:Теломералар хромасомалардың ядро матриксіне бекінуіне қатысады. Бұл хромасоманың ядроға дұрыс орналасуына қажет. Бұл мейоз процесінде байқалады.
Мейоздың профаза сатысының зиготнема кезеңінде хромасома ұштары беткі ядро мембранасына бағытталып орын ауыстырады.
- Бұдан басқа, теломера аналық хроматидтедің бір - бірімен байланысады ( хромасомада S - фазадан кейін қалыптасады ) .
- Екінші қызметі - бірқалыптылылығы:Егерде клеткада теломераза немесе ALT механизмі болмаса, онда теломердің болуын генетикалық мағынасы бар репликацияланбаған ДНҚ бөлімдері алдына алады. Егер де клеткада теломераза белсенділігі болса, үзілеген хромасома ұштарының бірқалыпты болуына тағы да бір мүмкіндік пайда болады.
Кездейсоқ хромасомалардың үзілуі кезінде фрагменттер пайда болады. Олардың бірінші немесе екінші ұшында да теломерлік қайталамалар болмайды. Теломераза болмағандықтан, бұл фрагменттер дегредацияға тап болып, клетканың циклін болдырмай, оның өліміне әкеледі.
Теломераза болған жағдайда үзілу болған жерлерге ДНҚ теломері қосылады. Бұл хромасома фрагменттерінің бірқалыптылылығы мен олардың функционалдылығына мүмкіндік береді.
Бұл феномен ά - талассемия ауруымен ауратын адамдарда табылған. ά - глобина гендерінде 16q хромасомалары үзіледі және жараланған ұштарына теломерлік қайталамалар жалғанады.
- Гендер экспрессиясына әсері.
Теломердің тағы да бір ерекше қасиеті - дәреже эффектісі: теломер жанындаға орналасқан гендер белсенділігі төмен болады. Бұл эффект транскрипциондық үнсіздік немесе сайленсинг мағынасын жиі береді. Теломердің қысқаруында дәреже эффекттісі жоғалып, прителомерлік гендер активтенеді.
- Сайленсинг теломерамен қызмет ететін белоктардың нәтижесі болуы мүмкін.
- Басқа жағынан алғанда, дәреже эффектісі ядро қабығына жақындығымен нақтыланған. А. М. Оловников гипотезасы бойынша, бұл қабықта Са - каналы мен иондар ағыны орналасуы мүмкін. Са белоктың жанында орналасқан гендермен бірігуіне әсер етеді.
Дәреже эффектісі ішкі гендерге де қатысы бар. Егер де қандай да бір осы гендердің біреуі транспозондық ( басқа ДНҚ участігіне орын ауыстыра алу қасиеті бар гендер ) болып, теломер ауданына қосылады. Немесе егер де хромасомалардың үзілуі жолып жатса, онда үзілудің соңында теломерлік қайталамалар құралады. Соңғы теломерлік қайталамалардың көмегімен теломер белоктарының байланысуы мүмкін және ядро мембранасына байланыстырады.
- «Есептеу» функциясы
ДНҚ - ның теломерлік бөлімдері сағат қызметін атқаратын құрылғы ретінде де белгілі. Ол теломер белсенділігі жоғалғаннан кейін клетканың бөліну санын есептейді. Әрбір бөліну теломерді 60 - 65 нуклеотид жұптарына қысқартады. Теломер - теломераза болмаған жағдайда қалыпты клетканың бөлінуін анықтайтын құрылғы. Белгілі бір қысқа ұзындыққа жеткеннен кейін, теломер өзінің бүкіл функцияларын жоғалтады. Клетканың циклі бұзылып, соңында клетка өледі.
В) Теломеразалар әрекетінің механизмі
Теломеразалар әрбір теломералардың G тізбегін ұзартады. Теломеразаға ұзындығы 450 нуклеотидтен тұратын теломеразалық РНҚ байланысқан. Оның ортаңғы қысқа участкесі бір жарым теломерлік қайталануға комплементарлы болады.
(3) АУЦ ЦЦА АУЦ (5)
Осы РНҚ - ның сол жағындағы триплет (АУЦ) ДНҚ - ның G тізбегінің шеткі теломерлік жарты қайталамаларымен байланысу үшін пайдаланылады.
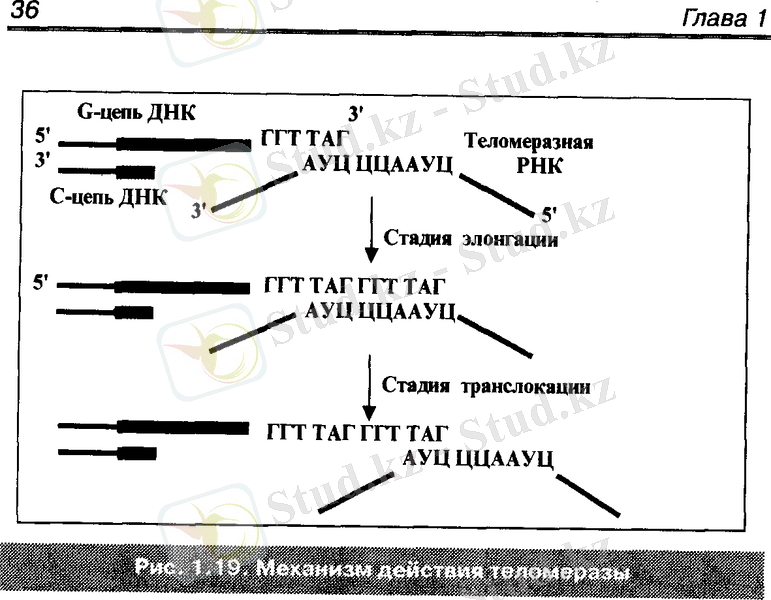
Қалған гексонуклеотид (АУЦЦЦА) G тізбекті 3 ұшынан ұзарту үшін матрица ретінде қызмет етеді. Ал нуклеотидтердің жүйелі қосылуы (матрицаның құрылысына байланысты) теломеразаның белоктық субъединицаларымен катализденеді.
Теломераза кері транскриптаза ферменті ретінде, ДНҚ синтезін РНҚ матрицада орындалуына қатысады. Осы уақытта ДНҚ тізбегінің ұзаруы 5→3 бағытында жүреді.
Жаңа теломерлік қайталамалардың құрылуы элонгация процесінің бастамасы болады. Содан кейін транслокация процесі - ферменттерді өз РНҚ - рымен бірге өсіп бара жатқан ДНҚ тізбегінің бір қайталамасын «солға қарай» 3 ұшына қарай орын ауыстырды. Транслокация процесінде РНҚ ДНҚ - мен байланысын жоғалтып, содан кейін оны жаңа синтезделген қайталаманың шеткі «жартысымен» алмасады.
Содан кейін ДНҚ тізбегі тағы да бір қайталамаға ұзарады. Солай жалғаса береді. Осылайша теломераза циклінің жұмысы өзіне екі кезеңді қосады - элонгация және транслокация. Екеуін алмастыра отырып, фермент ескі тізбекке бірнеше ондаған немесе жүздеген теломер қайталамаларын қосады.
Теломерлік РНҚ - жалғыз ғана матрица емес, сонымен қатар каталиттік машинаның маңызды бөлігі. Оның бірнеше нуклеотидтерін ауыстырғанда «дұрыс емес» қайталамалардың синтезі емес, теломеразаның өз белсенділігін жоғалтуы болады.
Г) Теломераза белсенділігін анқтау әдістері
Биологиялық нысанда теломераза белсенділігін анықтаудың екі негізгі әдісі бар: тікелей және жанама.
- Тікелей әдісінде- инкубациондық қоспаға зерттеліп жатқан үлгіден басқа, G тізбегінің праймерін (олигонуклеотидтер, құрамында бірнеше теломер қайтталамалары бар) және белгіленген дНТФ қосады. Онымен байланысатын және оны теломераза ұзартатын праймер G тізбегінің рөльінде шығады. Оның белсенділігін радиоактивтің құрамына олигонуклеотидтер қосылғанда ғана анықталады.
- Жанама әдісінде- TRAP анализі (Telomeres Repeat Amplification Protocol) . Онда ТПР - тізбектің полемеразалық реакциясын колданады. Мұнда қоспаға үлгімен қатар, G - праймер мен белгіленген нуклеотидтерді қосады. Сонымен қатар қоспаға С - праймерін және термошыдамды бактериялық ДНҚ - полемеразасын енгізеді. Осының арқасында ТПР - дың барысында теломеразамен ұзарған олигонуклеотидтерде көптеген рет «клондау» жүреді.
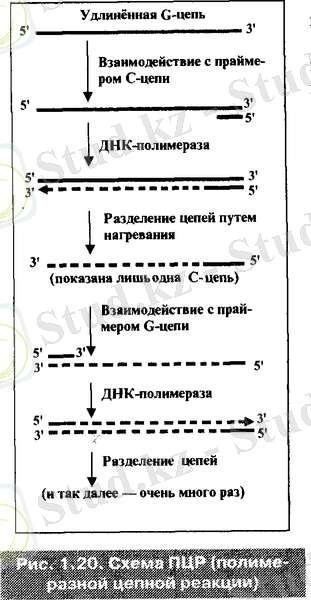
Ұзарған G - тізбекпен С - праймер әрекеттесіп, ДНҚ - полимеразамен өсіріледі ( G - тізбегі матрицалық қалып ретінде пайдаланылады) . Екі тізбекті ДНҚ құрайды. Оның тізбектерін қыздыру арқылы ажыратады.
Ұзарған С - тізбегімен G - праймер әрекеттесіп, ДНҚ - полимеразасымен өсіріледі. Екі тізбекті құраған тізбек тағы да қыздыру арқылы ажыратылып, ортасында қосымша ұзарған G - тізбегі пайда болады. Бірақ ол теломеразамен емес, бірінші G - тізбегінің екі сатылы көшірілу жолымен ДНҚ - полимеразада синтезделеді.
Осылай жалғаса береді. Көптеген рет циклдер алмасып, өзіне С немесе G - праймерлердің өздеріне сәйкес келетін жалғыз тізбекпен әрекеттесуін қосады, бұл тізбектің ДНҚ - полимеразалық көшірмесі және екі тізбектің қыздырылуы арқылы ажырауы.
Бұл бүкілі бір пробиркада болады, себебі термостаттық ДНҚ - полимераза көптеген циклдердің әрбіреуінің соңындағы қыздыруға шыдайды.
Нәтижесінде G - тізбектің саны орташа есеппен 10 рет көбейіп және соған сәйкес әдістің сезімталдығы да өседі.
TRAP - анализінің пайда болуы теломеразаның әр түрлі нысандарда табуын жеңілдетті.
Бұл әдістер теломеразаның қандай клеткаларда бар және оның функциясын, және оның қандай клеткаларда болмайтындығын анықтайды.
Д) Органимзнің қалыпты клеткаларында теломеразаның таралуы
А. М. Оловниковтың алғашқы болжамы бойынша, жасы үлкен адамда теломерлік белсенділік тек қана жыныс клеткаларының тізбегінде ғана болады. Ал бүкіл соматикалық клеткаларда ол болмайды және тек қана солардың опухольдық қайта тууында пайда болады. Экспериментальдық зерттеулер нәтижесінде толығымен бұл болжам дәлелденген сияқты болды.
Ең алғаш рет теломераза 1985 жылы бір жасушалы эукариоттарда табылды. Содан кейін адамның опухаль клеткаларында, ал кейін аналық және аталық жыныс бездерінде табылды. Қалыпты соматикалық клеткаларда теломеразалық белсенділікті детектировациялауға мүмкіндік болмады. Осының барлығы ескі гипотездік көзқарасты практикалық аксиомаға айналдырды.
Жоғары сезімталдығы бар TRAP - анализі пайда болғаннан кейін әртіүрлі ұлпаларда, жануарлар мен адам мүшелеріндегі теломераза белсенділігін анықтау басталды.
Егер де жасушаның бөлінуін ескерсек, теломераза белгілі бір заңдылықпен жасуша мен ағзада таралады.
А) Теломераза қалыпты соматикалық клеткалар арасында ең жиі тұрақтылығымен қан түзетін мүшелерде табылды: сүйек майы, сөл түйіндері және т. б. .
Сүйек майындағы оның белсенділігі бастапқы бағаналық клеткаларда жоқ, бірақ та жоғарғы дәрежеге белсенді бөлінетін клеткаларда жетіп, содан кейін қайрадан жоғалады.
Шеткі қан клеткаларында (гранулоциттер - боялған кезде цитоплазмасында түйіршіктер боялатын лейкоциттер, лимфоциттер) теломераза табылған . Оның мұнда белсенділігі аса үлкен емес, бірақ та лимфоциттердің бөлінуіне түрткі болатын митогендерде ол көбейееді.
Сонымен қатар теломеразаны эпителии терісінде және әртүрлі мүшелердің кілегейлі қабығәртүрлі мүшелердің кілегейлі қабығында кездеседі.
Эндометрияда теломераза белсенділігінің шыңына жетуі пролиферация (клеткалардың бөлініп көбеюі нәтижесінде белгілі бір мүшенің ұлғаюы) фазасының соңында болып, ал келесі фазасында секреция белсенділігі тез төмендейді.
Бұған ұқсас жұмыс басқа эпителиальдық ұлпаларда да бар: клеткалар терминальдық дефференцияға кіріп, теломераза белсенділігін жоғалтады.
Б) Бұл фермент өте аз жиілігімен шартты постмитотикалық клеткалары бар мүшелерде: бауыр, ұйқы безінде кездеседі.
С) Ми мен бұлшық ет ұлпаларының көп бөлігін құрайтын постмитотикалық клеткаларда теломераза табылмады.
Сонымен, теломераза соматикалық клеткалардың бөліну қасиеті бар ғана клеткаларында болады.
Берілген белсенділік бөлінуші клеткаларда теломераның қызқаруынан сақтамайды. Бөліну саны өскенде (өсін) немесе жануардың календарьлық жасында (in vivo) қан түзуші клеткаларда және лимфоциттерде теломердің қысқаруы болады.
Бұның түсініктемесі: теломераза белсенділігінің төменділігі немесе оның қандай да бір ингибаторлармен тежелуі.
Қартаюдың теломерлік теориясын екі негізгі жағдай бойынша А. М. Оловников құрастырған: 1) ДНҚ ұшының репликацияланбау проблемасы; 2) Хейфлик эффекті
1961 жылы аталған эффектің ашылуы қартаюдың табиғаты туралы жалпы көзқарасты өзгертті. Тағы да он жылдан кейін бұл эффект қартаюдың қартаюдың теломерлік теориясының негізгі қағидасы болды.
А) Вейсманның алғашқы идеялары
1981 жылдың басында цитология классигі Август Вейсман айтқан: организмнің қартаю үдерісінің себебі, соматикалық клеткаларда бөліну арқылы өсу қасиеті мәңгілік емес, шектеулі. Басқа сөзбен айтқанда, соңында жасуша тозады және бөліну арқылы өзіндік жаңара алмайды. Осының барлығы жасушаның өліміне әкеледі.
Вейсман жыныс клеткаларына немесе ұрықтық плазма үшін өзгерістер жасады. Қазіргі терминдер бойынша, жыныс жасушаларында қандай да бір жастық өзгерулер жоқ. Егер де ондай болса, ол өзгерген аномалиялар болады.
Бұл айтылған экспериментте негізделмеді, тек қана әйгілі ғалымның болжамдары болып қалды.
Б) Вейсманның алғашқы идеяларының Карралемның теріске шығаруы
20 ғасырдың басында жаңа пайдалы әдіс - ұлпа бөліктерін өсінде өсіру пайда болды.
А. Карреля эксперименті мынадан тұрады: ол үлкен емес тауық жүрегінің бөлігін алып, әйнек ыдыстағы желім сияқты эмбриональдық экстракт пен плазманың қореқтендіруші қоспасына салып, термостатта жетілдіреді. Ұлпа бөліктерінің перифериясында (шеткі жағы) клетка бөлініп, бірнеше күннен кейін ені бойынша үлкейіп, әрбір бөлігі жағынан жуықтап алғанда 0, 3 мм, ал ауданы бойынша екі есе өсті. Оны тағы да екіге бөліп, оның бір фрагментін сол шаттары бар келесі әйнек ыдысқа салды.
... жалғасыСіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz