Triticum durum L. қатты бидай сорттарының изоферменттерінің полиморфизмін электрофорез әдісімен анықтау

Әл-Фараби атындағы Қазақ Ұлттық Университеті
Биология факультеті
Биохимия, биотехнология және өсімдіктер физиологиясы кафедрасы
Дипломдық жұмыс
Бидай изоферменттерінің әр түрлігін электрофорез арқылы зерттеу
Орындаған: 5-курс студенті Бексеитов. Е. К
Ғылыми жетекшілері: б. ғ. к, доцент Қаржасова. А. В
ҚР ҰБО, Өсімдіктер биологиясы және биотехнологиясы институтының «өсімдіктер молекулалық генетикасы» зертханасының меңгерушісі, б. ғ. к, Әбугалиева. С. І
Норма бақылаушы Әкимбекова. А. Б
Қорғауға жіберілді:
Кафедра меңгерушісі,
Б. ғ. д, профессор Иващенко. А. Т
Алматы, 2007 ж
РЕФЕРАТ
Жұмыстың тақырыбы: Бидайдың изоферменттерінің әр түрлігін электрофорез арқылы анықтау. Жұмыс 28 беттен, 6 суреттен және 1 кестеден тұрады. Пайдаланған әдебиеттер саны - 29.
Зерттелген орны: Алматы қаласы, Өсімдіктер биологиясы және биотехнологиясы институты, «өсімдіктер молекулалық генетикасы» зертханасы.
Зерттеу нысаны: Triticum. durum L. қатты бидайдың Қазақстанға жерсіндірілген 14 сорты: Ангел, Безенчукская-139, Гордеиформе, Дамсинская-40, Дамсинская-90, Кустанайская-12, Кустанайския-52, Наурыз-2, Наурыз-3, Наурыз-4, Оренбургския-10, Омский рубин, Светлана, СИД-88.
Негізгі түйін сөздер: изоферменттер, эстераза, катодты пероксидаза, алкогольдегидрогеназа, малатдегидрогеназа, қышқылдық фосфатаза, қатты бидай T. durum.
Жұмыстың мақсаты : Қатты бидай сорттарының изоферменттік жүйелерінің полиморфизмін анықтау, сорттарда изоферменттің аллельдерінің таралу жиілігін анықтау, сол арқылы сорттарды идентификациялаудың әдістерін жетілдіру.
Зерттеу әдістері : Пісіп-жетілген дәннің белоктарын бөліп алу, центрифугалау, вертикалқы табақшада электрофорездеу, гистохимиялық бояу.
Жұмыстың міндеттері: Қазақстанға жерсіндірілген қатты бидай сорттарының генетикалық әр түрлігін зерттеу.
Зерттеу нәтижелері: глютаматдегидрогеназа, малатдегидрогеназа изоферменттерінің полиморфизмі анықталды. Зерттеу нәтижесі бойынша зерттелген сорттардың туыстық-өзгешілік дендрограммасы тұрғызылды.
Мазмұны
Кіріспе
Әдебиетке шолу . . . 5
- Изоферменттердің эволюциясы . . . 5Бір локуспен кодталатын изоферменттердің эволюциясы . . . 5Көп локуспен кодталатын изоферменттердің эволюциясы . . . 6
- Изоферменттер генетикалық маркер ретінде . . . 7
- Изоферменттер мен оларды реттегіш гендердің номенклатурасы . . . 10
- T. Бидай ферменттерінің генетикалық реттелуі . . . , . . . 10
Эстераза . . . 13
В-милаза . . . 13
Малатдегидрогеназа . . . 13
Пероксидаза . . . 14
Алкогольдегидрогеназа . . . 14
2. Материалдар мен зерттеу әдістемелері . . . 16
2. 1 Зерттеу материалдары . . . 16
2. 2 Үлгілерді дайындау және ферменттік белоктардың электрофорездік бөлінуі . . . 16
2. 3 Изоферменттерді гистохимиялық бояу . . . 17
3. Нәтижелер . . . 19
3. 1 Қатты бидай үлгілерінің энзимкодтаушы локустары бойынша салыстырмалы электрофорездік анализі . . . 19
Қорытынды. . . . 25
Пайдаланылған әдебиеттер тізімі . . . 26-28
Кіріспе
Қатты бидай Қазақстанның маңызды дәнді дақылдарының бірі болып табылады. Қатты бидай T. durum L генефондтық сорттарының идентификациялануы мен тіркеуі негізінен морфологиялық белгілері және қорлық белоктар бойынша жүргізілген. Өсімдіктердің сорттары, биотиптері және популяцияларын идентификациялауда молекулалық маркерлер эффективті (белоктық және ДНҚ-маркерлер) . Қазіргі кезде генетика және селекцияның әртүрлі мәселелерін шешуге ондаған полиморфты жүйелер пайдалануда. Изоферменттер ферменттердің генетикалық детерминацияланған көптік формасы ретінде белгілі болғаннан бергі уақытта селекция үшін өздерінің ақпараттығы, хромосомалық маркер ретінде тұрақтылығы және анықтау әдісі бойынша басқа да биохимиялық маркерлерген қарағанда қарапайым әрі олармен салыстырғанда анағүрлым нақтылығымен және тұқым қуалауы бойынша кодаминантылығымен т. б артықшылықтарымен көзге түсті. Молеклалық маркерлердің бұл класы қазіргі кезде туыстарды, түрлерді, сорттарды және линияларды идентификациялауда морфологиялық және басқа да генетикалық маркерлермен бірге кеңінен қолданысқа ие болды.
Жұмыс Қазақстан Республикасының Білім және Ғылым Министрлігі, «Ұлттық биотехнология орталығы», Өсімдіктер Биологиясы және Биотехнологиясы Институтының «Өсімдіктер молекулалық генетикасы» зертханасында жүргізілді.
1. 1 Изоферменттердің эволюциясы
1. 1. 1 Бір локуспен кодталатын изоферменттердің эволюциясы
Дарвиннің эволюциялық теориясына сәйкес түрдің генотипі табиғи сұрыпталудан өтеді, оның барысында қоршаған ортаға бейімді гендер сақталып қалады, ал бейімді еместері - бірнеше ұрпақтан кейін жойылып кетеді. Ғалымдар аллельдер варианттарының болуын бір-біріне қарама-қарсы екі көзқараспен түсіндіреді. Селекционерлер (сұрыпталу теориясының жақтаушылары) Дарвиннің теориясына сәйкес әрбір аллель түрдің өмір сүруге бәсекелестік кезінде не пайдалы не пайдасыз болады және дер берілген локуста екі альтернативті аллельдердің болуының келесідей екі себебі бар дейді. Біріншісі, полиморфизм эволюцияның аралық сатысы болып табылып, оның барысында бір аллель екінші аз пайдалы аллельдердің орның бірте-бірте басуы мүмкін; саны бойынша тұрақты популяцияда әр дарабас кем дегенде екі ұрпақ береді, егер дарабас жаңа пайда болған ген бойынша гетерозиготалы болса, ұрпағының біреуі ғана пайдалы аллельге ие болады. Осылайша пайдалы геннің таралуы - оның пайда болуынан бастап, алдындағы аллельдердің орның толығымен басқанға дейін - көптеген ұрпақтар бойы жүреді және оның барысында популяцияда екі аллель де болады. Екіншісі, плиморфизм - керісінше, реттелген болып табылады, себебі гомозиготалық жағдайда локус тек бір ғана өнім түзеді, ал гетерозиготалық жағдайда екі әртүрлі өнім түзетіндіктен локустың гетерозиготалық жағдайы гомозиготалықа қарағанда көп пайдалы болып келеді деп болжайды. Оның нәтижесінде популяцияда екі аллель де сақталып қалады. Реттелген полиморфизмнің мысалы ретінде мынадай фактіні келтіреді: адамдардың малярия таралған аймақтардағы көнеден тұрып келе жатқан тұрғындарына «орақ тәрізді» клетка анемиясы генінің Hb S сақталуы. Қалыпты генотип Hb A Hb B және «орақ тәрізді» анемия гені бойынша гомозиготалық Hb S Hb S -та бұл аймақта пайдалы емес, себебі біріншісі маляриямен ауруға бейімді, ал екіншісі тыныс алу жүйесінің патологиясы және ерте қайтыс болуға себеп болады. Ал гетерозиготалы генотип Hb А Hb S малярияға қарсы төзімділікті қамтамасыз етеді. Сәйкесінше, маляриямен ауыру қауіпі кезінде айтып кеткен екі ген бойынша гетерозиготалы генотип сөзсіз пайдалы болады. Нәтижесінде популяцияда екі ген де сақталып қалады.
Екінші көз қарасты нейтралистер айтады. Олардың айтуынша геннің құрамында болған аздаған өзгерістер міндетті түрде ол ген өнімінің қасиетіне әсер етпйді. Ол нейтралды-альтернативті аллельдер көбінесе кездесетін құбылыс және олар бейтарап болғандықтан сұрыпталуға түспейтіндігі себепті генефондта сақталып қалады деп болжайды. Әртүрлі аллельдердің кездесу жиілігі ұрпақтан-ұрпаққа өзгеруі мүмкін және олардың кездесу жиілігі Дарвиннің эволюциялық теориясына сәйкес емес, кездейсоқ гендердің дрейфі нәтижесінде болады дейді. Нейтралды мутация гипотезасына сәйкес түрдің эволюция барысындағы тарамдалуы (дивергенция) түрдің қоршаған ортаға біртіндеп бейімделуі нәтижесінде ғана емес, сонымен қатар нейтралды мутация негізіндегі болатын белоктың әр түрлігінің нәтижесінде де болады /1/.
1. 1. 2 Көп локуспен кодталатын изоферменттердің эволюциясы
Жоғарыда тек бір локустың көптік аллельдігі нәтижесінде түзілетін изоферменттер қарастырылды. Ондай аллельдер бастапқы геннің мутациясы нәтижесінде түзіледі, яғни дивергентті эволюция жолымен. Ал көптеген локуспен түзілетін изоферменттер болса - олар да дивергенция нәтижесінде түзіледі, бірақ олардың түзілуі тағы екі сатыны қажет етеді. Оны келесідей жазуға болады:
- бастапқы геннің мутациясы, көптеген аллельдердің түзілуі;
- гендік локустың екі еселенуі;
- аллельдердің сегрегациясы (бөлшектенуі) .
Бұл процесс осындай ретпен жүре ма, әлде 2-ші 1-ден бұрын бола ма белгісіз. Сұрыпталу теориясының жақтаушылары алдымен 1-ші кезең жүреді дейді. Одан әрі гетерозиготалықтың басымдығына сәйкес түрдегі аллельдердің популяцияда таралады. Содан кейін 2-ші, 3-ші сатысы жүреді, оның нәтижесінде түрдің әр дарабасы аман қалуы үшін пайдалы қасиет - әртүрлі аллельдермен кодталатын белоктардың екеуін де синтездеу қабілетіне ие болады /1, 2/.
Көптік локуспен кодталатын изоферменттердің эволюциясының 4-ші сатысы - әр локустың экспрессиялануының тәуелсіз механизмі қалыптасуы. Бұл бір ұлпада бір локус, екінші бір ұлпада басқа локус активті болып келетін көп клеткалы организмдерге тән. Оның нәтижесінде изоферменттердің ұлпалық спецификалығы пайда болады.
1. 2 Изоферменттер генетикалық маркер ретінде
Соңғы кездері селекцияның негізгі құралдарының бірі болып табылатын генетикалық маркерлер организмнің тұқым қуалаушылық конституциясын бағалауда аса маңызды роль атқарады. Генетикалық маркерлер деп геномдағы локализациясы белгілі генді айтады. Ол генді анықтай отырып басқа гендерді де анықтауға болады /2/. Алайда зерттеушілер ген-маркердің өзімен емес, оның айқын байқалатын сандық белгі ретінде көрінетін фенотиптік көрінісімен жұмыс жасайды. Бұндай белгі сәйкес генді және сонымен тіркескен басқа да гендерді анықтау факторы ретінде пайдаланыла алады.
Белокты бөліп алу мен идентификациялаудың (Электрофорез, изоэлектрофокустау, иммуноэлектрофорез т. б) қазіргі замандық әдістері молекулалардың әр түрлігі жайлы нақты мәліметтер алуа мүмкіндік береді. Ол арқылы генотиптердің әр түрлігі жайлы да мәліметтер алуға болады, себебі теориялық жағынан алғанда кез келген полипептид өзін кодтайтын геннің маркері ретінде қызмет атқара алады /3/.
Белок молекуласын зерттеудегі көптеген жетістіктерге агар, крахмал және полиакриламид гельдерінде электрофорездеу арқылы жетті. Белок молекуласының бөлінуі «молекулалық торда» тұрақты тоқ әсерінен жүреді. Бұнда «молекулалық тор» қызметін гель атқарады. Бұл кезде белоктар бір полюстен екіншісіне әр түрлі жылдамдықпен жылжиды. Олардың жылжу жылдамдығы олардың молекулалық зарядтары және көлемі арқылы анықталды. Белоктардың бөлінуі жүрген соң оларды фиксациялау және бояу жүргізіледі. Нәтижесінде зерттеліп жатқан белоктың компоненттік құрамы жайлы мәлімет беретін жолақтар спектрі түзіледі. Белоктардың суббөлікті құрамы жайлы мәлімет алу үшін электрофорезді денатурацаялйтын (мочевина, натри додесульфаты) жағдайда жүргізеді. Белоктың айтарлықтай күрделі қосылыстары жайлы нақты мәліметтер алу үшін оларды екіөлшемді электрофорездеу жүргізеді.
Электрофорез үшін көбіне гельмен толтырылған түтіктерді немесе белгілі қалындыққа ие гель пластинкаларын пайдаланады. Белоктарды олардың спектрдегі орны арқылы идентификациялайды және оларды қозғалғыштық коэффицент (Rf) көрсеткіші немесе реттік нөмірі арқылы белгілейді. Электрофорез сонымен қатар белоктың молекулалық массасы жайлы да мәліметтер алуға мүмкіндік береді. Ол үшін зерттеліп жатқан белоктың қозғалғыштығын белок-эталонның қозғалғыштығымен салыстыру жүргізеді.
Белоктардың иммунохимиялық анализі үшін антиген-антидене спецификалық әрекеттесетін реакцияны пайдаланады. Иммунохимиялық әдіс әр түрлі белоктардың биологиялық гомологиясын анықтауға мүмкіндік береді.
Негізінен электрофорез арқылы көптеген белоктар, әсіресе ферменттер көптеген формада кездесетіндігі анықталған. Соңғысы изоферменттер немесе изозим (Hounter, Markert, 1957) деген атауға ие болды. Сонымен қатар басқа да белоктардың, соның ішінде дәннің қорлық белоктарының (Jones, et. al., 1959) көптеген варианттары кездесетіндігі анықталды.
Полиморфизм термині алғаш рет 1940-жылы Форт морфологиялық өзгергіштікті белгілеу үшін қолданған. Оның анықтамасы бойынша полиморфизм - бір уақытта, бір аймақта бір түрдің екі немесе одан да көп формасының бар болуы онда болып өткен мутациялық өзгеруінің нәтижесі емес, бар аллельдер бойынша гетерозиготалықтың бар болуының нәтижесі болып табылады. Ал қазіргі кезде полиморфизм термині бір геннің аллельдің күйіне байланысты болатын индивидуалдық өзгергіштікті белгілейді /1, 3/.
Әдебиеттерде T. durum қатты бидайдың морфологиялық және биохимиялық белгілерін реттейтін гендердің локализациясы жайлы көптеген мәліметтер кездеседі.
Қазіргі кезде бидайдың селекциялық формаларының биохимиялық паспорттау кеңінен қолданылады. Селекциялық алғашқы материалдардың геномдық анализі кезінде сонымен қатар иммунохимиялық әдіс /4, 5/ және қорлық белоктар - глиадин мен глютенин анализі де қолданыс тапқан /6/. Алайда иммунохимиялық әдіс белокты бөліп алу, оған иммундық сарысу (сыворотка) алу сияқты жұмыстармен қиындатылған. Ал глиадин мен глютенинді кодтайтын локустар 1 және 6 хромосомамен шектелген. Бұл өз кезегінде шаруашылық бағалы белгілердің ген-маркермен тікелей зерттеуге мүмкіндік бермейді. Сондықтан одан да басқа биохимиялық маркерларды іздеу қажет. Ондай қызметті изоферменттер атқара алады /7, 8, 9/.
Изоферменттер деп бір түрдің дарабастарынан бөлініп алынған ферменттердің генетикалық детерминацияланған молекулалық көп формасын атайды. Олар бірдей субстратты спецификалыққа ие, бірақ бір-бірінен айырмашылығы өздерінің бірінші реттік құрылымы және физико-химиялық қасиеттері бойынша болады: электр өрісіндегі қозғалғыштығы, субстрат мен ингибитор молекулаларына туыстығы және температураға төзімділігі бойынша /10/.
Клеткадағы ферменттердің көп түрлігінің пайда болуының себептерін екі категорияға бөлуге болады:
- Гендік өзгергіштік :Гендік өнімдер in vivo будандасады:
- аллельді гендер;
- аллельді емес гендер;
- аллельді және аллельді емес гендердің өзара әрекеттесуі. Гендік өнімдер in vivo будандаспайды:
- аллельді гендер;
- аллельді емес гендер. Гендік өнімдердің in vivo будандасуы шектелген:
- Эпигенетикалық өзгергіштік:Молекуланың посттрансляциялық өзгеруі; Молекуланың iv vitro өзгеруі және артефакт /11/.
Алайда пайда болу механизміне қарамастан дәннің изоферменттік спектрі өсімдіктердің белгілі бір түр, сорт және линиялары үшін аса спецификалы болып келеді /12/.
Изоферменттерді молекулалық маркерлер ретінде пайдалану ыңғайлылығы себептері келесілер:
- Белоктар - гендік жүйенің алғашқы өнімдері және геннен белокқа дейінгі жол, геннен морфологиялық, физиологиялық белгіге дейінгі жолдан әлде қайда қысқа. Сондықтан белоктық белгі фенотиптік өзгергіштікке өз ұшырайды.
- Белок гендік жүйенің алғашқы өнімі болып табылатындықтан белок өзін кодтайтын геннің немесе гендік жүйенің (хромосома, геном) маркері ретінде пайдаланыла алады. Бұндай белок-маркерлердің жиынтығы геномның құрылымын және бүтін генотиптің спецификалығын көрсете алады /13/.
- Изоферменттерді көптеген гендер кодтайды, ал ол болса геномның барлық гомеологиялық хромосомасында орналасқан болуы мүмкін.
- Изоферменттер анализі үшін өсімдіктің аз ғана материалы қажет және де бір уақытта көптеген үлгіні анализдеуге болады.
Электрофорез әдісін пайдалану біріншіден белоктарды сәйкес көптеген гендер кластерлерімен кодталатын көп компонентті жүйе ретінде ажыратуға, екіншіден геннің аллелизмін анықтауға және әртүрлі популяциядағы геннің аллельдік құрамын бағалауға мүмкіндік береді.
Бірқатар ферменттердің изоферменттік спектрі температура, фотопериод, инфекция және өсімдіктің қоректену жағдайы сияқты көптеген факторлар әсерінен өзгергіштікке ұшырайды. Сонымен қатар, көптеген ферменттер энзимограммасы органдық, ұлпалық және субклеткалық спецификалыққа ие /14/. Сондықтан әдісті стандарттау қажет, ең алдымен фермент бөлініп алынатын, фермент көзін таңдау барысында оны ескеру қажет /9, 15/. Мұндай мақсатта ең қолайлы объект жетілген дән болып табылады /15/. Тыныштық жағдайдаға өткен дәнде ферментативтік процесстер жалғасып жатады. Оған негізінен оксидоредуктазалар мен гидролазалар қатысады. Бұл екі класс ферменттерінің негізгі қызметі - дәннің алғашқы өсу кезеңін қамтамасыз ету /16/.
1. 3 Изоферменттер мен оларды реттегіш гендердің номенклатурасы
Изоферменттердің номенклатурасы Халықаралық биохимиялық номенклатура комиссиясында (IUPAC-IUB Commission on biochemical nomenclature) құрастырылып, бекітілді /10/. Бұл номенклатура бойынша бір түрдің дарабастарынан бөлініп алынған, бірдей субстраттық активтілікке ие барлық белоктар «ферменттердің көптік молекулалық формалары» деп аталады. Ал «изофермент» немесе «изозим» терминдерімен ферменттердің тек генетикалық детерминацияланған көптік формаларын ғана айтады.
Көбіне изоферменттер мен олардың реттеуші гендерін ферменттердің көп қолданылатын атауының қысқартулары түрінді белгілейді: кодтайтын локустың бірінші әріпін үлкен, қалғандарын кіші әріптермен белгілейді. Мысалы : ADH - Adh белгілеуі ал малатдегидрогеназаға MDH - Mdh белгілеуі қолданылады. Кейде ферменттердің жүйелік атуы бойынша да қысқартылу қолданылады, мысалы MOR L-малат НАД-оксидоредуктаза, MOD L-малат: НАДФ-оксидоредуктаза. Егер фермент екі немесе одан да көп локустармен кодталатын болса, онда локустарды нөмірлейді, мысалы Mdh-1, Mdh-2 /1, 10/.
Изоферменттердің аллельдік гендері мен оларға сәйкес изоферменттерді белгілеуге әріптерді де, цифрларды да қолданады. Мысалы, Adh1-F және Adh1-S белгілеулері арқылы жылдам (fast) және баяу (slow) қозғалатын вариантын кодтайтын аллельдік гендерді белгілейді. Сонымен қатар аллельдік изоферменттер жиі аллозим деп те аталады.
1. 4 Бидай ферменттерінің генетикалық реттелуі.
Изоферменттердің тұқым қуалау табиғатын негізінен классикалық генетика әдісімен анықтайды. Ол негізінен: контрасты формаларды будандастыру, анализдік шағылыстырудың F 2 ұрпағындағы белгілердің ажырауын анализдеу. Сонымен қатар қалыпты және анеуплойдты (бір хромосомасы артық немесе бір хромосомасы кем) формалардың изоферменттік спектрін салыстыру арқылы анықтауға болады. Гетеромультимерлі молекула түзе алатын ферменттер үшін сомалық ұлпа мен тозаңқаптың изоферменттік спектрін салыстыру бұл спектрдің сәйкес ферментті реттеуші локустын гетерозиготалық жағдайымен ба, жоқ әлде аллельді емес ген активтілігі нәтижесінде пайда болды ма анықтауға мүмкіндік береді /17/. Харт және Ленгстон (Hard, Lengston, 1977) T. diccocum-ның алкогольдегидрогеназа (ADH) ферментін электрофорездеу арқылы бұл ферменттің димер екенін анықтады: ол α - және β-суббөліктен тұрады. Және де бұл ферменттің электрофорездік гельдегі үш варианты ADH-1, ADH-2 және ADH-3 екі суббөліктің әртүрлі ассосациясы нәтижесінде түзелетіні анықталды. Чайниз Спринг (T. aestivium) нули-тетрасомды линиялары көмегімен суббөліктерін кодтайтын локустарын локализациялау жүргізілді. Осылайша құрылымдық гендері 4A, 4B және 4D хромосомаларының β-иығында орналасқаны анықталды. Қатты бидайдың (T. durum) ADH-1 изоферментін кодтайтын Ahd-1 локусы да 4А хромосомасында қысқасабақтық генімен Gai (Rht-1) бірге орналасқаны анықталды.
Бидайдың α-амилаза ферменті де ең алғаш зерттелген бидай изоферменттерінің бірі. Көптеген аймақтар үшін өте маңызды белгі - дәннің тамырда өсуі - α-амилаза активтілігіне байланысты. Дәннің өнуі кезінде түзілетін изоферменттердің алтыншы және жетінші гомеологиялық хромосомаларымен байланыстылығы анықталған. Харт 6-шы хромосомада орналасқан локусты α- Ahd-1 деп, ал жетінші гомеологиялық топтың хромосомасындағы локусты α- Ahd-2 деп белгілеуді ұсынды. Марчило және оның әріптестері (Marchylo et al, 1980) α-амилазаның екі типін идентификациялады: 1) дәннің тізілуі және жетілуі кезінде активті болатын G1 және G2, 2) дәннің өнуі кезінде активті болатын G3. Пісіп-жетілген дәннің өнуі кезінде анықталатын екінші тип изоферментті (α-AMY2) α-Amy-2 локусымен кодталады. Бұл локус 7-ші гомеологиялық топ хромосомасының ұзын иығында орналасқан. Гибридологиялық анализ көмегімен α- AmyВ2 күрделі локусы карталанған ( қабылданған номенклатура бойынша берілген локус бидайдың хромосомаларының гомеологиялық тобының қайсысына жататының белгілеу үшін локустың белгісіне A, B және D әріптері қосылады) (сурет-1) /3/.
30 44 6 13 11 26
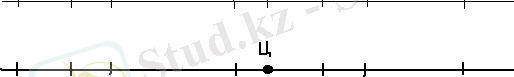
Lr14a Sr17 Pm5 α- AmyВ2 Pc Vrn Vrn5
Сурет1.
α- AmyВ2 геннің бидайдың 7-ші хромосомасындағы локализациясы. Цифрлармен рекомбинация бірлігі көрсетілген. Ц-центромера; Lr14a, Sr17, Pm5, α- AmyВ2, Pc, Vrn және Vrn5 басқа гендер (Marchylo et al, 1980) /3/.
Осылайша анықталған басқа да бидай изоферменттерінің кейбірінің локализациясы 1-ші кестеде көрсетілген.
Кесте 1.
Бидайдың идентификацияланған изоферменттерінің хромосомадағы локализациясы (Созинов А. А, 1985) /3/.
... жалғасыСіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz