Азот тотығының бидай алейрон клеткаларындағы супероксиддисмутаза белсенділігіне және супероксид радикалының жинақталуына әсері

Әл - Фараби атындағы Қазақ Ұлттық Университеті
Биология факультеті
Генетика және молекулалық биология кафедрасы
БIТIРУ ЖҰМЫСЫ
Бидай алейрон клеткаларындағы азот тотығының супероксиддисмутаза белсенділігі мен супероксид радикалының жинақталуына әсері.
Орындаушы
4- курс студенті Паренова Р. А.
Ғылыми жетекшісі
б. ғ. к., доцент Бисенбаев А. Қ.
“”2007ж.
Норма бақылаушы Сатылған И. А.
“”2007ж.
Қорғауға жіберілді.
Кафедра меңгерушісі,
биология ғылымдарының докторы Айташева З. Г .
“”2007ж.
Алматы, 2007
МАЗМҰНЫ
РЕФЕРАТ . . . 3
ҚЫСҚАРТЫЛҒАН СӨЗДЕР ТІЗІМІ . . . 4
КІРІСПЕ . . . 5
- ӘДЕБИЕТКЕ ШОЛУ . . . 6Астық тұқымдастарының алейрон қабаттары . . . 6Оттегінің белсенді түрлерінің химиясы . . . 9Азот тотығы мен гиббереллин қышқылының метаболизмдері . . . 12Супероксиддисмутаза ферментінің құрылысы мен функциясы . . . 16
2. ЭКСПЕРИМЕНТТІК БӨЛІМ . . . 20
2. 1 Материалдар мен зерттеу әдістері . . . ……. . . . … . . . 20
2. 1. 1. Бидай дәнінің жекелеген алейрон ұлпасын бөліп алу… . . . 20
2. 1. 2. Супероксиддисмутаза изоферменттерін экстракциялау және активтілігін анықтау . . . 20
2. 1. 3 Супероксиддисмутаза изоферменттерін электрофоретикалық сұрыптау және электрофорезден соң айқындау . . . 21
- Супероксид радикалы мөлшерін анықтау …… . . . . . . 22
3. НӘТИЖЕЛЕР ЖӘНЕ ОЛАРДЫ ТАЛҚЫЛАУ . . . 23
4. ҚОРЫТЫНДЫ . . . 28
5. ҚОЛДАНЫЛҒАН ӘДЕБИЕТТЕР ТІЗІМІ . . . 30
РЕФЕРАТ
Бітіру жұмысы 33 беттен тұратын компьютерлік текстен, 5 суреттен тұрады.
Жұмыстың мақсаты: Бидай алейрон клеткаларындағы азот тотығының супероксиддисмутаза белсенділігі мен супероксид радикалының жинақталуына әсерін зерттеу.
Жұмысты жасау үшiн объект ретiнде: Қазақстан 4 сортты гексаплоидты бидайдың (Triticum aestivum) дәндері қолданылды.
Жұмыс барысында келесi әдiстемелер қолданылды: Бiркелкi буферлiк жүйедегi ПААГ электрофорезi, супероксиддисмутаза ферментiн экстракциялау және белсендiлiгiн анықтау, белок мөлшерiн анықтаудың Бредфорд әдiстемесi.
Кілтті сөздер: Оттегінің белсенді түрлері, тотықтырушы агенттер, тотықтырушы қопарылыс, антиоксидантты ферменттер, супероксиддисмутаза.
ҚЫСҚАРТЫЛҒАН СӨЗДЕР ТІЗІМІ
ОБТ- оттегінің белсенді түрлері
СОД- супероксиддисмутаза
АБҚ - абсциз қышқылы
ГҚ - гиббереллин қышқылы
NO - азот тотығы
SNAP - S-нитрозо-N-ацетилпенициламин
КПӨ - клетканың программаланған өлімі
ПААГ - полиакриламидтік гель
О * - супероксид радикалы
Н 2 О 2 - сутегінің асқын тотығы
КІРІСПЕ
Супероксид, сутектің асқын тотығы және гидроксид радикалы секілді оттегінің белсенді формаларының клеткалардың программаланған өлімінiнiң iске асу механизiмiндегi ролі зор. Мысалы, гидроксил қалдығы лезде 7х10 -10 с ішінде клетка құрлымын бұзып, ондағы белок, нуклеин қышқылдарының құрылымын бұзып, липидтерді аса тотықтырып, тотығу өнімдері: альдегид, кетон, асқын тотықтармен қатар, аса улы қосылыстардың түзілуін жүзеге асырады. Осылардың ішінде нуклеин қышқылдарының зақымдалуы ДНҚ-дағы генетикалық информацияны өзгертіп, организмнің тіршілігіне қауіп тудыруы мүмкін.
Антиоксидантты ферменттердің ішінде ОБТ қарсы клетка ішілік қорғаныстың алғашқы қатарын супероксиддисмутаза (СОД) құрайтыны анықталған. СОД өсімдік клеткасында детоксикация реакциясын катализдейді. Ол супероксид радикалдарын өзара әсерлестіріп, аса улы супероксид радикалының улылығы төмен сутегінің асқын тотығына айналуын катализдейді.
Осыған орай жұмысымның мақсаты алейрон қабатының клеткаларындағы супероксид радикалының жинақталуы мен супероксиддисмутаза ферментінің активтілігінің азот тотығы арқылы реттелуiн зерттеу болып табылады.
- ӘДЕБИЕТКЕ ШОЛУАстық тұқымдастарының алейрон қабаттары
Астық тұқымдастарының алейронын зерттеу кем дегенде екі ғасырға созылып келеді. Астық алейроны өсімдіктердің ең қарқынды түрде зерттеліп келе жатқан ұлпаларының бірі болып табылады. Сыра қайнатуда және ашытқы дайындауда астық алейроны маңызды орын алатындықтан химия және биология мамандары шамамен 200 жыл бұрын арпаның маңызын егжей-тегжейлі зерттей бастады [1] .
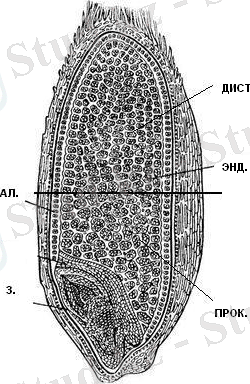
ЕСКЕРТУ: АЛ. - бидай дәнінің алейрон қабаты; ЭНД. - бидай дәнінің крахмалды эноспермі; ПРОК. - бидай дәнінің проксимальды бөлігі; ДИСТ. - бидай дәнінің дистальды бөлігі. 3 - бидай дәнінің ұрығы
Сурет 1- Бидай дәнінің құрылысы
Бидай дәнінің эндоспермі тозаң қапшығында, орталық клетканың аталық жыныс клеткасымен ұрықтану нәтижесінде пайда болатын, триплоидты ұлпа (1-сурет) . Тозаңдану процесінен кейін шамамен 8-10 тәуліктен соң, дән эндоспермі крахмалды эндоспермге және алейрон қабатына дифференциацияланады [2] . Алайда, ұрықтанған бір клеткадан пайда болатынына қарамастан, алейрон ұлпасы мен крахмалды эндосперм клеткаларының өлуі әртүрлі уақытта жүзеге асады. Крахмалды эндосперм клеткалары бидай дәнінің пісіп-жетілу барысында өліп, дән қуысын белокты және крахмалды гранулалар түрінде толтырып тұрады деген болжам бар. Ал алейрон клеткалары, дәннің пісіп-жетілу сатысында еш өзгеріссіз сақталып, тек қана оның өніп-өсу сатысын, гидролитикалық ферменттерді синтездеп, секрециялау арқылы индукциялағаннан кейін ғана элиминацияланады [3] .
Алейрон қабатының онтогенетикалық программаланған клеткалар өлімі (ПКӨ) апоптотикалық сипатта жүзеге асатыны анықталған [4-6] . Бұл процестің белсенділігінің артуымен, геномдық ДНҚ молекуласының фрагментациялануымен үйлесіп, фитогормондар - гиббереллин және абсциз қышқылдарымен бақыланылатыны көрсетілген [7] .
Жоғарыда айтылғандай, алейрон клеткаларының негізгі қызметі гидролитикалық ферменттерді (α-амилазаны қоса) синтездеп, крахмалды эндоспермге секрециялау.
Алейрон клеткаларының ультра құрылымы олардың қызметін көрсетеді. Дәндегі алейронның көптеген көрсеткіштері оның ерекше және жан-жақты тәжірибелік жүйе болуын қамтамасыз етеді. Астық тұқымдастарының алейрон қабаты біртекті, жоғары дифференциалданған клеткалардың бір қабатынан (сұлы, жүгері, қара бидай, бидай) немесе бірнеше қабатынан (күріш, арпа) тұрады [8] . Пісіп-жетілген, құрғақ алейрон клеткалары қалың, геммицеллюлозалық қабырғамен қоршалған және олар ешқандай да клеткалық бөлінуге ұшырамайды. Алейрон қабаттарын жабысып қалған өлі крахмалды эндоспермнен ажыратып алуға болады және ажыратып алынған алейрон ұлпасынан ферменттердің көмегімен протопластардың біртекті популяциясын дайындауға да болады [9] . Осындай жолмен жекеленіп алынған протопластар, тұтас алейрон ұлпалары сияқты гиббереллинге тәуелді α-амилаза изоферментінің активтенуі және синтезін қамтамасыз ететіні көрсетілген. Жекеленген протопластар алу әдістемесі сұлы және арпа алейрон ұлпасына қатысты жасалынған [10] Біздің лабораторияда бидай алейрон ұлпаларынан жекеленген протопластарды бөліп алудың және тазалау, оларды концентрлеу үшін фракциялаудың модификацияланған әдістемесі ұсынылған [11] . Ферменттік өңдеу арқылы алынған бидай алейрон протопластарының шығымы шикі ұлпаға шаққанда -35х10 6 клетка/1 г болды. Протопластарды фиколл градиенттінде центрифугалау әдістемесі тіршілік дәрежесі жоғары клеткалар фракциясын алуға мүмкіндік берді (шамамен 80-85 %) .
Алейрон клеткаларының цитоплазмасына белоктар қорынан тұратын көптеген вакуольдер тән, оларды көбінесе алейрондық түйіршіктер деп те атайды, олар бүкіл клеткаларды алып жатады. Бұл органелла алейронның гормондар әсеріне қайтаратын реакциясында негізгі қызмет атқарады. Протеиндердің қоры бар вакуольде гиббереллинмен өңделгеннен кейін гидролизге ұшыраған протеиндерді секреторлық белоктардың түзілуіне қажетті аминқышқылдармен қамтамасыз ету үшін қор ретінде сақтай бастайды [12] . Протеиндердің қоры бар вакуольдер, сондай-ақ, астық тұқымдастардың дәндеріндегі минералдардың негізгі қоймасы болып табылады. Олардың құрамынан минералдар фитин түрінде бөлініп алынған. Фитин К + , Mg 2+ , және Ca 2+ иондары мен фитинді қышқылынан (гексофосфат) түзілген кристалды, ерімейтін комплекс болып табылады. Бұл кристалды қосынды, глобоид деп те аталады. Рентгендік микроанализ нәтижелері бойынша дән құрамындағы РО 4 3+ , К + , Mg 2+ және Ca 2+ иондарының шамамен 75% алейрон қабаттарында сақталады [13] . Алейрон немесе протопластардың инкубациялық кезеңінде вакуольдер көлемдері ұлғайып, бір-бірімен орталық үлкен бір вакуоль түзілгенше қосыла бастайды. Инкубациялық ортада гиббереллин болған жағдайда клеткалар мен протопластардағы алейрон қабаттарының вакуольдену процесі жылдамдай түседі. Бейтарап майларды сақтайтын липидтік қосындылар немесе олеосомдар да алейрон клеткаларының маңызды органеллалары болып табылады, олар клетканың бүкіл кеңістігінің 30% алып жатқан үшглицеридтік негізден тұрады [14] . Алейрон клеткаларындағы олеосомдар эндоплазмалық торда пайда болып, кейіннен эндоплазмалық торға және протеиндердің қоры бар вакуольдер бетіне бекінген күйде болады. Олеосомдардың вакуольдер бетіне бекініп жатуы вакуолдердің кейбір түрлері тікелей эндоплазмалық тордан пайда болады деген болжамды растайды. Қор ретінде сақталған бейтарап майлар алейрон клеткаларын көміртегі көзімен де қамтамасыз етеді, эндоспермді мобилизациялау басталғанға дейін клетка көміртегіні осы жерден жұмсай алады, сонымен бірге мұндағы май қышқылдары мембрананы түзуге пайдаланылады. Бұл май қышқылдарының алейрон клеткалары арқылы метаболизденетініне бірнеше дәлелдер бар. Алейрон клеткаларында малатсинтетазадан және изоцитрат лиазадан тұратын көптеген глюкоксисомалар да болады [15; 16] және оқшауланып алынған алейрон қабаттары сахарозаны синтездей алады.
Жоғарыда айтылғандай, алейрон клеткалары - жоғары деңгейде дифференциацияланған ұлпа. Бұл ұлпаның негізгі қызметі - гидролитикалық ферменттерді (альфа-амилазаны қоса) синтездеп, крахмалды эндоспермге секрециялау. Олай болса, алейрон ұлпаларында оның секреторлық қызметіне байланысты эндоплазматикалық тор, Гольджи аппараты, митохондриялар өте жақсы дамыған.
Аталған гидролитикалық ферменттердің ішінде α-амилаза, дәннің өніп-өсуіне қатысты маңызды болып табылады. Крахмалды эндоспермге секрецияланған α-амилаза крахмалды қанттарға дейін ыдыратып, өсімдіктің өніп-өсуіне қажет энергиямен қамтамасыз етеді [17; 18] .
α-амилаза ферментінің практикалық маңыздылығына байланысты (ауыл-шаруашылығы, сыра дайындау, нан өндіру, фармакология және т. б. ) оларды зерттеу жұмыстарына ғалымдар ертеден көңіл аударуда. Осыған орай альфа-амилаза ферментінің құрылысы мен қызметі келесі бөлімде терең қарастырылады.
1. 2 Оттегінің белсенді түрлерінің химиясы
Өсімдіктер өздерінің тіршілік кезеңі барысында қоршаған ортаның әртүрлі қолайсыз ықпалдарына қарсы тұруы тиіс. Осыған байланысты олардың бойында өсімдік организмін биотикалық және абиотикалық қолайсыз жағдайлардан қорғауға бағытталған кең ауқымды бағдарламалар пайда болып, дамыды. Өсімдіктердің потогенді (ауру тудырғыш) бактериялармен, саңырауқұлақтармен және вирустармен зақымдалуы олар үшін ең қауіпті жағдайлардың бірі болып табылады [19] .
Микроорганизмдерді өсімдіктерге ендіргеннен кейін олар, патогендердің көбеюін шектеу мен жою мақсатында бір немесе бірнеше қорғаныс механизмдерін іске қосуы мүмкін [20] . Клетка қабырғаларының қалыңдауы, фитоалексиндердің түзілуі және антимикробиотикалық белоктардың жиналуы сияқты жалпы қорғаныстық реакцияларды жүзеге асырады. Бұл реакциялардың уақыт пен кеңістік аралығында реттелуі ие организммен патоген арасындағы қарым-қатынастың нәтижесін қамтамасыз етуде шешуші фактор болып табылады (организм не микроорганизмнің ықпалына душар болып, беріледі, не оған қарсы әрекет етеді) [21] . Көп жағдайда, бұл реакциялар зақымдалған ошақта бірнеше клеткалардың өлімге ұшырауымен байланысты болады (гиперсезімталды жауап) . Қорғаныстық реакциялардың жүре бастауы үшін не организмге енген патоген арқылы түзілген, не өсімдіктің клеткалық қабырғаларынан босап шыққан белгі бергіш молекулалардың қабылдануы қажет. Мұндай молекулалар "элиситерлер" (ағылш . elicit-қоздырып-шақыру, шығарып алу) деп аталады, олардың кейбі-реулері олигосахаридтер, белоктар және пептидогликандар ретінде иденти-фикацияланады [22] . Бұл элиситерлер ие организм -микроб түріндегі белгілі бір өсімдік жүйесі үшін арнайы түрдегі молекулалар болуы мүмкін, немесе арнайы емес сипаттағы молекулалар түріндегі, мысалы, өсімдіктердің клеткалық қабырғаларының бөліктері немесе өсімдік организміне енген патогеннің клеткалық бөлшектері ретінде болуы мүмкін.
Өсімдіктердің суспензия-культурасы ортасында қорғаныстық реакция-ларды тудыру үшін элиситер молекулаларының қоспасы қолданылған жағдайда, қорғаныс жауабының негізінен клетка қабырғасының бетінде жүзеге асатын, жылдам, тез жүруші процесстерден тұратын жаңа жақтары анықталады. Реакциялардың ішінде мынадай түрлері идентификцияланады : Оттегінің белсенді түрлерінің (ОБТ) босатылуы (тотықтырушы қопарылыс, ТҚ) [23] ; клеткадан тыс рН мәнінің, мембрандық потенциалдардың, иондар қозғалысының өзгеріске ұшырауы; белоктарды фосфарлау моделінің өзгеруі [24] ; клеткалық қабырғаның белоктық бөліктерінің тотыға иммобилизациялануы [25] .
Клеткалардың көпшілігі ОБТ-рін өндіруге және бейтараптауға қаблетті келеді. Қалыпты жағдайда ОБТ клеткада молекулалық оттегінің (О 2 ) бір электрондық тотықсыздануы нәтижесінде міндетті түрде түзіліп отыратын қосымша өнім ретінде кездеседі. Сонымен қатар, клеткадағы ОБТ деңгейінің мүмкіндігінше ең төмен болуын қамтамассыз ету үшін көптеген клеткаларда арнайы қорғаныстық механизмдер болады.
Сүтқоректілерде ОБТ өндіру реакциясы жүретіні бізге 30 жылдан астам уақыттан бері белгілі (фагоциттердегі респираторлық жарылу) өсімдіктерде бұл құбылыс едәуір кеш анықталады [26] . Жуық арада жаряланған жұмыстардың нәтижесі бойынша ОБТ тек организмдерге енген патогенге қарсы қорғаныс құралы ретінде ғана қызмет атқарып қоймай, өсімдіктердің бұдан кейінгі қорғаныстық реакцияларын, сонымен бірге, зақымдалған клеткалардың гиперсезімталды жауабын белсендіруші сигнал болып табылады.
ОБТ молекулалық оттегінің белгілі бір ретпен тотықсыздануы барысында түзелетін : супероксид (О), сутегінің асқын тотығы (Н 2 О 2 ), және гидроксил радикалы (ОН) өнімдері. Бұл қосылыстар стреске ұшырамаған клеткалардың митохондриялары мен хлоропластарындағы жүретін тотығу-тотықсыздану реакциялары нәтижесінде аз мөлшерде түзіледі.
Супероксид (О) түзілу үшін энергия аз мөлшерде жұмсалады. Тірі клеткалардағы супероксидтің мөлшері оның протондалған түрі гидропероксил радикалдың (О 2 Н) мөлшерімен бірдей болады. Гидропероксил радикалы супероксидке қарағанда анағұрлым гидрофобты келеді және мембраналардың липидті қабаттары арқылы оңай енеді. рН-мәні физиологиялық болған жағдайда супероксид клетканың макромолекулаларымен әлсіз түрде әрекеттеседі. рН мәні бейтарап немесе әлсіз сілтілі болған жағдайда супероксидте, сутегінің асқын тотығы да Н 2 О 2 және О 2 ыдырайды. Бұл реакция кенеттен жүруі мүмкін, сондай-ақ (СОД) қатысуыменде жүреді.
Н 2 О 2 салыстырмалы түрде алғанда тұрақты ОБТ болып табылады. Н 2 О 2 реакцияға түсу қаблеті жоғары емес. Ол электрлік тұрғыдан алғанда бейтарап бола отырып, клеткалық мембраналар арқылы еніп, Н 2 О 2 түзілген аймақтан алшақ жатқан жерлерге жетуге қабілетті келеді. Өсімдік клеткаларында жаңадан түзілген Н 2 О 2 мынадай өзгерістерге ұшырауы мүмкі: 1) өздігінше кенеттен немесе каталазаның қатысуымен су және оттегіге ыдырайды; 2) әртүрлі пероксидазалар үшін субстрат ретінде болуы мүмкін, мысалы, лигниннің түзілуін құрлымдық тұрғыдан тежейтін феноксил радикалдарының түзілуі үшін субстрат болып келеді; 3) және глутатион-редуктазамен бірлесіп әсер ететін аскорбат пероксидаза көмегімен зияндылығы жойылуы мүмкін (халливела -Асада жолы) . Белгілі бір жағдайда, тотықсыздандырғыш болған кезде, Н 2 О 2 пероксидазалар арқылы Compound III түзілуін қамтамассыз ететін реакциялардың ретті тізбегіне айналуы мүмкін.
Гидроксил радикалы бұл жұмыста аталған ОБТ ішінде реакцияға түсу қаблеті ең жоғары болып келетін түрін құрайды. Ол Н 2 О 2 мен О өзара әрекеттесуі нәтижесінде пайда болуы мүмкін. Қалыпты клеткаларда бұл реакция баяу жүреді және ОН көп мөлшерде өндірілуі үшін тиімсіз болып табылады. Бірақ, Fe 2+ немесе Cu + сияқты транзиттік металлдардың тотығуынан және бұл тотыққан иондардың супероксидтің қатысуымен тотықсызданған түрлеріне дейін регенерациялануынан тұратын реакциялар циклы барысында ОН айтарлықтай көп мөлшерде түзіледі. ОН радикалдық тізбекті реакцияларды бастауға қаблетті болғандықтан, ол клеткалық макромолекулалардың қайтымсыз өзгерістерге (модификацияға) ұшырауына және органеллалардың зақымдалуына жауапты болып келетін негізгі ОБТ ретінде саналады. ОН бос күйінде болуы шамамен 0, 2 сек тең болғандықтан оны көзбен көру және оның патоген-өсімдік қарым-қатынасындағы рөлін анықтау сәтсіз аяқталды. N-ацетилхит-олигосахаридті элиситермен өңделген күріштің (Oryza sativa) суспензиялық культура клеткаларына жақында ғана жүргізілген тәжірибелер нәтижесінде, бұл модельдік жүйеде ОН түзілетіні анықталды [27] .
Өсімдік клеткаларының көптеген жануар жүйелерінен ерекшелігі, олар ОБТ негізінен алғанда Н 2 О 2 түрі үнемі, айтарлықтай көп мөлшерде өндіруге қаблетті келеді, бұл процесс әдетте клетканың экстрацеллюларлық компар-таментімен байланысты болады және ол гормондар жарық сәулесі, механикалық зақымданулар сияқты әртүрлі фактолар арқылы реттеледі. Н 2 О 2 лигнификация процесіне ұшырайтын клеткаларда кездеседі, мысалы, трахеялық (түтікше) элементтерден және флоэма талшықтарынан көруге болады, ал тез ұзарушы клеткалардың клеткалық қабырғаларында әдетте болмайды; механикалық стреске шалдығып зақымдалған клеткаларда Н 2 О 2 қарқынды түрде өндіріледі.
Тотықтырушы қопарылыс дегеніміз, сырттан түрткі болған факторларға жауап ретінде ОБТ көп мөлшерде, жылдам өндірілуі болып табылады. ОБТ өндірілу жайында алғашқы мәліметтер 1983 жылы пайда болды, бұл мәліметтер бойынша картоп түйіндеріне (Solanum tuberosum) өзімен сәйкес келмейтін Phytophthora infestans тұқымын өндіріп өсірген (инокуляциялаған) жағдайда картоп түйіндері супероксид түзе бастайды. Бұл реакция сондай-ақ потогенді микроорганизмдермен зақымдалған өсімдіктерде және элиситерлік препараттармен, қоздырғыштың бөліктерімен өңделіп немесе механикалық стреске ұшыратылып өсірілген клеткаларда да байқалады. Көптеген мәліметтер бойынша, тотықтырушы қопарылысқа қатысатын негізгі ОБТ - Н 2 О 2 болып табылады, О қатысуы да мүмкін. Н 2 О 2 мен О түзілуі химиялық тұрғыдан алғанда ұқсас келеді, сондықтан тотықтырушы қопарылыстағы СОД (NN-диэтилдитиокарбамат) ингибиторының әсерінен бұзылатындықтан, Н 2 О 2 СОД қатысуымен О-ден түзіледі деген болжам бар.
ОБТ өнімдерінің кенеттен жоғарлауы әдетте жылдам түрде өтеді, сондай-ақ, әр-түрлі зиянды факторларды пайдаланған жағдайда зерттеліп жатқан түрлі жүйелерде өзгеше болады. Суспензиялық - культура клеткаларында реакция әдетте, ортаға элиситерлерді қосқаннан кейін 1-2 минут өткен соң басталып, бірнеше минуттан кейін ең жоғарғы шегіне жетеді де элиситер-леудің бсталғанына 30-60 минут болғанда аяқталады. Тотық-тырушы қопарылысты зерттеу үшін өсімдіктің бөліктерін пайдаланған кезде реакция айтарлықтай кеш байқалды: элиситерлеуден кейін 8-12 сағатта басталып, 2-4 сағаттан соң ОБТ түзілуінің айқын белгілері көрінді. Мұны салат-латук өсімдігіне әйкес келмейтін Pseudomonas syringеae тұқымын инокуляциялағаннан кейін 5 сағат өткен соң салат-латук өсімдігі in-vivo жағдайында Н 2 О 2 өндіретінінің анықталғандығы растай түседі. Басқа мәліметтердің хабарлауынша, қиярдың (Cucumis sativus) кутикуласын алып тастап, гипокатиль аймағын не салицил қышқылымен (СҚ) не 2, 6-дихлороизоникотин қышқылымен (ИНҚ) өңдеу арқылы қиярдың гипоко-тилінде жылдам жүретін реакцияны тудыруға болады, салицил қышқылы мен 2, 6- дихлороизоникотин қышқылы тіршілік ету барысында пайда болатын жүйелі төзімділіктің индукторлары болып табылады. Мұндай гипокотильдерде Н 2 О 2 өндірілуі элиситерлеуден кейін 20 минут өткеннен соң аяқталады, бұл суспензиялық культура клеткаларында байқалатын өзгеріске ұқсас келеді.
1. 3 Азот тотығы мен гиббереллин қышқылының метаболизмдері
NО- газ тәрізді бос радикал, сондықтан да ол клеткааралық зат пен плазматикалық мембрана арқылы оңай өтеді [28] . NО клетка ішілік мессенджер жүйесінің бір бөлігі бола тұра, жануарлар жүйесінің физиологиялық процесстеріне кең көлемде әсер етеді [29] .
Қазіргі уақытта әдебиеттерде жинақталған мәліметтерге қарағанда, NО сонымен қатар өсімдіктерде өтетін көптеген физиологиялық процесстерге қатысатындығын көрсетеді: патогенге жауап реакциясы, өсіп-өну, фитогормондардың мөлшері, сонымен қатар КПӨ (апоптоз) . Сонымен қатар азот тотығы антиоксидант, яғни клеткаішілік радикалдардың деңгейін төмендететін агент ретінде де қарастырылады [30] .
NО тасымалдануы. NО диффузияланушы газ екендігіне қарамастан, оның ізашарлары немесе кешендері жақын немесе алыс қашықтыққа көшіп отыратын NО қоры «депосы» немесе транспорттық түрі ретінде қызмет атқаруы мүмкін. NО тасымалдануы, этиленнің ізашары - АСС тасымалдануына ұқсас аналогиялық жолмен жүреді.
Жануарлардың қан айналымында S-нитрозогемоглобиннің болуы, оның циркуляциялық азот тотығының көзі ретінде қызмет атқаруы мүмкін деген болжамды тудырды. Қазіргі уақытта бұл болжам күмән келтіреді, алайда бұл болжамның орнына азот тотығының көзі ретінде нитрит қызмет атқаратындығы туралы мәліметтер ұсынылды. Нитрит өсімдіктерде де NО көзі ретінде қызмет атқара алады және бір орыннан екінші орынға жылжи алады.
... жалғасыСіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz