Өсімдіктердің вегетативтік көбеюі мен қолдан вегетативтік көбейту әдістері

Кіріспе.
Барлық тірі организмдер сияқты өсімдіктерге де қоректену, тынысалу, қимыл, тітіркену, өсу, даму және көбею сияқты қасиеттер тән. Бұл қасиеттер бір-бірімен өте тығыз байланысты. Осының ішінде көбею өзінің биологиялық мәні тұрғысынан басқа қасиеттерден ерекше. Өйткені көбею кезінде негізінен аналық особьке ұқсас жаңа организмдер пайда болады.
Көбею - тірі организмнің жеке санының артуы, өзінен кейін ұрпақ қалдырып, түрдің сақталып, табиғатта таралуын қамтамасыз ету қасиеті. Көбею барысында өсімдіктердің кейбір түрлерінде особь сандарының артып, өз ортасында кең таралып, жаңа кеңістіктерді қамтиды. Бұл прогрессивті түрлерге тән. Ал кейбіреулерінде особь саны тұрақты, олардың таралу кеңістігі, ареалы көлемін ұлғайтпайды не кемітпейді.
Өсімдіктердің келесі бір түрлерін особь сандары, соған сәйкес олардың таралу кеңістігі үнемі кеміп отырады. Бұл регрессивті түрлерге тән. Бұлар табиғи түрінде жойылып кетеді.
Көбеюдің үш кейпін ажыратады. Олар: жынысты көбею, жыныссыз көбею және вегетативтік көбею. Жынысты көбею кезінде жаңа организм әр түрлі жыныс жасушаларының қосылуынан пайда болады. Жыныссыз көбеюде жаңа организм жыныс тұрғысынан жіктеле қоймаған бір жасушаның өзінен жетіледі. Вегетативтік көбеюде жаңа организмге бастаманы вегетативтік мүшелер, олардың бөліктері және түр өзгерістері (метаморфоз) береді.
Вегетативтік жолмен көбею.
Вегетативтік көбею деп вегетативтік денеден тіршілікке қабілетті бөліктер бөліну арқылы түрдің жеке санының артуын айтамыз. Вегетативтік көбею алуан түрлі және өсімдіктердіңбарлық топтарында кең таралған. Бір жасушалы балдырлар мен бактерияларда вегетативтік көбею жасушаларының екіге бөлінуі арқылы жүзеге асады. Колониалды балдырларда, олардың жасушаларының вегетативті көбеюінен жас колониялар пайда болады.
Көп жасушалы балдырларда вегетативтік көбею, олардың талломдарының кездейсоқ бөліктерге бөлінуі арқылы жүреді. Жіпшелі көк-жасыл балдырлардың кейбір түрлері (носток, анабена) гетероцисталары арқылы көп жасушалы гормогонияларға бөлінеді. Көбеюдің мұндайқарапайым тәсілі тек төменгі сатыдағыларға ғана емес, суда тіршілік ететін кейбір гүлді өсімдіктерге де (сужапырақ, мүйіз жапырақ, егеушөп) тән. Осылай көбею нәтижесінде XIX ғасырда Еуропаға енген Канада сужапырағы су қоймаларында кең таралғандықтан, жергілікті тұрғындар «су чумасы» деп атаған.
Саңырауқұлақтарда вегетативтік көбеюдің қарапайым жолы - мицелийдің жеке учаскелерге бөлінуі, бүршіктенуі (ашытқылар) арқылы жүрсе, арнаулы маманданған түрі кондиилер, оидийлер және хламидоспоралар арқылы жүзеге асады. Оидийлер мен хламидоспоралардың пайда болуы көбеюдің вегетативтік жолынан жыныссыз жолында өтпелі түрі ретінде қарастырылады.
Қыналар, денесінің арнаулы бөліктері - соредийлері, кейде сирек изидийлері, ал бауыр мүктері өнімтал бүршіктері арқылы көбейеді. Өсімдіктердің вегетативтік көбеюі оның регенерациялық қабілетіне негізделген. Регенерация (латынша «регенерацио» - жандану, қалпына келу) - өсімдік денесінде жетіспейтін мүшелердің немесе тұтас өсімдіктің өсімдік денесінің жеке бөліктерінен, тіпті жеке вегетативті жасушасынан қайта қалпына келуі. Көптеген зерттеу жұмыстарының нәтижесінде өсімдіктердің регенарация қабілетінің өте жоғары болатынын байқаған. Мысалы, жасаңшөп жапырақ эпидермасының бір жасушасынан тұтас өсімдік, сол сияқты көптеген өсімдік топтарының гүл табанынан, гүл желегінен, аталығынан тұтас өсімдіктер өсірілетіні белгілі.
Вегетативтік көбею кезінде аналық өсімдіктен пайда болған жас особьтар жиынтығы клон деп аталады. Гүлді өсімдіктерде клонда тамырсабақтың бұтақталған ескі бөліктерінің біртіңдеп бұзылуынан, каудекс, кіндік тамырдың партикулдануынан, сол сияқты жер беті және жер асты столондарының ұшындағы түйнектер мен жуашықтардан жас өсімдіктер бастамасы ретінде жетіледі. Вегетативтік көбею табиғатта өздігінен жүретін табиғи және адамның қатынасуымен жүзеге асатын - жасанды деп екігебөледі. Бұл екеуінің арасында айтарлықтай айырмашылық жоқ. Өйткені табиғи вегетативтік көбею өсімдіктердің регенерациялық қасиетіне, ал екіншісі түрлі әдістерді қолдану арқылы қолдан вегетативтік көбейтуге негізделген.
Жоғары сатыдағы сабақты-жапырақты өсімдіктер тамыр атпалары, тамыр сабақтары, мұртшалар, желілері (көген сабақтары), түйнектері, столондары, жуашықтары, өнімтал бүршіктері арқылы табиғи вегетативтік жолмен көбейеді.
Көптеген сүректі өсімдіктер және біраз шөптесін өсімдіктердің кейбіреуі тамыр атпалары (тамырдың эндогенді қосалқы бүршіктерінен өскен жас өркеңдер) арқылы көбейеді. Эндогенді бүршіктер жанама тамырлар сияқты жетіледі. Қосалқы бүршіктерден өсіп шыққан жас өркендер көбейіп, тез тарайды. Мәдени түрінде мұндай өсімдіктер тамыр атпалары арқылы көбейтіледі. Тамыр атпалары арқылы көптеген сүйекті жемістер - қара өрік, шие, алма, сол сияқты терек, тал, көк терек, қандағаш, жөке, ақ қараған, серігүл, сиыр бүлдірген көбейеді. Шөптесін өсімдіктерден - сүттіген, шырмауық, бақбақ, желкек, қалуен т. б.
Тамыр сабақтары арқылы көбею негізінен көп жылдық шөптесін өсімдіктерге тән. Тамырсабақты өсімдіктерге: жыланқияқ, папоротник, селеу, қамыс, қоңырбас, кияқ, жатаған бидайық, адыраспан т. б. өсімдіктер жатады. Бұлардың жер астындағы өркендерінің буыны және буын аралықтары бар. Буындағы бүршіктерден жас өркендер өсіп, вегетативтік көбеюді қамтамасыз етеді. Қысқарған тамырсабақтарда бүршіктер жиі орналасады да, олардың жер бетіңдегі өркендері де жиі шоқтанып жатады, ұзарған тамырсабақтарда бүршіктер сирек орналасады, сондықтан олардың жер бетіңдегі өркендері де сирек. Ұзарған тамырсабақты өсімдіктер жан-жаққа тарамданып, тез арада жайылып өседі.
Көптеген жатаған өсімдіктер мұртшалары арқылы, ал төселмелі сабақты өсімдіктер желілері арқылы көбейеді. Жатаған өсімдіктерге ақ бас беде, қой бүлдірген, қазтабан, жатаған, сарғалдақ, тас бүлдірген т. б. жатады.
Жатаған өркеннің топырақпен жанасқан буынынан қосалқы тамыр жетіліп, осы жердегі жапырақтың қолтық бүршігінен жас өркен шығып, көбеюін жалғастырады. Жатаған өркендердің буын аралықтары өліп, жас өсімдік пен аналық өсімдіктің байланысы үзіледі. Жас өсімдік те, аналық өсімдік те өз алдына жоғарыдағыдай көбею жолын қайталайды, яғни әрқайсысынан жаңа мұртшалар - жатаған өркендер жетіледі.
Төсемелі желі сабақты өсімдіктерде оның ылғалды топырақпен жанасқан жерінде жатаған өсімдіктердегі сияқты қосалқы бүршіктерінен жас өркендер пайда болады. Желі сабағы арқылы көбею асқабақ тұқымдастарының өкілдеріне тән. Мысалы, колдан отырғызған асқабақ, қауын, қарбыз, қияр желілеріне өсімдік вегетациясының соңында ылғалды топырақ төгіп, сонымен жанастыру арқылы көбейтуге болады.
Вегетативтік көбею қызметін атқаратын түйнектер - тамыр мен өркеннің түр өзгерістері. Түйнектердің өзі жер бетіңдегі түйнектер және жер астындағы түйнектер болып бөлінеді. Бұлардың бет жағындағы орналасқан бүршіктерден (көзше) жас өркендер өсіп шығады. Түйнектері арқылы жер алмұрты, цикламен, альпі қоғажайы, нарғызгүл, кольраби орамжапырағы және т. б. өсімдіктер көбейеді. Өнімтал бүршіктер - кейбір папоротниктерде, мүк тәрізділер мен гүлді өсімдіктерде вегетативтік көбею қызметін атқаратын арнайы мүше. Өсімдіктерде олардың мөлшері тым көп болғандықтан, тұқым сияқты шашылып, қолайлы ортаға түссе, тез өсе бастайды. Мысалы, жасан шөптер тұқымдасына жататын бөлме өсімдігі бриофиллюмның жапырақ жиегіндегі тісшелерінің арасында (ойыс жерінде) қосалқы бөлшектер бар. Бұл бүршіктерден 2-3 жапырағы, 1-2 қосалқы тамыршасы бар кішкентай өсімдіктер пайда болды да, жапырақтан түскен жерінен қаулап өсе бастайды. Осы сияқты өнімтал бүршіктер кейбір шықшылдықтар жапырағына, папоротниктер жапырақ жиегіне, жүйкелеріне тән. Кейбір өсімдіктерде өрімтал бүршіктер түрі өзгеріп жуашық (тісшөпте) немесе түйнекше (мықыр таспада) түрінде кездеседі.
Табиғи вегетативтік көбеюді адам өз мақсатына пайдаланып, вегетативтік көбейту жолымен мәдени өсімдіктерді өсіреді.
Қолдан вегетативтік көбейту. Қолдан вегетативтік көбейту ерте заманнан қолданылып келе жатқан әдіс. Қолдан вегетативтік көбейтудің бірнеше жолы бар. Соның бірі - телу әдісі.
Телу дегеніміз бір өсімдіктің бүршікті бөлігін екінші өсімдіктің денесіне ұластыру. Телуді кейде трансплантация деп те атайды. Өз тамырыңда өсіп тұрған өсімдікті телітуші (подвой) деп, ал оған әкеліп ұластырылатын бүршігі бар бөлікті телінуші деп атайды.
Телу үшін өсімдіктер таңдап алынады да бірнеше мынадай талаптар орыңдалуы қажет: телінушінің өткен жылдары жаппай гүлдеп, мол жеміс берген, ауруға шалдықпаған сау бұтақтары алынады, телітушінің тамыр жүйесі жақсы жетілуі, телінушіден шыққан өркендерді қоректі затпен қамтамасыз етеалатындай қуатты болуы керек; ауруға шыдамды, ортаның қолайсыз әсеріне төтеп берерліктей болуы қажет.
Осындай шарттар орындалғаңда ғана жақсы нәтижеге жетуге болады, Мысалы, жеміс-жидекті өсімдіктерден жүзімді алайық. Оның аса қауіпті ауруы - филлоксера. Жүзімнің өнімін төмендетіп жіберетін осы дерт. Ғалымдар мәдени жүзімнің сорттарын жабайы жүзімге телу арқылы бұл аурудан құтқарды. Өйткені жабайы жүзім тамыры филлоксера ауруына төтеп бере алады. Сол сияқты раушан гүлін итмұрынға теліп телітушінің хош иісті, әсем гүлдерін алуға болады. Осы қасиеттеріне бола негізінде ғалымдар лимон, мандарин, алхоры, алманың мәдени сорттарын өзді-өзінің жабайы түріне теліп олардың жаңа сорттарын алды.
Компоненттер бір-біріне тез ұласуы үшін телінуші мен телітушінің түзуші ұлпасы - камбий, тағы басқа аттас ұлпалары бір-бірімен сәйкес келуі қажет. Компоненттердің арасындағы енген жасушалардан пайда болған қабат біртіндеп айналасындағы тірі жасушаларға сіңіп кетуі, телінетін екі өсімдік жүйелілік тегі жағынан бір-біріне жақын болуы керек. Алшақ болса, оңайлықпен теліне қоймайды және күткен нәтижені бермейді. Мысалы, екі тұқымдасқа жататын өсімдіктерге қарағанда бір туыстың екі түрі өзара тез ұласады.
Телу тәсілдері. Қазіргі кездегі телудің жүзден астам тәсілі белгілі. Олардың көп қолданылып жүргендері - жанастыра телу, кесінділеп (қалемшелері арқылы) телу және көзшелеп телу.
Жанастыра телу үшін қатар өсіп тұрған екі өсімдіктің бірін телітуші, екіншісін телінуші етіп алады. Екеуінің де жанасар жақтағы беттерінің қабығын шамалы сылып тастап, екеуін өзара жанастырады да сыртынан жұмсақ жіппен орап байлайды. Сүректі өсімдіктерді теліген кезде байлауға жөке ағашының суға салып жұмсарылған тін талшығы пайдаланады. Осы қалыпта қалдырылған компоненттер шөптесін өсімдікте 10 күнде, сүректі өсімдік те бір-екі вегетациялық кезеңде ұласады. Телінушінің ұласқандығы бүршіктердің өркендей бастауынан байқалады.
Жанастыра телу кезінде есте болатын жағдайлар: біріншіден, компоненттердің сылынған жерінің көлемі бірдей болуы; екіншіден, телінуші ретінде алынған өсімдік өркенінің кесетін жерден жоғарғы бөліміңде бүршіктерінің болуы. Телінгеннен бір немесе екі вегетациялық кезең өткен соң компоненттер ұласады, осы кезде телітушіні жоғарғы жағынан, телінушіні төменгі жағынан кеседі.
Кесінділеп телу тәсілдері қолданылады. Кесінділеп телудің: жарықшақтап телу, беттестіре телу және т. б. түрлері бар.
Копулировкада - телушіге алынған қалемшенің 2-3 бүршігі болуы кажет, телінушінің қалемшесі мен телітушінің сабағынан қиғаш кесінді жасалады және де компоненттердің камбий қабаттары сәйкес келетіндей диаметрлері бірдей болады. Копулировка жасалған жерге балшық немесе ағаш көмірінің, ұнтағын сеуіп қояды. Бұл телінген жерге ауру тудыратын микроорганизмдердің ұяламауы үшін қажет. Ал ағаштар мен бұталарды теліген кезде компоненттердің диаметрі сәйкес келе бермейді. Мұндай жағдайда кесінділеп телудің басқа тәсілдері қолданылады.
Көзшелеп телу - мұнда телінушіге көзше, яғни бүршік, телітушігеөсіп тұрған өсімдік алынады. Бүршікті алу үшін өркеннің бүршік тұрған жерінің жоғарғы және төменгі жағынан көлденең кесінді жүргіземіз де, жоғарыдан төмен қарай аздап қабығын ілестіре көзшені сылып аламыз. Телітушіден Т әрпіне ұқсас бір-біріне перпендикуляр кесінді жасаймыз. Кесілген жердегі қабықты пышақтың ұшымен көтеріңкіреп, бүршікті ендіреміз. Бүршіктің қалқаншасын бастыра байлап қоямыз. Телінген жерге қайың көмірінің ұнтағын себеміз. Бұл жарақаттанған жерді микроорганизмдердің зиянды әсерінен сақтайды. Көзшелеп телудің басқа тәсілдерден тиімділігі компоненттер мықты болып тез ұласады, телуге материал аз кетеді, арзанға түседі.
Қолдан вегетативтік көбейтудің телуден басқа метаморфозданған мүшелерден, кесінділерден, сұлатпа шыбықтан, түбін ажыратып көбейту және тамыр атпаларынан көбейту сияқты жолдары бар.
Метамофрозданған мүшелерден вегетативтік көбейтуге түйнектерді, пиязшықтарды, қабыршақтарды пайдаланады. Мәселен картоп сабағының метаморфозы түйнегінен өсіріледі. Оның көзшелері - бүршіктер. Сол бүршіктердің әрқайсысы жер бетіне жеке өсімдік болып шығады. Сондықтан картоп түйнегі үлкен болған жағдайда әр бөлігінде 1-2 көзшеден қалдырып бірнешеге бөліп отырғызады. Сол сияқты күрделі гүлділер тұқымдасына жататын жер алмұртының сабағының метаморфоздануынан пайда болған жер асты түйнегі арқылы көбейтеді. Жер алмұртының түйнегі тағамға, малға жем ретінде, ал техникада спирт жасау үшін пайдаланылады.
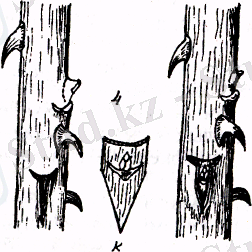
Сурет. Телу тәсілдері:
1 - телу үшін дайыңдалған телінуші қалемше, 2 - жарықшақтап
телу, 3 - копулировка, 4 - окулировка, К- көзше (бүршік) .
Кейбір өсімдіктерді тамырының метаморфоздануынан пайда болған тамыр түйнектерінен көбейтеді. Мәселен, гүлді өсімдік нарғызгүлдің жанама тамырлары түйнекке айналған. Әсемдік үшін отырғызылатын жуашықты өсімдіктердің (лалагүл, қызғалдақ, нарцисс, сүмбіл, амарилис т. б. ) пиязшықтары вегетациясының соңында баданалар береді. Бұл баданаларды 3-5 айдан соң қайта отырғызса. 4-5 жылдан соң қайтадан жетіліп жас өсімдік өседі.
Кейбір пиязшылықты өсімдіктерді түрі өзгерген жапырақтары - қабыршақтары арқылы көбейтуге болады. Мысалы, лалагүлін қабыршағы арқылы көбейтерде баданасының сыртқы қабыршақтарын сындырын алып, оның түп жағын құмға көміп қояды. Бір жылдан соң қабыршақты құмнан шығарып алады да қайта өсіре бастайды. Сонда бұл қабыршақтар 3-4 жылдан соң гүлдейді.
Кесінділеп көбейту үшін өсімдік кесінділері пайдаланылады. Кесінді дегеніміз - бойында бүршіктері бар өсімдіктердің бір бөлігі. Мұны қалемше деп, көбейтудің бұл жолын қалемшелеп көбейту деп атайды. Өсімдіктердің түріне қарай олардың біреуін сабағының, екіншісін жапырағының, ал қайсыбірін тамырының кесіндісінен көбейтуге болады. Сабақ қалемшелері жасыл (жаздық) және қыстық деп бөлінеді. Жасыл қалемшелер арқылы негізінен бөлме өсімдіктерін көбейтеді. Ол үшін аналық өсімдіктен киғаш етіп кесілген 3-5 жапырағы бар (бүршікті) қалемшелер дайындап, оны ылғалды құмға отырғызып қояды. Қалемшелер түптенген (қосалқы тамырлар шығарып) кезде бүршік өсе бастайды.
Сүректі өсімдіктердің басым көпшілігі қыстық қалемшелер арқылы кебейтіледі. Қыстық қалемшелерді бір-екі немесе үш-төрт жылдық өркендерін (ұзындығы 20-25 см) дайындайды. Бұларды күзде ылғалды құмға отырғызады да, көктемде питомникке көшіреді, онда күш алып, ұлғайып өскеннен кейін тұрақты орынға отырғызады.
Сабақ кесіндісінен жүзім, тұшала, тобылғы, жыңғыл, ақгүл ырғай, терек, тал, т. б. өсіріледі.
Тамыр қалемшені жуандығы 0, 5 - 2 см, ұзындығы 16-20 см күзде дайындап, сабақ қалемшелері сияқты өсіреді. Жапырақ қаламшелері арқылы бегония, глоксиния, лалагүл, қазтамақ сияқты өсімдіктер тобының түрлері ғана көбейе алады. Бегония жапырағынан дайындалған қаламшелерде қосалқы тамырлар жапырақ тақтасының астыңғы бетінің ірі жүйкелерінде, ал бүршіктер мен өркендер жапырақтың үстіңгі бетінде жетіледі.
Сұлатпа шыбықтан көбейту. Сұлатпа шыбық дегеніміз - өсімдіктің вегетативтік жолмен көбейтуге арналған өркендерін өсімдіктің өзінде тамырландырып алу. Сұлатпа шыбығы арқылы өсімдіктерді үш түрлі жолмен көбейтеді. Олар: жатқыза сұлату, иілдіре сұлату, тік сұлату.
Түбін ажыратып көбейту - түптеніп өсетін бар сабақтарын бір-бірінен ажыратып өсіру. Түбінажыратып көбейтудің селекцияжұмысында мәні зор. Жеке-жеке тамырлы сабақтарды көбейту арқылы бағалы сорттардың тұқымын алуға брлады.
Тамыр атпаларынан көбейту - тамырдағы қосалқы бүршіктерден жер бетіне өсіп шыққан өркендерді аналық өсімдіктерден бөліп алып өсіру. Бұл түбін ажыратып көбейтуге ұқсас, айырмашылығы өз тамырында өсіп тұрған аналық өсімдікті орнынан қозғамай тамыр атпаларынан өсіп шыққан жас өркенді бөліп алып өсіреді. Таңқурай, алхоры, шие, қара бүлдірген, мойыл, терек т. б. өсімдіктерді осы жолмен көбейтуге болады.
Жыныссыз көбею
Жыныссыз көбею споралары немесе зооспоралар арқылы жүзеге асады. Споралар (грекше spore-тұқым) құрлық өсімдіктеріне тән, олар өз бетінше қозғала алмайды, салмақтары өте жеңіл болғандықтан, ауа толқыңдары арқылы таралады. Зооспоралар (грекше «zoo» - жануар, zoospore - қозғалатын спора) суда, ылғалға қаныққан топырақта өсетін өсімдіктерге тән, бұлар талшықтармен жабдықталған, соның көмегімен өз ортасында қозғала алады. Споралар арқылы төменгі сатыдағы өсімдіктердің басым көпшілігі (балдырлар, саңырауқұлақтар), жоғары сатыдағы мүк тәрізділер мен папоротник тәрізділер көбейеді. Бұл кейінгілерін тұқым арқылы көбейетіндерденжеке-жекетипке бөлді. Мысалы, мүктер типі, папоротник тәрізділер типі, ашық тұқымдылар типі және т. б. деген сияқты. Ботаниктердің басым көпшілігі осылардың ішіндегі соңғы пікірді құптауды.
Жылбұрындардың даму циклі. Жылбұрын - өзі аттас келген тұқымдасқа жататын жоғары сатыдағы споралы (архегонийлі) шөптесін өсімдіктердің бір туысы. Мұны осы күні қырықбуын деп те жүрміз. Жылбұрынның жер астында бұтақтанып өсетін тамырсабағы болады. Одан жер бетіне бұтақтанып өсетін сабақтары шығады. Сабағының буын аралығы қамыстікі сияқты қуыс болады. Жасыл түсті болатын сабағы мен өркендерінің эпидермис жасуша қабықшаларына кремнезем сіңіп қатайып кетеді де, оларға қашырлағыштық қасиет береді. Бұл қасиет инкрустация деп аталады.
Папоротник сияқты, өсіп тұрған тұтас жылбұрын да спорофит болып табылады. Бұның өркендері ұшында айырықша болып өскен масақшалары болады. Олар шоқтанып орналасқан бірнеше ұсақ-ұсақ жапырақтардан - спорофиллден - тұрады, оның пішіні алты бұрышы бар қалқанша тәрізденіп келген. Қалқаншаның сабаққа қараған төменгі жағында 8-15 шамасыңда қалталанып біткен бүртіктері болады, олар - спорангий. Олардың археспорияларындағы жасушалар редукция жолымен бөлінгеннен кейін ядроларында гаплоид санды хромосомасы бар жасыл түсті домалақ көптеген споралар пайда болады. Әрбір спораныңжасуша қабықшасы үш қабатты: оның сыртқы қабатынан гигроскопты қасиеті бар ширатылған таспа тәрізді екі өскінше жетіледі. Олар ауа ылғалды болса, спораға жабысып қалады да, ал құрғақ кезде спорадан ажырап, өрмекшінің аяқтары сияқтанып тармақтанады. Осы таспа сияқты бөлімімен құрғақ жеңіл түйіршіктерге жабысып, жел арқылы бір жерден екінші жерге барып түседі. Қолайлы ортаға түссе споралар өсіп, олардан өскінше пайда болады. Бұның түсі жасыл, үстіңгі жағы тілімденіп, телімденіп кеткен, көлемі кіші.
Әрбір телімдердің ұшында антеридий орналасады да, олардан көптеген сперматозоид пайда болады. Жылбұрынның аналық жыныс мүшесі архегоний папоротниктің архегонийіне өте ұқсас келеді де, өскіншенің көп қабатты бөлімінде жетіліп шығады. Ұрықтанған аналық жыныс жасушасынан жылбұрынның жыныссыз ұрпағы пайда болады.
Бұрын барлық жылбұрын өскіншелерін дара жынысты: ұсақтары тек антеридий, ал ірілерін архегоний деген. Кейініректе жылбұрынның кейбір түрлерінің өскіншілері қос жынысты болып келетіні анықталды. Жылбұрын өскіншілерінің дара жынысты болып көрінетін себебі: олардың екеуі бір мезеттің ішінде бірдей жетілмеуі (архегоний бұрынырақ жетіледі), сонымен қатар олардың қоректену қарқыны да бірдей болмайды.
Жылбұрынның папоротниктерден ерекшелігі - (вегативтік мүшелерін есептемеген кезде) жапырақтарына ұқсамайтындай болып спорофиллдерінің айрықша жіктелуі. Жоғары сатыдағы өсімдіктердің көбею эволюциясының бағыты: спорофилдерінің әрмен қарай өзара жіктелулері, жыныстарының морфологиялық белгілерінің өзгешеліктері, өскіншелерінің редукциялануы.
Тең споралы папоротниктердің даму циклі. Ұрпақ ауысу папоротник тәрізділерде, әсіресе аталық папоротникте жақсы байқалады.
Аталық папоротник - қоңыржай белдеудің орман аймағында өсетін көпжылдық шөптесін өсімдік. Бұлардың жер астында қуатты тамыр сабақтары, жер бетінде ірі қос қауырсын телімді жапырақтары (вайя) жетілген. Аталық папоротниктің тамырсабағынан ұлу тәрізді ширатылған өркендер өсе келе одан вегетативтік жолмен және жыныссыз, жынысты жолдармен көбейеді.
Ересек аталық папоротник - ядро фазалары тұрғысынан диплобионт (грекше «дипло» - екі есе, екі еселенген: «бионт» - тірі организм), денесін құрап тұрған жасушалардың хромосома саны диплоидты. Өсімдіктің жапырақтары толық қалыптасқаннан кейін оның астыңғы бетінде орталық жүйкені қуалай қарама-қарсы орналасқан сарғышқоңыр дөңгелек дақтар көрінеді. Олар - спорангий топтары. Спорангийдің тобы сорус, ал споралы жапырақтар спорофиллдер делінеді.
... жалғасыСіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz