Молекулалық биология: даму тарихы, зерттеу әдістері және клеткалық эволюция

Жоспары
- Молекулалық биология
- Молекулалық биологияның қысқаша даму тарихы
- Клетканы зерттеу әдістері
- Клетка эволюциясы
- ДНК молекуласының құрылысы
- Вирустар-молекулалық биологияның зерттеу объектісі
- Пайдаланылған әдебиеттер
Молекулалық биология
Молекулалық биология - тіршілік белгілері мен олардың негізгі қасиеттерін молекулалық деңгейде зертейді. Молекулалық биологияның негізгі зерттеу бағыттары - 1) клеткалардыц генетикалық аппаратының құрылымдық-функционалдық ұйыдасуы мен тұқым қуалау иформациясының жүзеге асу механизмдерін (молекулалық генетика), вирустардың клеткалармен өзара әсерлесуінің молекулалық механизмдерін (молекулалық вирусология), ағзаның иммундық реакцияларының заңдылықтарын зерттеу (молекулалық мунология), ағзаныц жеке дамуьшдагы әр түрлі сапалы клеткалар мен маманданған клеткалардың пайда болуып (дамудың молекулалық биологиясы) зерттейді.
Молекулалық биологияның практикалық маңызы - ауыл шаруашылығында (бағытталған және бақыланатын жануарлар мен өсімдіктердің тұқым қуалау аппараттарындағы өзгерістер, жоғарты өнімді сорттарды, ұйтұқымдарды шығару), микробиологиялық өнлірісте (биологиялық белсенеді комноненттер мен ақуыздардың бактериалды синтезі) ; молекулалық биология медицинаның көптеген бөлімдерінің теориялық негізі болын табылады (вирусология, иммунология және т. б. ) .
Молекулалық биологияныц алдында тұрған міндеттері - қатерлі ісіктердің молекулалық негіздерін анықтау мәселесі, тұқым қуалайтын аурулардың алдын алу, гормондардың, ұлы және дәрілік заттардың молекулалық өсерін анықтау, естің механизмдерін, жүйке процсетерінің табиғатын тану. Жануарлардың генетикалық аппаратын бағытталған түрде өзгертуге мүмкіндік беретін гендік инженерияның дамуының маңызы зор. Молекулалық биология биохимия, биофизика, биоорганикалық химиямен бірігіп әдетте-физико-химиялық биология бағытын құрайды.
Молекулалық биологияның негізгі зерттеу объектілері - вирустар, соның ішінде бактериофаттар, клеткалар және субклеткалық структуралар (ядролар, митохондриялар, рибосомалар, хромосомалар, клеткалық мембраналар), сонымен қатар макромолекулалар (ақуындар, нуклеин қышқылдары) болып табылады.
Молекулалық биологияның маңызды жетістіктеріне - кейбір ақуыздардың құрылымын анықтау және олардың құрылысы мен қызметінің байланысын тағайындау (М. Перуд, Дж. Кендрю, Ф. Сенгер, К. Анфинсеи және т. б. ), нуклеин қышқылдары мен рибосомолардың құрылымы мен биологиялық механизмін анықтау (Дж. Уотсон, Ф. Крик, Р. Холли т. б. ), генетикалық кодты ашу (М. Ниренберг, С. Очоа), кері транскрипцияның ашылуы (Х. Тсмин, Д. Балтимор), ақуыз молекуласы (Ф. Крик, Ф. Жакоб, Ж. Мопо) мен нуклеин қышқылының (А. Корнберг, С. Очоа) бирсинтезініц негізгі кезеңдерінің механизмдерін зерттеу, вирустардың құрылысын және олардың фепликациясының механизмін ашу, генетикалық инженерияның (П. Берг, В. Арбер, Г. Смит, Д. Натанс) және геннің синтезінің өдістерін ашу, орыс галымдары биополимерлердің матрицалық синтезініп принциптерін (И. Кольцов) анықтуы, жоғары сатыдағы өсімдіктерде ДНҚ-ның болатындыгы дәлелдеуі (Н. Белозерский) қатерлі ісіктердің пайда болуының вирусогенетикалық теориясы жасалды траспорттық РНҚ-ғы нуклеотидтердің кезектесуі анықталды (Л. Басв) .
Молекулалық биологияның қысқаша даму тарихы
Тұқым қуалау процестерін молекулалық деңгейде түсіну - зат алмасуға байланысты ауруларды зерттеу нәтижесінде алынған мәліметтерден басталды. 1908 ж. ағылшын дәрігері Гэррод алкаптонурияның тұқым қуалайтының байқады. Яғни зат алмасу процестсрінің (химиялық реакция) де басқа да қазір белгілі феиотитітік белгілер тәрізді тұқым қуалайтыны анықталды,
1930-1950 ж. ағзалардаға зат алмасудың негізгі жолдары анықталды. Микроорганизмдер, өсімдіктер мен жануарлардағы биокинетикалық реакциялардың (гликолиз, цитраттық цикл, амин қышқылдарының синтезі, нуклеотидтер синтезі және т. б. ) маңызды сатыларының ұқсас екендігі белгілі болды.
Бидл, Татум 1941, 1948 жылдары бір ген - бір фермент гинотезасын ұсынды. Сөйтін генетика мен биохимияның арасындағы байланыс табылды. XX ғасырдың бірінші жарынысында биохимиялық зерттеулердің негізгі объектілері - тек кіші молекулалар болды. Молекулалық биология мен казіргі биохимия макромолекулаларды зерітеумен айналысады. Олардың қызметін түсіну үшін "әлсіз" байланыстарды (сутрінік байланыс, иондық байланыс, ван-дер-ваальс күштері және т. б. ) білу керек. Бұл байланыстардың кемегімен клеткалар мен молекулаүстілік комплекстердің компонеінтерін біріктірін ұстан тұрады жәнс олардың өзіндік ерекшелігі макромолекулалардың матрицалық қызметі үшін жауапты.
Молекулалық генетика бірнеше даму сатысын егін, жинақталған көптесін ғылыми нәтижелер қазір классикалық болын табылды. 1950-60 жылдарда гендік материалдың құрылысы белгілі болды. Генстикалық ақпараттың ДНҚ-ның сызықтық нуклеотидтік кезектесуінде екендігі дәлелделді.
Уотсон мен Криктің моделі ДНҚ-ның ренликациясы қалай жүзеге асатының көрсетті. Информацияның ДНҚ-дан РНҚ-ға - одан әрі ақуызға (транскрипция, трансляция) қарай жүретіні анықталды, генетикалық код табылды.
Молекулалық генетикадың даму нәтижесінде молекулалық биология мен клеткалық биология пайда болды. Эмбрионалдық дамуды молекулалық деңгейде зерттеу басталды. Бөліну негізінде әр түрлі гендердің кезектесіп қосылатыны керсетілді. Осыған байланысты қазіргі уақытта да канағаттанарлық шешімі өлі табылмаған сұрақ туды: гендердің белсендігі қалай реттеледі? (прокариоттар вирустар мен бактерияларда транскрипцияның жылдамданы анықтайтын көптеген сигналдар алынады.
Күрделі процестер тек күрделі структураларда жүреді. Мембраналар, рибосомалар, митохондриялар, жиырылған элементтер және басқа да структуралар көптеген макромолекулалардан тұратын жүйелерден тұрады, бұл жүйелер бір-бірімен көптеген әлсіз байланыстардың көмегімен бірігіп тұрады.
Олигомерлік комплекстердегі макромолекулалар кооперативтік жүйе болып табылады.
Клетка ішінде жүретін процестерді түсіну үшін молекулалардың молекулаүстілік структурадагы өрінен, әрскетін зерттеу қажет. Молекулалардың кооперативтік өзара байланысы - фотосинтез, тыныс алу, ақуыз биосиитезі процестерінің жүзеге асуының негізгі алғы шарты. Қарапайым клетканың болуы мүмкін емес. Е. соlі-діц өзі күрделі молекулалардан құралған.
Е. соlі зерттелген соң in vitro адамның клеткалары өсіріле бастады.
Неғұрлым ағза күрделі болса, соғұрлым генстикалық информациясы кен болады. Ұсақ вирустарда бар болганы 3 ген болса, адамда 50 000. Эукариоттарда информацияны тасымалдауыны (ДНК) мен орындаушы механизмдер (ақуыз биосинтезі) кеңістікте бір-бірінеп бөлінген.
Кәпклеткалы организмдерле информацияны өндісудің күрделі механизмдеріне иммундық және нерв жүйесі жатады.
Биохимияға молекулалық идеялардың өнуі клеткалық биологияға бурый бір-бірімен байланыссыз пәндердін-морфология.
Молекулалық биологияның маңызын дарвиндік эволюциялық ткорияның маңызымен салыстыруға болады.
Криктің айтуынша молекулалық биологияның дамуы мен жетістіктері үш себеппен түсіндіріледі:
1) негізгі механизмдердің салыстырмалы түрде қарапайымдылығымен;
2) модель (үлгі) молекулалық биологияда фундаменталды рөл атқарады;
3) қазіргі заманғы эксперименталды әдістерді меңгеру: хроматография, радиоактивті изотоптар, электрофорез, рентген сәулелерінің дифракциясы, электрондық микроскопия.
Клетканы зерттеу әдістері
Клеткаларды зерттеу үшін көптеген микроскопия әдістерін қолдануға болады.
Фазалық-коптрастық, иітерфенциялық, қара негізді, оптикаларды қолдана отырып жарық микроскоптарының көмегімен тірі клеткаларды бақылаға болады.
Өлген клеткаларды әр түрлі бояулармен және клетканың белгілі бір компоненттерімен байланысатын арнайы реактивтермен бояп зерттеуге болады.
Жарық өткізетін электрондық микроскоп клеткаларды одан да жоғары деңгейде зерттеуге мүмкіндік береді: клетка органеллаларының мембраналардың, ақуыз филаменттерінің орналасуын бақылауға болады! Клеткада немесе клетка бетінде армайы макромолекулаларды жинақтау үшін электронды-тығыз белгі енгізетін реактивтерді пайдалануға болады.
Клетка мембранасының ішкі құрылысыи анықтау үшін мұздатып қатыру әдісін, ал клетка бетінің контурын үш өлшемді кеңістікте зерттеу үшін сканерлі микроскоптар қолданылады.
Жарық өткізетін электрондық микроскопты сондай-ақ ауыр металдармен боялған жеке макромолекулалардыц пәннін зерттеу үшін де қолдануға болады. Бірақ молекуладағы әрбір атомның орналасуын тек молекулалар ірі кристаллдар түзгенде ғана анықтауға болады. Бұл жағдайда ренгетен сәулслсрінің шоғы кристалл арқылы өтеді де алынған рентгеіюграммаиыц нсгізіндс кристалл түзгсн молскулалардагы атомдардыц үш кепістікті өлшемде сорналасуы есентеледі.
1611 ж. Кеилер. Күрделі жарық микроскопын жасау жолын ұсынды.
1655 ж. Р. Гук жарық микроскопының көмегімен тоз қабатының клеткаларын көрді.
1674 ж. А. Левенгук бір клеткалыларды, ал 9 жылдан соң бактерияларды ашты.
1833 ж. Браун ядрюларды сипаттады.
1835 ж. Шлейден мен Шванн клетка теориясын тұжырымдады.
1857 ж. Колликер бұлшық ет клеткаларындары митохондрияларды сипаттады.
1876 ж. Аббе дифракция құбылысының бейненің пайда болуына әсерін сараптай келе микроскопты жетілдіру жолын ұсынды.
1879 ж. Флемминг митоз кезіндегі хромосомаларды сипаттады.
1881 ж. Ретциус, Кахал - жануарлар ұлналарын, бояп қарау әдістерін ұсынды.
1882 ж. - Кох анилинді бояулардың көмегімен туберкулез, холера бактерияларын бөліп алды. Клебс, Пастер - боялған препараттар алды.
1886 ж. - Цейсс ең жетілдірідген жарық микроскопын жасады.
1898 ж. - Гольджи АgNO 3 бояп, Гольджи аппаратын сипаттады.
1924 ж. Лакассань - радиоактивті полонийді анықтау үшін алғашқы радиавтограмма әдісін жасады.
1930 ж. Лебедев - бірінші интерференциялық микроскоп, Зериике-фазалық-контрастық микроскоптыц көмегімен боялмаған клеткаларды керуге мүмкіндік берді.
1941 ж. Кунс- клеткадағы антигендерді анықтау үшін флуоресцентті бояулармен байланысты антиденелерді қолданды.
1952 ж. Номарский жарық микроскоптары үшін дифференциалды интерференциялық контрасттар жуйссін жасады.
1931 ж. Руске - бірінші сәуле өткізетін электроидың микроскопты жасады.
Өсімдіктер мен жануарлардың көптеген клеткалары белгілі бір құрамдағы қоректік орта болғанда табақшада өсуге, дамуға қабілетті. Клеткалардың әр түрлі типтері әр түрлі қоректік заттарды, соның ішіндебірнеше ақуыздық өсу факторларын қажет етеді. Жануар клеткаларының көпшілігі белгілі бір шекті бөлінуден соң тіршілігін жояды, бірақ кейде клетка дақылында спонтанды түрде шексіз ұзақ уақыт бөлінуге қабілетгі клетка варианттары пайда болады. Олардан (клеточные линии) бір бастапқы клеткадан пайда болған клондарды алуға болады. Осылай бір ақуыз бойынша мутанттық клеткаларды бөліп алуға болады. Клеткалардің әр түрлі екі типінің қосылуынан гетерокариондар (2 ядросы бар клетка) алуға болды, ал одан ең сонында гибридтік клетка (ядролары бірігіп кеткен клеткалар) түзіледі.
Гибридтік клеткаларды екі әр түрлі клеткалардың компоненттері арасындағы өзара әсеріп зерттсуде қолдануға болады. Сонымен қатар бұл әдіс белгілі бір геннің нақты қай хромосомада орналасқаның анықтауға да мүмкіпдік береді.
Микроскоп клеткалар мен ұлпалардағы органадалар мен макромолекула агрегаттарының өзара орінтасуып анықтауға мүмкіндік береді. Арнайы бояу әдістерін қолдана отырып клеткада, белгілі бір молекулаларды шоғырландыруға болады. Бірақ молекулалық денгейде зерттеу үшін биохимиялық анализ жасау қажет. Ал бұл үшін клетканы бүзу керек. Әдсіте белгілі бір типті клеткаларды бөлін алатын бастапқы материал ретінде эмбрионалдық үлпа не жанадан туылған жануарлар үлналарының клеткалары алынады. Осындай тазартылған клеткалар немесе гомогенді клетка дақылдарын ультрацентрифугалау арқылы клетканың құрам бөліктерін бөлін алын, биохимиялық ананізде колданды.
Фракцияланған клеткалық экстракттарды күрделі клетка іигілік процсетерді (мысалы акуыз синтезі немссе ДНК репликациясы) зерттеуде клеткасыз жүйе ретіде пайдаланады.
Бағаналы хроматография жолымен клеткалық экстракттағы көптеген белоктарды тазартуға болады. Бағаналы хроматографияда колданылатып матрикстер зерттелін отырған ақуыздардың биологаялық белсенділігін сақтай отырып олардың молекулалық массасы, заряды бойынша бөліп алуға мүмкіндік береді. Әдетте тазалау кезінде бірнеше осындай бағаналардан өткізілген соң белокты тек таза гомогенді күйге ғана өткізіп коймай, сонымен қатар оның амин қышқылдарының кезектесуін де анықтауға болады. Мүнда алдымен акуызды кіші пептидтерге ыдыратып, кейін сезімтал автоматтандырылған әдістердін көмегімен пептидтердегі амин қышқылдарының кезектесуін анықтайды.
Клетка ішілік макромолекулаларды зерттеу үшін молекулалардың барлық қасиеттерін - физикалық, химиялық. биологиялық пайдалануға болады. Биологиялық қасиеттерінде олардың оптикалык қасиеттері сонымен қатар биохимиялық белсенділігі бойынша анықтайды.
Клеткадағы молекулаларды зерттеудіи 2 әдісі бар:
1-радиоактвиті изтонтарды қолдану әдісі;
2-аитидсислсрді қолдану әдісі.
Екі әдісті де күрделі қоспадағы белгілі бір молекуланы анықтауда колдануға болады.
Қлеткадағы кез-келген молекуланы белглеуге болады. Оларға бір немесе бірнеше радиоактивті атомдар енгізеді. Тұрақсыз радиоактивті атомдар ыдырай отырып сәуле шығарады. Бұл зерттелін отырған молекудалардың тағдырың бақылауға мүмкіндік береді.
Клетка биологиясында радиоактивті изотоптардың пайдаланылуының мысалы - 1-ден метаболиттік жолдардың анализі және 2- клеткада жеке молекулалардың шоғарлануын радиоавтография жолымен анықтау болады.
Антидсиелер - белгілі бір биологиялық макромолекуларды шоғырландырудың қолайлы және сезімтал әдісі болып табылады. Омыртқалы жаңуарлардың денесінде миллиондаған әр түрлі антиденелер түзіледі. Олардың оркайсысында белгілі бір молекулалар тобын танитын байланысу бөліктері болады. Гибрид өдісініц кемегімен моноклональды антиленелер алуға боладіл. Клеткадағы кез-келген макромолекулаға қарсы моноклональды антиденслерді алуға болады.
Рекомбинанттық ДНҚ технологиясы клетканы зерттеуде төңкеріс жасады. Қазіргі кезде реструктуралаушы нуклсазаларды пайдалана отырып, клетка ДНҚ-нідің кез-келген бөлігін кесін алуға, клондауға және осы генетикалық материалды шексіз мөлшерде алуға, содан соң оның кезектесуін күніис бірндес жүздеген нуклеотидке дейін анықтауға болады. Осы әдіснен эукариоттардың көптеген өндерінің және қоюмдарының кодталмайтың бөдіктері анықталған.
Нуклеин қышқылдарының гибридизациясы әдісіп қолдана отырып, клеткасыз жүйеде клондалған ДНҚ-молекулаларына сәйкес иРНК молекулаларын анықтауға, бөліп алуға және трансляциялауға болады. Сондай-ақ кері бағытқа қаран да баруға - белоктан оны қозгайтын генге қарай, янғи белоктың кысқа фрагменттерінің амин қышқылдық кезектесуін анықтап, осы белокты кодтайтыи сэйкес РНҚ мен ДНҚ-мен гибридизацияланатын маманданған арнайы ДНҚ-зондтар сиитездеугсболады.
Рекомбинантты ДНҚ технологияларының мүмкіндіктері жоғары. Шексіз мөлшерде сүт қоректілердіц ақуызын синтездейтін бактериялар немесе ашытқылар жасалуы мүмкін. Бұл ақуыздың структурасы мен функциясын анализдеуге немесе медициналық мақсатта қоданылатын (вакиина немесе дәрілік препараты) ақуыздар алуға мүмкіндік берді.
ДНҚ клондау - табиғаты кез-келген ДНҚ фрагментін плазмидаға немесе бактериофатқе енгізіп, осы генетикалық элементтің бактерия немесе ашытқы клеткаларында көбейтуге мүмкіндік беретін әдіс. Клон - қажетті клеткалардың үлкен популяциясы; плазмида - клондау векторы.
Рекомбинанттық ДНҚ технологиясы көптеген әдістердің жиынтығы. Ескі, жаңа, басқа пәндерден алынған әдістер микроорганизмдер генетикасынан алынған әдістер.
Ең негізгі әдістер:
1) ДНҚ-ны арнайы рестриктуралаушы нуклсазалармен ыдырату.
2) Нуклеин қышқыдарының гибридизациясы. Ол нуклеин қышқылынын өзара комплементарлы бөліктерге байланыса отырып жоғары дәлдікпен ДНҚ мен РНҚ-ныц нуклеотидтік кезектесуін анықтауға мүмкіндік береді.
3) Белгілі бір ДНҚ фрагменттерін жылдам репликацияланатын генетикалық элементтерге (плазмида, вирустар) енгізу үшін қолданылатын ДНҚ-ны клондау.
4) Клонданатын ДНҚ фрагментіндегі нуклеотидтердід кезектесуін анықтау.
Гендік инженерия - ис-клеткасында көбейіп, зат алмасудың соңғы өнімдерін синтездеуге қабілетті in vitro генетикалық материалдың жаңа комбинацияларын бағытталған түрде жасаумеи айналысатын молекулалық генетиканыц бөлімі. Гендік инженерия 1972 жылы, П. Берг лабораториясында алғаш рет рекомбинантты (гибридтік) ДНҚ (рекомбинантты РНҚ) алмнганиан бастан пайда болды. Бұл ДНҚ лямбда фаг ДНК-мың фрагменті және маймыл вирусы SV40-тыц сакиііалы ДНК-ныц ішск таяқшасымси қосылуынаи қүралгаи еді. In vitro рек-ДНК-ны жасауда рестриктаза және ДНК-лигаза фермситтерінің маңызы зор. Рестриктазалар ДНҚ молекуласын белгілі бір жерден фрагменттерге кеседі. ДНҚ-лигазалар ДНҚ фрагменттерін біртұтас бүтін етіп тігіп шыгады. Осы ферменттерді белін алғашап соң ғана жасанды генетикалық структураларлы жасау техникалық жағынан мүмкін болды.
Рек-ДНҚ-ның селекциясының 3 жолы бар: генстикалық (маркерлар бойынша, тандамалы орталардың көмегімен), иммунохимиялық және таңбаланған ДНҚ нсмссс РНҚ-лы гибридизациялық.
Гендік инженерия әдістсрінің жедел дамуының нәтижесінде рибосомалық, траспорттық және 5S РНК, гистондар, тышқан, қоян, адамның глобиидері, коллаген, овальлбумин, адам инсулины жэне т. б. пептидтік гормондары, адамның интерфероиы және т. б. гендердін клондары алынган.
Гендік инженерия негізінде қазіргі биотехнологияның бір бағыты болып табылатын "ДНК индустриясы" деп аталатын фармацевтиканың саласы пайда болды.
1869 ж. Мишер алғаш рет ДНҚ-ны бөліп алды.
1944 ж. Эвери бактериялардағы трансформация кезінде ақуыз емес ДНҚ генетикалық ақпаратты тасымалдайтының дәлелдеді.
1953 ж. Уотсюн және Крик Франклин мен Уилкинсонның жүргізген рентгеноструктуралық анализіне негізделе отырып ДНҚ-ның қос спиральді моделін ұсынды.
1961 ж. Мармур мен Доти ДНҚ-ның ренатурациясы құбылысын ашты. Яғни нуклеин қышқылдарының гибридизациялану реакцияларының дәлдігі мен өзіндік ерекшелігін тағайындады.
1962 ж. Арбер ДНҚ-ныц рестрикциялаушы ферментнрінің бар екенді туралы мәліметтер алды, келгін оларды Натсон мен Смит бөліп алын, ДНҚ-ның нуклеотидтерінің кезектесуін анықтауда қолданды.
1966 ж. Нирснберг, Очоа, Корона генетикалық кодты ашты.
1967 ж. Геллерт ДНҚ фрагменттерін біріктіруде қолданылатын фермент - ДНҚ-лигазаны ашты.
1972-73 жж. Бойер, Коэн және Берг ДНҚ-ны клондау технологиясын жасады.
1975-77 жж. Сэнгер және Баррел, Максам және Гилберт ДНК-ның нуклеотидтерінге кезектссуін анықтаудың жылдам әдісін жасады.
Клетка эволюциясы
Төмендегі сызбанұсқада клетканың молекулалық ұйымдасу деңгейлері көрсетілген.
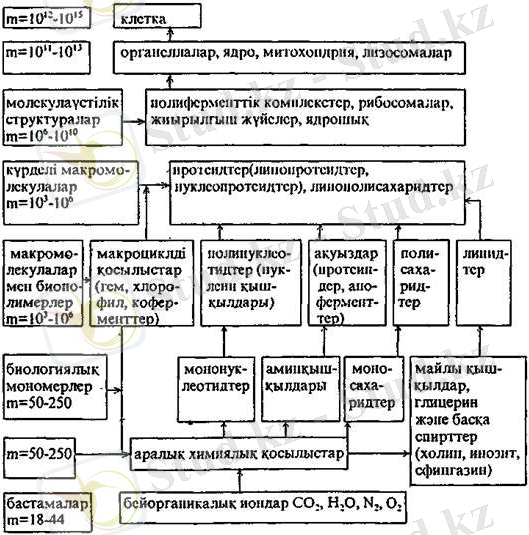
Сызбанүска 1. Клетканыи молекулярлық үйымдасу дедгейі.
Барлық тірі ағзалар клеткалардан - өте ұсақ, мембранамен қоршалған, химиялық заттардың концентрлі сулы ерітіндісімен толтырылған қуыстардан тұрады. Тіршіліктің қарапайым формалары - бұл жеке клеткалар, олар бөліну арқылы көбейеді.
Барлық ағзалар мен оларды құрайтын клеткалардың барлығы эволюциялық жолмен ортақ ата - тек болып табылатың бір клеткадан таралған деп есептеледі.
Эволюцияның екі исгізгі процесі - бұл:
1. агзаның өзінің ұрпақтарына беретін генетикалық ақпаратының кездейсоқ вариациялары;
2. тіріпілігіп сақтау мен көбеюді қамтамассыз ететіп генетикалық ақпараттың сұрынталуы.
Эволюциялық теория биологияның негізгі принципі болын табылады.
Тірінілікті жеке молекуладан генклеткалы апаларға дейін дамуын қарасырайық.
Тірі клеткалар жер бөлінде 3, 5 млрд жыл бұрын молекулалардың сиоптанды агротациясы нәтижесінде найда болды. Біздің қазіргі кездегі ағзалар мен олардағы молекулалардың құрамы, бірінші клетканың пайда болуының төмендегідей 3 алғы шартм болды ден өсентсуге мүмкіндік береді:
1) комплементарлық негіздердің жүнтасу жолымен өздігінен репликациялануға қабілетті нолимерлор (РНҚ) пайда болуы керек.
2) соның көмегімен РНҚ ақуыз синтезін бағыттайтып механизм түзілуі керек.
3) өздігінен ренликациялануға қабілетті РНҚ мен ақуыз молекулаларының қолдасын шектелген кеңістікте түйықтаған линидік мембрана түзілуі қажет еді.
Эволюциялық процестің кейнгі сатыларының бірінде тұқым қуалау заты ренде РНҚ-ны ДНҚ ауыстырды. Жердің пайда болуының бірінші миллиард жылында атмосфера болды ма жоқ па, озон қабаты болды ма, О 2 болды ма жоқ па, қандай мөлшерде (өте аз) атмосферада NН 3 , СН 4 болды ма?
Осындай жағдайда қарапайым органикалық молекулалар пайда болды (яғни С бар) . Мұны лабораториялық эксперименттер дэлендеп отыр. СО 2 , СН 4 , NН 4 , Н 2 +Н 2 О газдармның қыздырылған қоспасына электр разрядымен, улыракүлгін сәулелермен әсер еткенде кіші органикалық молекулалар түзіледі.
Цианды сутегі Н-С=N, формальдегид цианобактериялар пайда болды деген болжам болды. Олар СО 3 мен N 2 -ні фиксациялауға қабілетті болды.
Олар сулы ерітіндіде оңай әрекеттеседі. Ең бастысы, олар клетка ішілік кіші молекулалардың 4 негізгі класын түзеді: амин қышқылдары, нуклеотидтер, қанттар мен майлы қышқылдар.
Осындай жағдайда белгілі бір уақытта, белгілі бір жерде қазіргі клеткалардың құрамына кіретін қарапайым органикалық молекулалардың көпшілігі бірігуі мүмкін.
Амин қышқылдары немесе нуклеотидтер түріндегі жай органикалық молекулалар үлкен полимерлер түзе отырып ассоциациялануы мүмкін. Полипептидтер - белоктар
Полииуклеотидтер - РНК және ДНК.
Комплементарлық нуклеотидтердің маманданған жүптасуы тіршіліктің пайда болуында шешуші рөл атқарды.
Комплементарлық матрицалық көшірме механизмедері нәзік және қарапайым: әр клетканың генетикалық информациясы оның полинуклеотидтерінің негіздерінің кезектесуінде жазылған және бұл информация ұрпақтан ұрпақка негіздердіц комилементарлық жүптасуы арқылы тасымалданады.
Г-Ц Г
А-Т 'У
А-Т У
Г-Ц 'Г
РНК ДНҚ-ға ұқсас полинуклиотид, ол екі маңызды қасиеттермен сипатталады: біріпшіден - нуклеотидтердің кезсктесуі түрінде информация жазылған және ол ренликация кезіпде тасымалданады.
Екіншіден, ол ерекше өзіне тән кеңістіктік структура түзеді, бұл структура оның функциясы мен қоршаған орта жағдайларына жауабын (реакциясын) анықтайды.
Бұл екі қасиет - информациялық және функциоиалдық - эволюция процесінің қажетті алғы шарты болып табылады.
Нуклеотидтер кезектесуі - генотип.
Кеңістікте орналасуы - фенотип, яғни генотиптің көрінісі.
... жалғасыСіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz