Везикулярлы және трансмембраналық тасымалдау: механизмдері, иондық арналар және сорғыштар

Везикулярлы тасымалдау
Жоспар
1. Жалпы мәліметтер
2. Ұсақ молекулалы заттардың тасымалдануы
3. Заттардың тасымалдануының кейбір жүйелері (сорғыштар және арналар)
4. Катиондық арналар және Н-холинорецепторлар
5. Көлденең жолақ бұлшықет ұлпасында Са 2 + иондарының тасымалдану жүйесі
6. Бүйректе глюкозаның тасымалдануы
7. Мембрана арқылы түйіршіктердің және ірі молекулалы қосылыстардың тасмалдануы
1. Жалпы мәліметтер
Жасуша цитоплазмасының маңызды қызметтерінің бірі - заттар ағынын қамтамасыз ету болып табылады. Заттар ағыны дегеніміз: біріншіден -жасуша ішінде, кедір-бұдыр эндоплазмалық торда синтезделген ақуыздардың органеллалар арасьгнда әрлі-берлі тасымалдануы; екіншіден -кептеген жасушалар мен үлпаларда синтезделген пептидтік гормондардың, асқорыту ферменттерінің, антиденелердің, есу факторларының және басқа да секреторлық молекулалардың жасуша сыртына шығарылуы; үшіншіден-сыртқы ортадан жасушаға үнемі өртүрлі заттардың еткізілуі.
Заттардың жасушаішілік-везикулалық тасымалдануының әмбебап және тиімді құралы болып тасымалдану (мембрана) көпіршіктері (липосомалар, мицеллийлар) арқылы секреторлық механизм негізінде тасымалдануы болып табылады.
Везикулалық тасымалдануда тасымалданатын ақуыздар мен липидтер кепіршік (липосома, мицелла) қабырғасын (мембранасын) құрастырады, ал оның қуысында басқа органеллаларға арналған не жасушасыртына шығарылатын «жүк» молекуласы болады.
Жасушаішілік везикулалық тасымалдау эвдоплазмалық ретикулум (ЭПТ) мембранасынан басталады. Бұл жерде ақуыз молекуласының гликозилденуінің алғашқы кезеңдері етеді. Содан кейін ақуыз молекулалары тасымалдау көпіршіктеріне іріктелініп, Гольджи кешенінің цис-полюсіне өтеді. Гольджи цистерналарыңда ақуыздардың гликозилденуі әрі қарай жалғасады, ал Гольджидің транс-полюсі мен транс-торларыңда ақуыздың гликозилденуі толығымен аяқталады. Сонымен қатар олар фосфорланады жөне сульфатганады. Гольджи цистерналарынан ақуыздар жиекті көпіршіктер арқылы өтеді. Гольджидің транс-торларында толық модификацияланған ақуыздар нақтылы органеллаларға тасьшалдану үшін тасымал көпіршіктеріне іріктелінеді. Гольджи кеш^енін тастап шыққаннан кейін, ақуыздар алғашқы лизосомаларға, конститутивтік көпіршіктерге және секреторлық гранулаларға үлестіріледі.
Заттардың цитоплазмалық мембрана (плазмолемма) арқылы сыртқа шығарылуын (экзоцитоз) не жасуша ішіне өткізілуін (эндоцитоз) трансмембраналық тасымалдану деп атайды. Ол өте күрделі құбылыс және әртүрлі жасушаларда түрліше жолдармен жүзеге асады, сол сияқты, әртүрлі заттарда түрліше әдістер арқылы өткізіледі.
2. Ұсақ молекулалы заттардың тасмалдануы
Ұсақ молекулалы заттардың биомембрана арқылы өткізілуінің 3 жолы белгілі:
а) жай диффузия;
б) жеңілдетілген диффузня;
в) белсенді тасымалдану.
Жай диффузия - өздігінен, ешбір көмексіз, затгардың концентрация градиенті (жоғары концентрациядан төменгі концентрация) бағытында мембрана арқылы өтуі.
Мұндай әдіс арқылы кіші молекулалы гидрофобтық органикалық крсылыстар (май қышқылдары, зәр қышқылдары) және ұсақ, бейтарап молекулалар (Н 2 0, С0 2 , 0 2 ) өтеді.
Мембрана арқылы шектелген қуыстардың (органеллалар) концентрация айырмашылығы көбейген сайын диффузия жыддамдығы да пропорциональ өседі, ал олардың концентрациясы теңессе диффузия тоқталады.
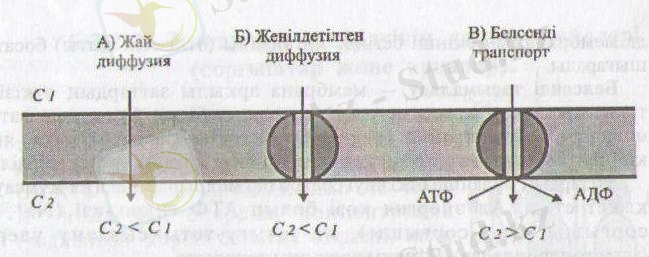
1-сурет. Ұсақ молекулалы заттардың өткізілу жобасы (Мушкамбаров, Кузнецовтан, 2003)
Жеңілдетілген диффузия бұл өдісте де заттар ездерінің концентрация градиенті бағытында мембрана арқылы етеді, яғни жоғары концентрациядан теменгі концентрация бағытында, бірақ бұл құбылыс ездігінен жүзеге аспайды, ал ерекше тасымалдау ақуызы-транслоказаның кемегімен жүреді.
Транслоказалар - ездері еткізетін заттарға азды-кепті сай болып келетін интегралдық ақуыздар. Мысалы, эритроцит мембранасындағы аниондық арналар (канаддар), қозғыш жасушалар плазмолеммасындағы К+ арналары (канадцары), саркоплазмалық ретикулум мембранасындағы Са+-арналары (каналдары) .
Транслоказалар арқылы жай диффузия жолымен ете алмайтын заттар ғана еткізіледі, бірақ кейде, кейбір заттар, жай диффузия және жеңілдетілген диффузия арқылы да етеді, мысалы судың (Н 2 0) бүйрек араншықтары және секреторлық эпителий жасушалар мембранасы арқылы етуі. Аталған мембраналарда су молекулаларының диффузиялану қарқынын арттыратын транслоказа-аквапорин деп аталатын ақуыз болады.
Транслоказалар-бірнеше бөлшектерден (субъединицалардан) түрады, олардың әрекет ету тетіктерінің (механизмінің) бірнеше түрлері болуы мүмкін:
1) Транслоказа белшектері (субъединицалары) арасында белгілі бір өлшемді заттарды ғана еткізетін және барлық уақытта ашық болатын гидрофильдік арна (канал) болады;
2) Транслоказа арнасы барлық уақытта ашық болмайды, оның ашылуы үшін транслоказа белшектері бетімен арнайы лиганданың байланысуы қажет;
3) транслоказаларда ешқандай арна болмайды, олар лигандамен (еткізілетін зат) байланысып, мембрана жазықтығында 180" айналады да мембрананың екінші бетінде лиганданы (өткізетін затты) босатып шығарады.
Белсенді тасымалдау - мембрана арқылы заттардың өткізілуі транслоказалар көмегімен жүзеге асады, бірақ бұл кезде заттар олардың концентрация градиентіне қарама-қарсы бағытта, яғни концентрациясы аз ортадан концентрациясы жоғары ортаға өткізіледі.
Заттардың бұлайша өткізілуі белгілі бір мөлшерде энергия жүмсауды қажет етеді. Ал энергия көзі болып АТФ гидролизі (Ка + , К + сорғышы, Са 2 + -сорғышы), не тотығу-тотықсыздану үдерісі (митохондрияларда) -Н + ионы сорғышы саналады.
Белсенді тасымалдауды энергиямен қамтамасыз етудің тағы бір тетігі-концентрация градиенті бағытында өткізілетін бір заттың-У концентрация градиентіне қарама-қарсы бағытта өткізілетін екінші бір затпен-Х, қабаттасып өткізілуі. Бұл жағдайда, У өткізілуінде бөлінетін энергия мөлшері Х-өткізуге жүмсалатын энергиядан артық. болуы қажет.
Бұл құбылыстың 2 нүсқасы белгілі: симпорт және антипорт.
Симпорт кезінде транслоказа екі затты (У, Х) бір бағытта өткізеді, оның біреуі-У концентрация градиенті бағытында диффузияланып екінші затты-Х, өзімен бірге ілестіріп өткізеді. Мысалы, бүйрек арнашықтарьшан глюкозаның реабсорбциялануы (кері сорылуы) осындай тетік (механизм) арқылы Иа + ионымен бірге симпортталынады. Егер симпортқа қатынасатын заттардың екеуі де иондар болатын болса, олар түрліше зарядталған болуы қажет.
Антипорт - транслоказа арқылы заттардың (У, Х) қарама қарсы бағыттарға өткізілуі, яғни У молекуласы Х-молекуласымен алмастырылады.
Эукариоттарда антипорт өте сирек кездеседі.
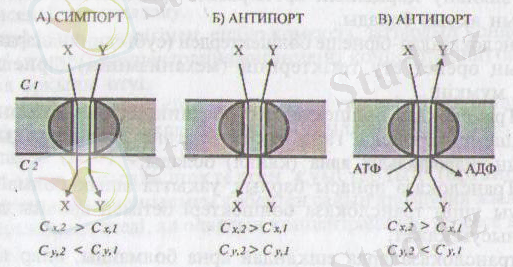
2-сурет. Екі заттың бірлесіп тасымалдану жобасы (Мушкамбаров, Кузнецовтан, 2003)
3. Заттардың тасымалдануының кейбір жүйелері (сорғыштар және арналар)
1) Nа + , К*-сорғышы немесе Иа + , К + -т2уелді АТФ-аза-2а -ширатпадан, 2р-құрылымнан түратын интегралдық ақуыз. Ол АТФ энергиясын пайдаланып № + және К + иондарын олардың концентрация градиентіне қарсы бағытқа өткізеді, яғни № + ионын-жасушадан сыртқа, ал К + ионын-жасуша ішіне.
Осы сорғыш қызметінің арқасында Иа + ионының концентрациясы жасуша сыртында, ал К + ионының концентрациясы жасуша ішінде айтарлықтай жоғары болады, яғни иондардың жасушаішілік және жасушааралық ассиметриялық үлестірілуі орын алады.
АТФ гидролизінде бөлініп шыққан фосфат тобы ақуызға (транслоказа) беріледі және оның конформациясын өзгертеді, нәтижесінде № + иондары толтырылған қуыс мембрананың екінші беті жағында ашылады. Иондараралық электрлік кері тебілу күші иондардың (№ + ) жасуша сыртындағы ортаға, оның концентрациясының жоғары болуына қарамастан, бөлініп шығуын тудырады.
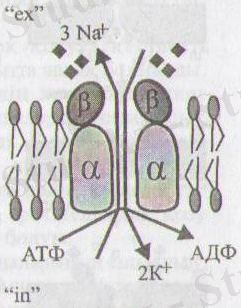 "
"
4-сурет. N +, К+-сорғышы (Мушкамбаров, Кузнецовтан, 2003)
№+ ионының орнына соргыш қуысына 2 К + ионы толтырылады. К + ионының байланысуы транслоказаны фосфорсыздандырады, бұл оның бастапқы конформациясына қайтып кЪлуіне ықпал етеді, нәтижесінде оның қуысы қайтадан мембрананың ішкі беті жағында ашылады да, К + ионы жасуша ішіне босанып шығады.
2) К + арнасы (ішкі диаметрі-ОЗнм), көптеген жасушалар
плазмолемасында кездеседі және үнемі ашық болады. Осының арқасында № + К + сорғышы қызметі нәтижесінде пайда болған өте жоғары концентрация градиентіне байланысты, К + ионының біршама иондары осіы арна арқылы жасушадан тыс ортаға қайтып келеді. К + ионының шамалы ғана мөлшерінің шығарылуы (әрбір 1000 нм 2 мембрана бетінде небәрі 6 К + ионы шығарылады) мембрана беттерінде, концентрация градиенті энергиясымен теңестірілетіндей, потенциалдар айырмашылығы қалыптастырады, ол -75 мв тең. Сондықтан да динамикалық тепе-теңдік орнап К + ионының арна арқылы әрі қарай шығарылуы тоқтайды.
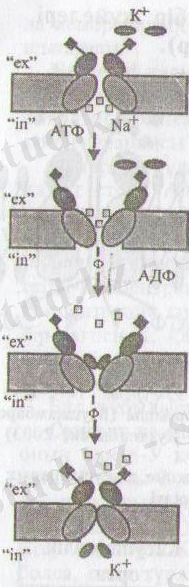
5-сурет. П+, К+
сорғышының әрекет
егу тетігі (механизмі)
(Мушкамбаров,
Кузнецовтан, 2003)
Нәтижеде осы иондардың жасушаішілік және жасуша сы концентрациялары езгермейді, бірақ жасуша трансмем потенциалға ие болады. Бұл кезде плазмолемманың сыртқі ал ішкі беті-теріс зарядталған болады.
3) № + арнасы (ішкі диаметрі-0, 55нм), тек қозуға қаоілет мембраналарда ғана болады және ол барлық уақытта ашық болмайды. Иа^арнасы-нерв жасушаларының, миоциттердің және бұлшықет талшықтарының, сперматозоидтардың, сезім мүшелерінің сенсорлық жасушаларының плазмолеммаларында кездеседі. Бұл жасушаларда № + арнасының тығыздығы түрліше болады, яғни плазмолемма бетінің 0, 2-1%-ын, яғни 1мкм2-та 50-200 арнаға дейін кездеседі.
№ + арналарының К + арналарынан ең басты ерекшелігі тек қозу кезінде «ашылып», тыныштық күйінде жабық болуы.
Мембраналардың белгілі бір учаскесінде № + арналарьшың ашылуьш, осы учаскеде трансмембраналық потенциалдьщ 50мв-қа дейін төмендеуі инициациялайды. Потенциалдың мұндай төмендеуі мембрананың көрші учаскесінің қозуының салдары болып табылады.
Ашылған Иа + арналары арқылы Ма + иондары жасуша ішіне қарай концентрация градиенті бағытында ағылады. Осылайша жасуша сыртында К + арналары арқылы қалыптасқан оң зарядтар қоры азаяды. Сондықтан да потенциаддар айырмашылығы өрі қарай төмендеп, жабық № + арналарының ашылуын индукциялайды.
Есептерге сәйкес, әрбір арна арқылы бір импульста жасушаға 500 № + ионы өтеді, бұл олардың жасушаішілік концентрациясының айтарлықтай жоғарылауына алып келмейді, тек трансмембраналық потенциалды едәуір өзгертеді.
Бір миллисекунд ішінде потенциалдар айырмашылығы теңесіп, нөлге дейін жетіп қана қоймай, сәлден кейін ол оң зарядталған болады. Бұл оң зарядталған иоңдар артықшылығы-ның жасуша сыртында емес, жасуша ішінде болуына алып келеді, себебі: № + иондарының жасуша ішіне ену қарқыны К + иондарының жасушадан шығарылу жылдамдығына қарағанда әлде қайда жоғары болады.
Потенциалдар айырмашылығының оң көрсеткішке ие болуы, ез кезегінде, Ыа + арналарының жабылуын индукциялайды, сондықтан да мембрананың осы жеріңдегі потенциалдар айырмашылығы үнемі ашық болатын К + араналар есебінен, тез арада қалыпты күйіне (-75 мВ) келтіріледі.
Сонымен, Ыа + арналары-мембрананың, синапстан тыс қозу және мембрана арқылы сигналдары өткізу үдерістерінде маңызды рөл атқарады
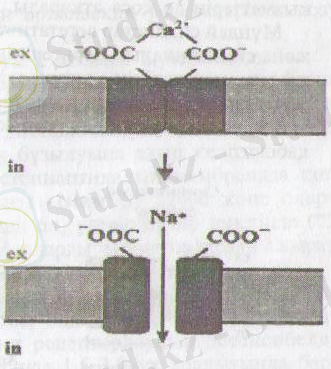
6-сурет. Са 2 + иондарының N8*арнасының күйіне әсерлері
(Мушкамбаров, Кузнецовтан, 2003)
Nа + арналарының ашылуы сыртқы ортадағы Са 2 + ионының концентрациясына тікелей байданысты болады. Егер сыртқы ортада Са 2 + ионы концентрациясы жоғары болса, онда Ыа + арналары өздерінің карбокситоптарымен (СОО-) Са 2 + ионымен байланысады, ал Са 2 + иондары арнаның ашылуына кедергі келтіреді (97-сурет) . Нәтижесінде Са 2 + ионы мембрананың қозуын төмендетеді.
4. Катиондық арналар және Н-холинорецепторлар
... жалғасыСіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz