Майлардың қорытылуы мен сіңірілуі, деполануы және мобилизациясының гормональдық реттелуі

Жоспары:
- Майлардың қорытылуы мен сіңірілуі
- Майлардың деполануы мен мобилизациясы
- Майлардың мобилизациясы (липолиз) . Майлар мобилизациясының гормональдық реттелуі.
Майлардың қорытылуы мен сіңірілуі
Майлардың қорытылуы - бұл панкреаттық липаза ферменті әсерінен майлардың гидролиздену процесі. Ересек адамдарда майлардың қорытылуы тек ішекте келесі схема бойынша өтеді: «өттің бөлінуі майлардың эмульгирленуі панкреаттық липазаның әсері». Бұл ферменттің әсері үшін келесі жағдайлар қажет: бейтарапқа жақын рН мәні, колипаза белогы. Колипаза панкреаттық липазамен бірге, ұйқы безінде синтезделіп, секреттеледі. Қажетті рН мәні (асқазаннан түскен қышқылдың құрамы) ұйқы безінен бөлінетін бикарбонатпен бейтараптанады.
Липидтердің ыдырауы 12 елі ішекте өтеді. Ол ұйқы безі сөлінен түсетін липаза және өт құрамындағы коньюгирленген өт қышқылдары қатысуымен атқарылады.
Өт. Өт - бұл жасыл-сары түсті, бауыр жасушаларында өндірілетін мицеллалар компоненті құрамды, тұтқыр сұйық. Өт бауырдағы өт қапшығында жиналады. Өттің маңызды компоенттері: өт қышқылдары - 6, 2%, өт пигменттері (гем ыдырау өнімдері) - 3, 2%, холестерин - 0, 5%, фосфолипидтер - 0, 5%.
Өттің негізгі қызметі - липидтердің қорытылуы мен сіңірілуіне қатысуы. Өт қышқылдары майларды эмульгирлейді және сондықтан панкреаттық липазаның майларының гидролизін атқаруына мүмкіндік береді. Өттің секреттелуі ағзадан холестериннің артық мөлшерінің шығарылуының негізгі жолы болып табылады. Ол өт қышқылдары (холестериннен бауырда синтезделетін) түрінде де, бос холестерин түрінде де шығарылады. Холестерин гирофобтық зат, сондықтан оны өт қышқылдары өт мицеллалары түрінде ерітілген қалыпта ұсталыып тұрады. Өт қышқылдары мен холестериннің қатынасы 12:1 жуық болады.
Бауырда түзілетін холь және хенодезоксихоль қышқылдары бірінші ретті өт қышқылдары деп аталады. Олар глицин және тауринмен эфирленіп, жұп немесе конъюгирленген өт қышқылына айналады да, сол формада секреттеледі. Бұл олардың гидрофильділігінің артуына әкеледі. Гликохоль қышқылының гидрофильдік бөлігі глицин қалдығы, таурахоль қышқылынікі таурин қалдығы болып табылады. Гидрофобтық бөлігі холестериннің туындылары болып табылады.
Өт қышқылдары синтезінің негізгі реттеуші ферменті, бұл 7-α-гидроксилаза. Оның активтілігі фосфорлану және дефосфорлану, фермент мөлшерінің өзгерістері арқылы реттеледі. Холестерин бұл ферменттің генінің транскрипциясын индуцирлейді, өт қышқылдары оны тежейді. Бұл ферменттің синтезінің активаторына қалқанша безінің Т 3 және Т 4 гормондары, ал корепрессоры эстрогендер болып табылады. Бұл құбылыс әйелдердегі өт тасы ауруларының ерлерге қарағанда, 3-4 есе жиі кездесетінін түсіндіре алады.
Ішектерде конъюгирленген өт қышқылдары таурин мен глицинді жоғалтып, екінші ретті өт қышқылдарына дезокихоль, литохольға айналады. Адамда ішекке түсетін өт қышқылдарының 95%-ы ішектер арқылы сіңіріліп, қанмен қайтадан бауырға түседі де, өт қапшығына жиналады. Бауырдағы өт қышқылынан өт қышқылдарының ішекке түсіп сіңіріліп, бауырға қан арқылы қайтарылу процесі - энтерогепатикалық айналым деп аталады.
Майлардың эмульгирленуі - бұл қорытылудың міндетті жағдайы, себебі, гидрофобтық субстратты суда ерітілген тамшыға айналдырып, гидролиттік ферменттер липаза әсеріне тартымды етеді. Эмульгирлену өт қышқылдарының қатысуымен, олардың амфофилді болғандықтан, май тамшыларын қоршап күрделі мицеллаларды түзілдіруімен өтеді.
Майлардың гидролизі панкреаттық липаза әсерімен атқарылады. Ол май тамшыларының бетіне сорбтеліп, (ТАГ) эфирлік байланыстарды ыдыратады. Бұл фермент аса активті емес. Оның активтілігі ұйқы безінде өндірілетін және сөлдің құрамында ішекке түсетін колипаза әсерінен жоғарылайды. Колипаза активсіз проколипазадан ішекте трипсин арқылы түзіледі. Колипаза классикалық активатор емес, ол тек субстратты байланыстырып және оны липазаның активті орталығына жақындатады.
Алғашқыда α-жағдайдағы май қышқылдары бөлінеді. Нәтижесінде, диацилглицерин, сонан кейін фермент әсерінен β-моноглицерин, гидролиздің негізгі өнімі түзіледі. Липидтердің эстеразасы әсерінен моноглицеридтердің бір бөлігі глицерин мен май қышқылдарына бөлініп гидролизденеді. Сондықтан майлардың қорытылуының өнімі бұл - глицерин, май қышқылдары және моноацилглицериндер.
Сіңірілу процесі де өт қышқылдарының қатысуымен өтеді. Олар холестерин, май қышқылдары, тұратын ерігіш комплексті түзеді. Ол гидролиз өнімдерін ішектің шырышты торшаларына өтуіне жағдай жасайды.
Өт қышқылдары қан ағысы арқылы бауырға жеткізіледі, сонан соң өт құрамында ішекке секреттеледі немесе бауыр → ішек → бауыр айналысына түседі. Бірақ тәулігіне 0, 3 г-ға жуық өт қышқылдары сіңірілмейді, нәжіс құрамында шығарылады. Бұл жоғалтылған өт қышқылдары бауырда холестериннен синтезделу есебінен толық толықтырылады.
ТАГ-лердің ресинтезі ыдыраған өнімдерден ішектің шырышты торшаларында өтеді. Ресинтез процесінде құрамы ағзаның майларына ұқсас ТАГ-дер түзіледі.
МАГ ДАГ ТАГ
α СH 2 -OH α СH 2 -OH α СH 2 -O-CO-R 1
β CH-O-CO-R 2 β CH-O-CO-R 2 β CH-O-CO-R 2
α CH 2 -OH α CH 2 -O-CO-R 3 α CH 2 -O-CO-R 3
R 1 -CO-S-KoA HS-KoA R 3 -CO-S-KoA HS-KoA
Ресинтезделген майлар лимфа жүйесі арқылы тек липопротеиндер құрамына қосылғаннан кейін ғана тасымалданады. Ішектерде ЛП-лардың екі түрі түзіледі: ХМ-дар және аз мөлшерде ТӨТ ЛП ХМ құрамында экзогендік майлар мен ұлпаларға жеткізіледі. Энтероциттерде түзілген ЛП-лар жетілмеген ХМ болып табылады және экзоцитоз жолдарымен алғашқыда лимфаға түседі. Ол арқылы үлкен қан айналу шеңберіне бауырды айналып өтіп түседі. Қанда жетілмеген Х-дар апобелоктар С ІІ мен Е-ні құрамына қосып жетілген ХМ-дарға айналады. ХМ қысқа өмір сүретін бөлшектерге жатқызылады, жартылай өмірінің кезеңі 0, 5-2 сағат.
Липопротеинлипаза (ЛП-липаза) - экзогендік майларды май ұлпаларының қабылдауын қамтамасыз ететін фермент. ЛП-липаза тамырлардың эндотелиінде орналасқан, қан ағынындағы ХМ-дармен әсерлеседі және жасушаға түскен ТАГ-дерді глицерин мен май қышқылдарына гидролиздейді. ТАГ-дердің алынуының мөлшері бойынша, ХМ-дар қалдық ХМ-дарға айналады және бауырға түседі (сур. 5) . Май ұлпасында ТАГ-дің гидролизі өнімдерінен қайтадан майлар ресинтезі өтеді (екінші ретті) . Ол сол жерде қажет болғанға дейін деполанады.
Майларға қажеттілік 50-100 г/тәулік және оның мөлшері қоректену мен энергия жұмсалуына тәуелді болады.
Сур. 4. Липопротеиндердің тасымалдаушы қызметі.
Майлардың қорытылуы мен сіңірілуі негізгі құбылыстары келесі схемадан байқалады:
Майлар қорытылуы Тағам майлары
(эмульгирленуі, гидролизі) өт қышқылы
тұздары
Эмульгирленген май
панкреаттық липаза
колипаза
Гидролиз өнімі
ДАГ өт қышқылы
МАГ (80%)
Мицеллалар түзілуі және
ішектің қабығында
сіңірілуі
Қоспалы мицелла
Майлардың ресинтезі 2 ацил-КоА
H 2 C-OH H 2 C-O-CO-R 1
HC-O-CO-R 2 HC-O-CO-R 2
H 2 C-OH H 2 C-O-CO-R 3
HS-KoA
Майлардың ХМ
құрамына жиналуы апо В-48
ХМ жетілмеген
Жетілген ХМ
түзілуі апо С-ІІ апо С-ІІ
апо В-48
апо Е апо Е
Майлардың деполануы мен мобилизациясы
Ағзадағы майлардың көзі:
- Тағамдық май (экзогендік) .
- Эндогендік майлар (бауырда көмірсулардан синтезделген) .
Майлар гликоген тәрізді энергетикалық материалдың деполануының формасы болып табылады. Майлар - біршама ұзақ мерзімді және тиімді энергия көзі. Егер майлардың түсуі ағзаның энергиялық қажеттілігінен жоғары болса, онда олар адепоциттерде деполанады. Сонымен бірге, егер көмірсулар көп мөлшерде (гликогенге деполанудан артық) болса, онда артық глюкоза мөлшері де майларға айналады.
Майлар май ұлпаларында келесі үш процестердің нәтижесінде жиналады:
- Хиломикрондардан (олар ішектен экзогендік майларды әкеледі) ;
- ТӨТ ЛП-дан (эндогендік майларды әкеледі) ;
- Май ұлпаларының торшаларында глюкозадан түзілуі;
Бірінші және екінші жағдайларда майлар ЛП құрамында ЛП-липаза арқылы гидролизденіп, торшаға май қышқылдары түседі, олар ТАГ синтезіне жұмсалады. Эндогендік майлардың синтезі көмірсулардан бауыр мен май ұлпасына түседі.
Липогенез
Бауырда да, май ұлпасында да майлардың синтезі фосфатид қышқылының түзілуі арқылы өтеді. Бірақ бауырда глицерофосфат екі жолмен жүреді:
- Глицериннің глицеролкиназамен активтену жолымен:
Бұл реакция қаннан бауырға түскен глицеринге тән.
- Фосфодиоксиацетонның тотықсыздануы жолымен глюкозаның ыдырауында түзілген.
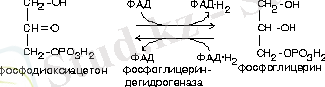
Глицериннен басқа бейтарап майлардың синтезі үшін май қышқылдарының активті формалары қажет. Кез-келген май қышқылының активті формасы - бұл ацил-КоА.
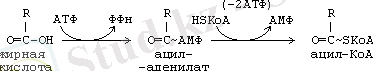
Бұл жерде АТФ-тің АМФ-ке дейін терең ылырауы байқалады (тотығып фосфорлануға түседі) . Сондықтан АТФ+АМФ 2 АДФ реакция болады. Сондықтан май қышқылдарынан активтенуіне 2 АТФ жұмсалуына эквивалентті болады. Майлардың синтезі жолындағы келесі этап фосфатид қышқылының түзілуінен басталады.
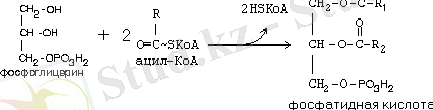
Реакция липогенездің негізгі ферменті глицерол-3- катализденеді. Бұл - ферменттің аллостерлік эффекторы, бірақ адипсин (ацилстимулдеуші белок) ацил-КоА-ның ферментпен әсерлесуін жеңілдетеді. Адипсин компонент жүйесінің бір компонентінің протеомерінің өнімі болып табылады. Жергілікті әсер ететін гормонға жатқызылады, себебі, май ұлпасында өндіріледі және сол жерде әсер етеді.
Триацилглицериннің ситезіне келесі екі реакция аяқтаушы болып табылады:
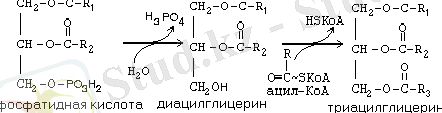
Май ұлпасындағы глицерил-3-фосфаттың көзі тек диоксиацетонфосфат болады, сондықтан адипоциттерде міндетті түрде гликолиз өтуі керек, ол глицеринге айналатын диоксиацетонфосфатты береді (сур. 6) .
Глюкоза
ДАФ Глицерин } тек бауыр жасушасында
АТФ өтеді
НАДН+Н + АДФ
глицерин-3-фосфат
НАД ацил-КоА
ацил-КоА
фосфатид қышқылы
Н 3 РО 4
ДАГ
Ацил-КоА
ТАГ
Сур. 6. Бауыр мен май ұлпасындағы майлар синтезі.
Синтездің реакциясы реакцияға қатысушы заттардың шығу көзіне тәуелді емес.
Бауырда синтезделген майлар ТӨТ ЛП-ға жинақталып, қанға секреттеледі. ТӨТ ЛП құрамында аполипопротеин В-100, С-ІІ, Е болады. ТӨТ ЛП-мен тасымалданатын майлар липопротеинлипаза әсерімен әртүрлі ұлпаларға гидролизге түседі, ерекше активті түрде май ұлпасындағы қан капиллярларында. Май қышқылдары жасушаларға өтеді және әртүрлі ұлпаларда әртүрлі жұмсалады; адипоциттерде майлар синтезі үшін миокардта, қаңқа бұлшықеттерінде АТФ түзіп тотығады (бұл ұлпалардың қызметі үшін қажет) .
Липопротеинлипазаның активтілігі инсулиннің әсерімен абсорбтық кезеңде жоғарылайды, адипоциттерде майлардың синтезі өтеді, онда қаннан түскен және глюкоза өнімдерінен түскен май қышқылдары пайдаланылады.
Тағам қабылдаған соң қандағы глюкоза концентрациясы жоғарылағанда инсулиннің секреттелуі жоғарылайды. Инсулин активтендіреді:
- Глюкозаның адипоцит ішіне тасымалдануын (ГЛЮТ-4) ;
- Липопротеинлипазаны, оның адипоциттердегі синтезін және капилляр бетінде бекітілуін. Май қышқылдары адипоцитке өтеді, ал глицерин бауырға тасымалданады. Адипоциттерде глицеринкиназа ферменті болмағандықтан, бос глицерин бұл ұлпада ТАГ синтезі үшін пайдаланылмайды;
- Активтелген май қышқылдары глицерол-3-фосфатпен әсерлеседі де, фосфатид қышқылы арқылы ТАГ-ге айналады, олар адипоциттерде деполанады (Сур. 7) .
Қан капиллярларының
қабырғасы
глюкоза
+
инсулин
липопротеинлипаза
ХМ
ХМ қал
май
қышқылдары АДИПОЦИТ
глицерин
ТӨТ ЛП бауыр
ТА ЛП
ТТ ЛП
ҚАН
... жалғасыСіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz