Fusarium solani саңырауқұлағымен залалданған картоптағы Са2+-АТФ-аза ферментінің қорғаныштық рөлі

ӘЛ-ФАРАБИ АТЫНДАҒЫ ҚАЗАҚ ҰЛТТЫҚ УНИВЕРСИТЕТІ
БИОЛОГИЯ ФАКУЛЬТЕТІ
Биохимия, биотехнология және өсімдіктер физиология кафедрасы
БІТІРУ ЖҰМЫСЫ
Саңырауқұлақпен залалданған картоптың қорғаныштық реакцияларына Са 2+ -АТФ-аза ферментінің қатысы
Орындаушы:
IV курс студенті Маллахожаева Г. С.
Ғылыми жетекші:
Б. ғ. к., профессор:Валиханова Г. Ж.
Б. ғ. к:Манадилова А. М.
Норма бақылаушысы:Әкімбекова А. Б.
Қорғауға кафедра
меңгерушісімен жіберілді:Иващенко Т. А.
б. ғ. д., профессор
Алматы 2007
РЕФЕРАТ
Бұл жұмыс М. А. Айтхожин атындағы молекулалық биология және биохимия инстетунда табиғи қосылыстар знзималогиясы лабораториясында орындалды.
Бітіру жұмысқа реферат, мазмұны, кіріспе, әдебиетке шолу, объектілермен мен әдістемемелер, нәтижелер және оларды талқылау, қорытынды, пайданылған әдебиеттер тізімі, 5 сурет кіреді.
МАЗМҰНЫ
КІРІСПЕ
1. ӘДЕБИЕТКЕ ШОЛУ
1. 1. Са 2+ -АТФ-азаның өсімдік клеткасындағы маңызы 5
1 . 2. Са 2+ - АТФ-аза ферментіне жалпы сипаттама беру 6
1. 3. Fusarium solani саңырауқұлағының жалпы сипаттамасы 7
2. ЗЕРТТЕУ ОБЪЕКТІЛЕРІ МЕН ӘДІСТЕМЕЛЕРІ
2. 1. Зерттеу объектілері 10
2. 2. Зерттеу әдістері 10
2. 1. 1. Сұйық ортада картоптың каллус клеткаларын өсіру 10
2. 2. 2. Fusarium solani саңырауқұлағын өсіру 11
2. 2. 3. Fusarium solani саңырауқұлағымен картоп түйіндерін
және сұйық ортада өсірілген каллус клеткаларын залалдау 12
2. 2. 4. Са 2+ - АТФ-азаның активтілігін анықтау 12
2. 2. 5. Са 2+ - АТФ-азаға әсер ететін факторларды анықтау
әдістері 13
3. ЗЕРТТЕУ НӘТИЖЕЛЕРІ МЕН ОЛАРДЫ ТАЛҚЫЛАУ
3. 1. Са 2+ - АТФ-азаның активтілігін е Са 2+ иондары концентрациясының
және рН деңгейінің әсері 14
3. 2 . Картоптың каллус клеткаларын Fusarium solani саңырауқұлағы
залалдағанда Са 2+ - АТФ-азаның активтілігіне әсері 17
3. 3. Fusarium solani саңырауқұлағымен залалданған картоп
түйіндерінде
Са 2+ - АТФ-азаның активтілігінің өзгеруі
ҚОРЫТЫНДЫ
ПАЙДАНЫЛҒАН ӘДЕБИЕТТЕР ТІЗІМІ
КІРІСПЕ
Табиғаттағы өсімдіктер әртүрлі микроорганизмдердің арқасында залалданып қауіпке ұшырайды. Сол сияқты, қазіргі кезде фузариоз проблемасы ауылшаруашылық өсімдіктері үшін өте маңызды болып саналады.
Олар әртүрлі топырақта кездесетін спора түрінде сақталады. Ең көп тараған және қатерлі Fusarium solani саңырауқұлағы болып табылады. Негізінен инфекцмя споралары бар топырақ арқылы жұғады. Бұл ауру қыстық-көктемдік кезңдегі картоп шығынының негізгі себебі болып саналады. Осы ауру әсерінен сақтау кезеңінде шығын 20% дейін жетеді. Фузариоздық аурудың зияндық әсері картоп түйіндерінің шығынымен ғана емес, инфекцияланған түйіндерді себу кезінде өсімдіктердің өсуі тежеліп және ол өз кезегінде түсімінің азаюына себеп болады. Ал инфекцияланған түйіндердің біреулері өспей, дамымай қалады.
Осы мәселені шешудің бір көзі болып Fusarium туысына жататын саңырауқұлақтардың әсеріне қарсы тұра алатын өсімдіктердің метаболизмін зерттеу табылады. Осыған байланысты көптеген ғалымдар патоген мен өсімдік арасындағы қатынасты зерттеуге көңіл аударуда.
Сонымен қатар картоптың патогенге тәуелді Са 2+ -АТФ-аза ферментін зерттеу маңызды болып отыр. Бұл фермент картоп клеткасының әртүрлі қорғаныштық реакцияларына қатысады.
Бұл жұмыстың мақсаты: Fusarium solani саңырауқұлағымен залалданған картоп клеткаларында Са 2+ -АТФ-аза ферментінің қатысын зерттеу.
Мақсатқа байланысты туатын міндеттер:
- Каллус клеткаларын агарланған қоректік ортада өсіру.
- Картоп клеткаларын сұйық ортада өсіру.
- Fusarium solani саңырауқұлағын өсіріп, әртүрлі картоп сорттарын залалдау.
- Fusarium solani саңырауқұлағымен залалдаған картоп түйіндеріндегі және сұық отрада өсірілген клеткалардағы Са2+-АТФ-аза ферментінің активтілігін анықтау.
I. ӘДЕБИЕТТЕРГЕ ШОЛУ
1. 1. Са 2+ -АТФ-азаның өсімдік клеткасындағы маңызы
Табиғатта өсімдіктер көптеген патогенді микроорганизмдердің арқасында қауіптілікке ұшырайды, бірақ соның өзінде көпшілік түріне қарсы тұра алады. Патогенді микроорганизмдерге қарсы жауап ретінде өсімдіктер төтеп бере әртүрлі индуцияланған төзімділік механизмдерін қосады. Осылардың қатарында: оттегінің түзілуі, клетка қабығының қалындау (каллоза мен лигниннің бөлінуі) фитоалексиндер синтезі, программаланған клеткалардың өлуі, яғни жоғары сезімтал реакциясы гидролитикалық және транспорттық ферменттердің босалуы. Төзімділік процесінде плазмалық мембрананың ең негізгі ферменті Са 2+ -АТФ-аза маңызды роль атқарады.
Cа 2+ иондары өсімдік клеткаларында қалыпты жағдайында және әртүрлі стрестік факторлар әсер еткенде (температураның төмендеуі, қуаншылық, патогенез) реттегіш қызмет атқарады. . /1/
Cа 2+ ионы өсімдіктің өсу және даму процесінде маңызды роль атқаратын бұрыннан белгілі. Cа 2+ ионы Allium сера in vitro өсірілген клеткаларында фитоалексиндер мен каллозалардың синтездейтін индукциясының екіншілік мессенджерлік қызмет атқаратыны көрсетілген /2/. Cа 2+ мүмкін клеткаларды элиситормен өңдегенде бастапқы кезеңінде клетка ішінде Cа 2+ концентрациясы өсіп фитоиммундық жауапқа жағдай туғызады /3/. Өсімдік клеткасындағы Cа 2+ ионының концентрациясының қалыпты жағдайда 10 -7 -М шамасында болады. Соның сақталуы гомеостаздың өзгермеуіне жағдай туғызады. Кальций концентрациясының цитоплазмада тұрақтылығы Cа 2+ -ді тасымалдайтын жүйесі арқасында жүзеге асады, ол артық иондардың клетканың органеллаларына немесе клетка қабығына ауысуына тікелей қатысады. Осы процесте плазммалеммамен, тонопластпен, эндоплазмалық ретикулуммен байланысқан Cа 2+ -ді тасмалдайтын (Са 2+ АТФ-азалар) маңызды роль атқарады/4/. Минорскийдің гипотезасы бойынша Cа 2+ ионының көбеюі суықтың нәтижесінде орын алады. Сонымен қатар, өсімдікке әртүрлі факторлар әсер еткенде Cа 2+ ионының маңызды роль атқарады. Мысалы: 2˚ С температурада Cа 2+ ионының жүгері тамырына сіңірілуі 20-25%- ға өседі. Жылу сүйгіш өсімдіктерде төмен температураның әсері кальций бос иондалған формасында цитоплазмада көбеюі анықталған, яғни Cа 2+ ионының компартментациясның өзгеруіне әкеледі. Нәтижесінде цитоплазманың қозғалысына және субклеткалық құрылысына әсер етеді. Кальцийдің құрамына байланысты өсімдіктер: кальциофильді, кальциофобты және нейтралды болып бөлінеді. Бұршақ тұқымдасты, күнбағыс, картоп, капуста өсімдіктерінде кальцийдің құрамы көп. Кальцийдің жетіспеушілігінен бірінші кезекте жас меристемалық ұлпалар және тамыр жүйесі зардап шегеді. Мұндай кезде мембрананың саңылауларының жұмысы нашарлайды, мембрананың транспорттау процесі бұзылады, клетканың бөлінуі, созылуы және тамырдың жетілуі тежеледі. Нәтижесінде жемістерде некроз пайда болады. Жапырақтардың түсі өзгеріп, бірінші ұштары ағарып кейін қараяды. Бірақ кальцийдің жетіспеушілігі аз байқалады. Себебі топырақ кальцийге бай.
Иондық баланста кальцийдің ролі өте күшті. Cа 2+ иондары өсімдік клеткаларын ионмен қамтамасыз ету барысында реттегіштік маңызы өте күшті.
Кальцийдің цитоплазмада реттелуі Cа 2+ - каналы және Са 2+ - АТФ-азаның активті транспортына байланысты болады. Кальций клеткалық сигнализация процесі кезінде екіншілік делдалдық қызмет атқарады. 1980 жылы Г. Расмуссеном Cа 2+ ионың екіншілік месенджерлік қызмет атқаратыны туралы айтты. /1/ Өсімдіктерде Cа - модульдейтін белоктар болып кальмодулин болып табылады.
1. 2. Са 2+ - АТФ-аза ферментіне жалпы сипатама беру
Клетка цитоплазмасындағы кальций иондарының концетрациясы 50-100 нм (5·10 -8 -1·10 -7 М) . Осы концентрацияны кальций насосы- Са 2+ - АТФ-аза ферменті тасмалдайды. Са 2+ - АТФ-аза ферменті клеткада орналасуына, құрылысына және реттегіш қабілеттілігіне байланысты бірнеше топқа бөлінеді. Бірақ бұл ферментердің барлығы кальций иондарын клетка ішілік сұйықтықтан клетканың сыртқы сұйықтығына тасмалдайды. АТФаза гидролизіндегі энерияны пайдалана отырып цитоплазмада кальций ионының төменгі концентрациясын сақтайды. Бұл процесс клеткалық мембранадан кальций иондрының өткізгіштігін реттейді және клеткаға кіріп, бұл иондар клетка ішілік процестердің активтенуіне әсер етеді. Са 2+ - АТФ-азалар цитоплазмалық мебрананың және клетка ішілік мембрана құрамына еніп қасиетіне байланысты
өзара бөлінеді. Са 2+ - АТФ -аза мономерлі белок болып табылады, яғни бір полипептидтік тізбектен құралады, бірақ молекулалық салмағына байанысты бір-бірінен айырмашылығы бар. Саркоплазмалық ретикулум Са 2+ - АТФ-аза ферментінің молекулалық массасы 108 кД, ал плазмалық Са 2+ - АТФ-аза ферментінің молекулалық массасы 120 кД.
Кальций ионының транспорты тік ішек клеткасында және бұлшақет ұлпаларында жақсы зерттелген. Са 2+ - АТФ-азаның әсер ету механизмі:
Е 1 +АТФ↔︎ Е 1 -АТФ
Е 1 -АТФ+2Са 2+ ↔︎ Е 1 +АТФ- Са 2
Е 1 -АТФ- Са 2 ↔︎ Е 1 ~Р- Са 2 +АДФ
Е 2 ~Р- Са 2 ↔︎ Е 1 -Р- Са 2
Мg
Е 2 ~Р~Са 2 ↔︎ Е 1 -фосфат- 2Са 2+
E 2 ↔︎ Е 1
Е 1 - микросомның сыртқы бетінде орналасқан
E 2 микросоманың ішкі жағында орналасқан. Са 2+ - ға тәуелді АТФазаның активациясы үшін фосфолипидтер мен магний иондары қажет. Бірақ магний соңғы кезеңінде тікелей фосфатазды реация үшін қажет.
1. 3. Fusarium туысына сипаттама
Fusarium туысындағы саңырауқұлақтар табиғатына кең таралған және олардың көбі сапрофитті болып келеді. Бірақ кейбір жағдайда олар паразитті болуы мүмкін. Олар көбіне ауыл шаруашылық өсімдіктерін зақымдайды /1/.
Топырақ Fusarium туысына жататын түрлердің табиғи таралған ортасы болып саналады /1/. Топырақта Fusarium активті формада болады, өсімдіктің әр түрлі қалдықтарында, ризосферада және тамырдың үстіңгі жағында өседі. Бұл түрге жататын саңырауқұлақтар ішінде ризосфералық және тамырмекенділер көп кездеседі. Өсімдіксіз топырақта олардың саны өте аз, себебі олар басқа саңырауқұлақтар және бактериялардың бөлген антибиотиктері мен литикалық ферменттеріне өте сезімтал келеді.
Олар қоректік орта, яғни көмірсутектің көп мөлшері бар жерде жақсы өседі. Бұл саңырауқұлақтар факультативті анаэробты болып келеді. Көптеген түрлері pH мағынасының үлкен ауытқуларында да өсе алады. Сонымен қатар олар өсу барысында ортаның pH - ын өзгертуі қышқылдатуы немесе сілтіденуі мүмкін. Фузариум мезофильді саңырауқұлақтары болып есептеледі, оптимум температура 18-24-28С 0 . Ал температураның және рН - ың орталық көрсеткіштері шекті мағыналарында спора түзу және кейбір арнайы метаболиттердің синтезі жүреді.
Конидиялық спора түзу Fusarium түрлерінде морфологиялық және спораны түзу әдістері жағынан әр түрлі болады. Макрокондиялардың морфологиялық сәйкестік белгісі оның бір немесе екі ұшындағы сүйірленіп келуінде, аяқшалардың болуында, төменгі бөлігіндегі сарғыш секілді өсіндінің болуында.
Макроплидиялардың спорадохияларда және плонноттарда, мейелилерде, конидияносецтерде түзілуі мүмкін. Fusarium түрінің кейбіреулерінде канидиялар кәртейген кезде бөлек клеткаларға бөлініп, клетка қабығы қалыңдаған хламидоспоралар түзеді. Орта жағдай оптимальды болғанда конидиялық спара түзу үлкен қарқынмен жүреді. Бұл түрдің сәйкес ортаға көптеп көбеюіне алып келеді.
Макроконидиялар элипс тәріздес бүгілген болып келеді. Көптеген түрлеріне сәйкестік қасиет - жоғарғы клетканың біркелкі жіңішкеруі. /7/
Әр түрге жататын макроконидиялар бөліктерінің саны 3 тен 5, сирек 5 тен 7 дейін болуы мүмкін. Макрокомидиялар мен гидатердің сырты көбіне боялмаған болып келеді.
Сонымен қатар көбею және өмірін сақтап қалу маңыздылығында Fusarium кейбір түрлерінде микроконидиялар түзіледі. Олардың бөліну саны 0-1 болады.
Фузарий мицелилері де көп түрлі болып келеді. Оның дифференциясы әр түрлерде, өсіру әдісіне және өсіру жасына байланысты болады. /7/
Жоғарыда айтылғандай, бұл саңырауқұлақ фузариоз ауруын тудырады, яғни өсімдіктің клеткаларының құрылысын немесе сол клеткаға әсер ететін токсиндер, сол сияқты клетканың ядролық ақпаратын бұзады. Соның ішінде клетка құрылымын бұзып, яғни өсімдік клеткасының мембранасын ерітіп, ішке кіргеннен ауру туғызады. Осындай механизммен зақымдайтын Fusarium solani болып келеді. Ол клетка қабығын бұзу үшін көптеген ферменттер синтездейді. Мысалы: пектиназа, т. б
Fusarium solani конидиялары.
.
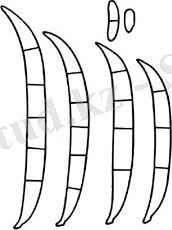
1 сурет
Fusarium solani туғызатын шірік . Саңырауқұлақтар тудыратын аурулар өсімдіктер және картоп түйінддерінде кең тарадған. Картоптың ауру тудырғыштарының түрлік құрамы, таралуы және зияндылығы, топырақтық - климаттық ерекшеліктерге, агротехниканың деңгейіне және картопты өңдеу технологиясына байланысты болады. Ауаның және топырақтың жазғы жоғарғы температурасы, ылғалдылықтың жеткіліксіз болуы, әлсіз желдер, ауаның және топырақтың температурасының кенеттен түсуі, саңырауқұлақтар тудыратын аурулардың дамуына себеп болады. Оған фитофториоз, ризоктониоз, жәй порша, құрғақ шірік және олардың аралас инфекциялары жатады /1/.
Құрғақ фузориялық шірік - ең көп таралған және зиянды картоп ауруы. Ол сақтау негізінде дамиды. Құрғақ шірік тудырушылары тілімдер мен жаралардан енеді. Әсіресе ол механикалық жинауда және сақтау орнындағы температураның тез түсуі кезінде көп болады. Ауру түйіндерде қоңырлау - ашық дақ, кішкене ішке қарай түскеннен басталып және түйіндердің жабынды ұлпасының әжімделуімен жалғасады. Түйіндегі дақтың астындағы жемісті бөлік үгімшелі, құрғақ және қышқыл қоңыр түсті болады. Зақымданған ұлпада бос кеңістік пайда болып, ақ, сары және қызыл түсті саңырауқұлақ мицелилерімен толады. Ылғалды болғанда түйін сулы консистенцияға ие болады. Ал құрғақ жағдайда түйіндер әжімделген қабықпен жабылған мумияға айналады. /1/
Сақтау кезіндегі құрғақ шіріктен шығатын шығын 15-30 % болады. Ауру түйіндер өсудің баяулауы және вегетациясы кезіндегі алдын ала салудың себебі болып саналады. Қорытындысында өнім алу 12 % және одан да көпке төмендейді. Ауру түйіндерінің ұрпақтары сырттай сау болса да, оларды сақтауда құрғақ шірікті түйіндер пайызы жоғары болып келеді.
Соған байланысты көптеген ғалымдар өсімдік және патоген арасындағы әсерлесудің молекулалық механизміне көп көңіл бөлуде. Сонымен қатар көптеген ферменттік жүйелердің активтенуімен, соның ішінде Са 2+ - АТФ-аза ферментінің активтенуін зерттейді.
2. ЗЕРТТЕУ ОБЪКТІЛЕРІ ЖӘНЕ ӘДІСТЕРІ
2. 1. Зерттеу объектілері
Зерттеу объектісі ретінде Тамаша және Санта картоп сорттарының каллус клеталары мен картоп түйіндері алынды. Тамаша Fusarium solani саңырауқұлағына төзімді сорт, ал Санта сезімтал сорт болып табылады. Каллус клеткалары агарланған және сұйық қоректік орталарында өсірілді. Сонымен қартар, зерттеу объектісі ретінде Fusarium solani саңырауқұлағы өсіріліп пайдаланылды.
- Зерттеу әдістері
2. 2. 1. Каллус клеткаларын алу және өсіру .
Мурасиге-Скуг қатты агарланған қоректік ортасы(г/л)
Макро және микро тұздары (орта Мурасига Скуг дайын ұнтақ) 4, 4 г.
Сахароза 30 г, Казеин гидролизаты 2 г, Мезоинозит ерітіндісі 10 мл, Гормондар (2, 4 Д) және кинетин, Агар 7, 2 г.
Каллус клеткаларын шығару үшін эксплант ретінде пробиркалық екі картоп сортының өсімдіктерінің сабақ кесінділері алынды да, агарланған Мурасиге- Скуг қоректік ортасы құйылған Петри табақшаларына отырғызылды. Жарық бөлмеде температурасы 25-27˚С өсірілді.
Каллус клеткаларын пробиркалық картоп өсімдіктерінің сабағынан кесіп алып, петри табақшасындағы агарланған қоректік ортаға отырғызамыз. Тамаша сорты 2, 4Д (0, 8 мл) гормоны бар агарланған қоретік ортада өседі. Ал Санта сорты Д 1 к-0, 5 (кинетин 1мл) гормоны бар қоректік ортада өсіріледі. Бұл клеткаларға жарық жақсы түсуіне байланысты қарқынды өседі. Әр апта сайын жаңа қоректік ортаға егіп отыру қажет.
Сұйық ортада клеткаларды өсіру.
Каллусогенез процесі жүріп өсіп шыққан, каллустар Мурасиге-Скуг сұйық қоректік ортасына әр апта сайын отырғызылды. Сұйық қоректік орта Мурасиге-Скуг бойынша дайындалды, агар қосылған жоқ.
Тәжірибе үшін бір күн бұрын жаңа отырызылған клетканы пайдаланамыз Клеткалар 250 мл колбаларда өсіріледі. Колбаға шамамен 50 мл орта құйылып, ішіне каллусты салынып, оны шайқауыштың үстінде, жарығы бар бөлмеде өсіріледі. Клеткаларды заттық әйнекке салып, фуксин бояуымен бояп, микроскоптың астында көріп оларды бағалайды. Боялған клеткалар өлі клеткалар деп есептелінеді, себебі өлген клеткалардың цитоплазмалық жүйесіне бұзылған мембрана арқылы бояу өтіп, клеткаларды бояйды. Ал тірі клеткалар болып боялмайды. Сонымен қатар, басқа микроорганизмдермен немесе бактериялармен залалданбағанын тексереміз. Клеткалар ашық түсті немесе хлорофил арқасында жасыл түсті болуы тиіс.
2. 2. 2. Fusarium solani саңырауқұлағын өсіру.
Fusarium solani саңырауқұлағы үшін Чапектің агарланған қоректік ортасы дайындалды.
Ортаның құрамы (1 литрге) :
Сахароза 30 г
NaNO 3 2 г
KH 2 PO 4 1 г
MgSO 4 0, 76 г
Kcl 0, 5 г
FeSO 4 0, 5 г
Чапек агары 20 г
Картоп экстраты 100-150 г
Тұздардың барлығын өлшеп алып агарды қосып қайнауға деін жеткізіп, содан кейін картоп экстраты құйылады. Бұдан кейін 25 мин стерилдейді. Стерильді пробиркаларды жатқыза орналастырып, дайын болған орта құйылады. Конидиялары өсіп шыққан пробиркалардан ілмек арқылы саңырауқұлақ мицелиилерін алынып, зигзак бағытымен ортаға енгізіледі. Конидийлер термостатта 20-25˚ С температурада және қараңғы жерде өседі. Конидийлер бір апта көлемінде өсіп шығады, мицелий алу үшін бір ай өсіріледі.
Ал оны ұзақ сақтау үшін тоңазытқышта ұстауға болады. Себебі суық жерде саңырауқұлақтың өсуі тежеледі. Осындағы патогенді саңырауқұлақ қызғылт түсті береді.
2. 2. 3. Fusarium solani саңырауқұлағымен картоп түйіндерін және сұйық ортада өсірілген каллус клеткаларды залалдау.
Картоптың екі картопт сорты алынды, олардың бірі-Тамаша Fusarium solani саңырауқұлағына төзімді, ал Санта сезімтал болып келеді.
Картоп түйіндерін 15 мин калий перманганатының 5% ерітіндісінде ұстап, шайылады және 10 мин стерилизация үшін натрий гипохлоридінде ұсталынады ( 0, 1% NaClO, 0, 1% Tween) . Түйіндердің сыртын тазалап, ламинарлы бокста картоп түйіндерін кесіп, Петри табақшасына 4 данадан кесіп салынады. Петри табақшасының түбінде фильртлі қағаз төселінді және оның үстіне 2 мл стерильденген құбыр суы құйылды. Бұл кесінділер кеуіп кетпеуі үшін. Картоп түйіндерін 1, 5 мл Fusarium solani саңырауқұлағының фильтратымен залалдандырылды. 4 Петри табақшасының әр біреуіне төрт кесінді орналастырылды. Екі табақшада Тамаша сортынан алынған үлгілері бақылау мен тәжірибе үшін. Олардың қалған екеуі Санта сортының бақылау және тәжірибе үлгілері. Бақылау үшін табақшаға 1, 5 мл құбыр суы қолданылды. Залалданған картоп түйіндері 24 сағаттан кейін қарайып түсін өзгертеді. Ал сұйық ортадағы картоп клеткаларын залалдау кезінде бақылау және тәжірибе үшін екі колба алынып, бақылау колбасына тек 5 мл стерилденген құбыр суы құйылды да, ал тәжірибе колбасына 5 мл Fusarium solani саңырауқұлағының фильтраты құйылды.
2. 2. 4. Са 2+ - АТФ-азаның активтілігін анықтау.
Картоп түйіндерінен 2 г кесіп алынып, 10 мл ортасында құрамы (0, 1 M сахароза 50 mM Трис-HCl буферінде рH 7, 4) гомогенезделіп, сол гомогенаттан 1 мл алынып ол инкубация ортасына қосылды. Инкубация ортасының құрамы: 50 мМ трис- Нсl буфері рH 7, 4, 0, 5 мМ АТФ және 2, 25 мМ Сасl 2 , Ерітіндінің көлем барлық жерде 3 вариантта да 4 мл құрайды. 30 мин 38˚С температурада инкубацияланады да, 0, 5 мл 30 % - ды үшхлорсірке қышқылымен реакция тоқтатылады. Осы процестен кейін минутына 1 айналымында/5мин центрифугаға қойылады. Одан 2 мл гомогенаты бөлініп алынып, 2 мл 1М натрий ацетаты, 4 мл 0, 025М буферлі ацетат және 1 мл аскорбин қышқылы қосылады. Осы қоспаға 10 минуттан соң 1 мл аммоний молибдаты қосылады. Содан 10 мин ұсталынып, спектрофотометрде 720 нм толқын ұзындығында тығыздығы өлшенеді. Реакциялық қоспа фильтрден өткізкізіліп, 0, 5 мл фильтраттан бейорганикалық фосфор анықталынады /5/.
Зерттеліп отырған экстракттан 2 мл алынып, оған 2 мл натрий ацетатын және 4 мл (рН=4) ацетатты буфері, 1 мл аскорбин қышқылы қосылады. 10 мин өткеннен кейін ерітіндіге 1 мл аммоний молибдаты қосылады. 10 минуттан кейін спектрофотометрде 660 нм толқын ұзындығында фосфордың тығыздығы анықталынады. Жалпы белоктың құрамын микробиурет әдісімен /6/ анықталынды. 50 мкл гомогенатты пробиркаға құйып, үстіне 2 мл дейін су құйылады. Оның үстіне 2 мл 6 % натрий гидрооксиді және 0, 2 мл Бенедикт реактиві құйылып, аралыстыралады. Бөлме температурасында 15 минут қалдырылып, кейін 330 нм толқынды спектрофотометрде өлшенеді.
Сұйық ортада каллус клеткаларын залалдау үшін стерильді ортада алдын ала өсірілген суспензияға Fusarium solani саңырауқұлағының қоректік орта фильтратын 1 ׃ 10 қатнасында қосылды. . Бұл жұмысқа пассаждан кейінгі бір күндік клеткалар суспензиясы қолднды. Уақыт өткенне кейін айтылған әдістеме бойынша Са 2+ - АТФ-аза ферментінің активтілігі анықталды.
2 . 2. 5. Са 2+ - АТФ-азаға әсер ететін факторларды анықтау әдістері.
Әсер ететін факторлар ретінде кальций концентрациясы, рН деңгейі және ингибиторлар әсері болады. Са 2+ - ионының концентрациясын анықтау үшін концентрациясы қолданылды: 2, 25 мМ СаСl 2 Бұл Лукаткин, А. С. Еремкин әдісі бойынша анықталынды. Рн және ингибиторлар әсері де осы әдіспен анықталды. Ингибитор ретінде ЭГТА () қолданылды. РН деңгейін анықтау үшін мына көрсеткіштері пайдаланылды: 5, 0. 6, 0. 7, 0. 8, 0. 9, 0.
3. ЗЕРТТЕУ НӘТИЖЕЛЕРІ МЕН ОЛАРДЫ ТАЛҚЫЛАУ
3. 1 Са 2+ - АТФ-азасына Са 2+ иондары концентрациясының және рН деңгейінің әсері
... жалғасыСіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz