Гендік инженерия: ген алу әдістері, рестрикциялық ферменттер және векторлар

Жоспар:
1 Гендік инженерия
2 Гендік инженериядағы генді алу әдістері
2. 1 Генді тікелей бөлу әдісі
2. 2 Генді синтездеудің химиялық әдісі
2. 3 Ферменттік (энзимдік) әдіс.
3 Хромосома және геном деңгейіндегі генетикалық инженерия
4 Ген инженериясы негізінде биотехнологиялық өнімдер алу
5 Ген инженерия әдістерінің медицинада қолданылуы
5. 1Қан факторларын алу
5. 2. Ген инженерия әдістерінің иммуномодуляторлар алуда қолданылуы
5. 3Вакциналарды өндіру
Гендік инженерия
Биотехнология 70-жылдары ғылымда жаңа серпін алды және гендік (генетикалық) инженерияны молекулалық және клеткалық инженерия белгілі бір мақсатпен жасанды айқын қасиеттері бар генетикалық материалдарды алдын ала құрастырып, оларды басқа клеткаға енгізіп, көбейтіп, зат алмасу процесін өзгеше жүргізу. Бұл әдіспен организмдердегі тұқым қуалайтын информацияны көздеген мақсатқа сай өзгертіп, олардың геномдарын белгілеген жоспармен қайта құруға болады. Бұл биотехнология құралы болып табылады және молекулалық биология, биохимия, клеткалық биология, генетика, микробиология және т. б биологиялық ғылымдардың мәліметтерін пайдаланады. Гендік инженерияның құрылуының нәтижесінде генетикалық ақпарат тірі организмдегі тұқым қуалаушылықтың басқару әдістерін дамыту туралы жаңалықтардың сериясы жоқ болуы мүмкін емес.
Осындай жаңалықтарды жатқызуға болады:
1. Оқшауланған «химиялық» таза гендердің алынуы. Бастапқыда тиісті жасушалар гендерді оқшаулауға тырысты, бірақ содан кейән өзінің құрылымын біле тұра, химиялық және молекулалық -биологиялық синтез арқылы алу оңай болып шықты. Содан кейін гендерді клеткаға кірістіру әдісін жасау керек болды. Гендерді цитоплазмаға кірістіруді үйреніп қана қоймай, ДНҚ клеткасының молекуласын құрастырып, жаңа ақпарат биосинтетикалық ақпаратты оқи алатындай, ақуызды өндіргенде және бөлінген кездегі гендерді шығару.
2. Гендердегі «жазылған» биологиялық ақпаратты «оқу» әдістерін жасау. Бұл жұмысты ағылшын ғалымы Ф. Сенгер және америкалық ғалым У. Гилберт жасады. 1980 жылы ғалымдар осы жұмыстың арқасында химиядан Нобель сыйлығын алды.
3. Клеткадағы генетикалық ақпараттың құрылымы мен қызметінің ашылуы.
4. Гомологты және гомологты емес рекомбинантты ДНҚ механизмдерінің ашылуы.
5. Репликацияға қабілетті бактериалдық клеткадағы хромосомаішілік сақиналы ДНҚ табылуы. Олар 50 жылдардың басында табылды және «плазмид» деген атқа ие болды. Ф. Гриффит бактериалдық трансформацияның пайда болуын ашқаннан кейін, бактерияларға гендерді кірістіру технологиясы жасалды. Бұл құбылыстың пайда болуында қарабайыр жыныстық процесс жатыр, бактериядағы хромосомалық емес ДНҚ, плазмиданың кішігірім фрагменттері алмасады.
6. Плазмидалар автономды репликациялық хромосомалар және клеткада бірнеше данада сақталуға қабілеті бар. Плазмидалар гендік анықтауыштар арқылы айырылады. Плазмидалардың аз мөлшерінің өзі бұзылмаған, туғандай жағдайда клеткадан бөлініп шығады.
7. Рестриктаза және лигазаныі ашылуы. 1970 жылы америкалық ғалымдар С. Келли және Г. Смит өздерінің әріптестерімен, алғаш рестриктазаны бөліп алды - қатаң орындарда ДНҚ гидролитикалық бөлшектенбеуін тудыратын фермент. 60 жылдардың аяғында швейцариялық ғалымдар С. Линна және В. Арбер осындай ферменттерді өздерінің тәжірибесінде дәлелдеді. Рестриктаз гендердің көмегімен әр түрлі комбинациялармен біріктіруге болады. Қазіргі уақытта көптеген ферменттер гендік инженеряда қолданылады: ревертаза, ДНҚ полимераза, терминалды трансфераза, нуклеаза.
70 жылдардың басында гендік инженерияның теориялық алғышарттарының негізгі принциптері және тірі организмдегі нуклеин қышқылдарының және белоктардың қызметтері құрылды.
Гендік инженериядағы тұңғыш тәжірибені 1972 ж. американ биохимигі Т. Берг (Нобель сыйл. лауреаты) іске асырды. Ол маймылдың онноген вирусы SV-40-тың толық геномын, бактериофаг - L геномының бір бөлігін және Е. Colі бактериясының галактоза генін біріктіру арқылы рекомбинантты (гибридті) ДНҚ алды. Гендік инженерияның жұмысы мынадай кезеңдерден тұрады:
1. басқа организмге көшірілетін құрылымдық генді алу;
2. оны вектордың құрамына енгізу, яғни рекомбинанттық ДНҚ жасау;
3. рекомбинанттық ДНҚ-ын өсімдік клеткасына тасымалдау;
4. өсімдік клеткаларында бөтен ДНҚ-ның экспрессиясын талдау;
5. геномы өзгерген жеке клеткалардан регенерант өсімдігін алу.
1973 жылы амарикалық зерттеушілер С. Коэн және Э. Чанг бақаның ДНҚ бактериалдық плазмидасын құрастырған. Кейін осы құрастырылған плазмиданы бактерияның клеткасына орналастырып қойылды, сол бақаның ақуызын синтездейді және бақаның ДНҚ өзінің ұрпағын беріп отырады.
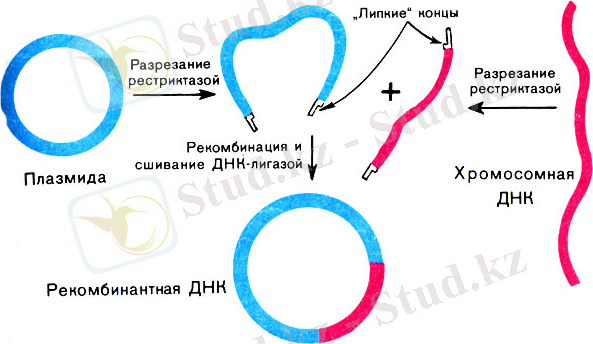
Сурет. 2. Гендік инженерияның негізгі схемасы
Гендік инженерия генетикалық ақпараттың функционалдық механизмдерін реттейді, ал басқа әдістермен мүмкін емес. Сонымен қатар практикалық әдістерді алға қояды, ал кейбіреулері қазіргі уақытта шешілмеген. Бактериалдық синтез арқылы көптеген препараттарды алуға болады. Бірақ бұл технология әлі күнге дейін жасалып жатқанмен, ауыл шаруашылығында және медицинада көптеген жетістіктерге жетті. Мысалы: медицинада көптеген вакциналар алуда, ал ауыл шаруашылығында рекомбинантты ДНҚ көмегімен өсімдік культуралары алынды. Олар кеуіп кетуден, суықтан, құрт-құмырсқадан гербицидтерге төзімді.
Гендік инженерия әдісімен бірнеше түрлі вакцина алынды және соның арқасында СПИД-ке қарсы вакцина алынуда. Рекомбинантты ДНҚ көмегімен көптеген мөлшерде адамның өсу гормоны алынуда, ол гипофиздік ергежейлікті емдеуде жалғыз құрал болып табылады.
Гендік инженерияның жетістіктерінің бірі - диагностикалық жиынтық болып табылады. Трансгенді организмнің ашылуы көптеген микроорганизмдердің жаңа штамдары, трансгенді жануарлар мен өсімдіктер, адамға қажетті мүмкіндіктер туғызуда.
... жалғасыГендік инженериядағы генді алу әдістері
1. Генді тікелей бөлу әдісі
2. Генді синтездеудің химиялық әдісі
3. Ферменттік (энзимдік) әдіс.
1. Генді алудың үш әдісі бар: табиғи генді тікелей бөлу (сирек мүмкіндік), химиялық және ферменттік синтез. Табиғи генетикалық материалдан - ДНҚ-дан арнайы ферменттердің (рестрикциялық эндонуклеазалардың) көмегімен қажет ген «кесіліп» алынады. Бұл әдістің елеулі кемшіліктері бар. Біріншіден, ДНҚ-дан қажет генді танып, кесе алатын ферментті таңдап алу қиын. Фермент тенді әрқашанда наңтьі шекарасыпда емес әр қилы үзеді: не геннің екі жағынан артық нуклеотидтерді үзуі, не түгел үзбеуі мүмкің, мұндай ДНҚ фрагментерінің қызметі жеткіліксіз, сондықтан оларды пайдалану мүмкін болмайды. Екіншіден, эукариоттық организм геномының экзон-интрондық құрылысы, олардың гендерін бактерияларға енгізгенде функциялық тұрғыдан қиындық туғызады, өйткені бактериялық клеткада сплайсинг процесі (ин-трондардың кесілуі) өтпейді. Үшіншіден, егер ген барлық ДНҚ-ның аз ғана бөлігін құраса, онда оны бөлу мен анықтауда елеулі қиындықтар пайда болады. Сондықтан бұл әдіс негізінен генетикалық эксперименттер талабына сәйкес вирус пен бактериялардың генін бөлуде қолданылады.
2. Генді-синтездеудің химиялық әдісі. Бұл әдістер белоктың немесе полипептидтің бірінші құрылымы ( амин қышқылдар қатары) белгілі болса, оның генінің нуклеотидтер қатары химиялық жолмен синтезделеді.
Берілген нуклеотидтер тізбегі бойынша ДНҚ - синтездеу әдісін 1969 жылы, ген инженериясының дәуірі басталмаған кезде-ақ, Г. Корана ұсынған болатын. Ашытқы тРНҚ-ның гені осылай синтезделді. Бұл ген 77 н. ж. құралған. Алдымен, ұзындығы 5-12 нуклеотидтерден тұратын ДНҚ-ның қысқа фрагменттері синтезделді, онан соң олар арнайы ферменттің (лигаза) әсерімен бір-біріменқосылды. Алайда, алғаш синтезделген бұл генді ішек таяқшасының клеткасына енгізген кезде жұмыс істей алмады, өйткені онда реттеуші элементтер - промотор және терминация бөліктері жоқ болатын. Кейін, 1976 ж. Г. Корана қызметкерлерімен тирозиннің тРНҚ-ының генін синтездей алды. Геннің ұзындығы 126 н. ж. тең болды, оған 52 н. ж. құралған промотор, 21 н. ж. - терминатор және ұштарына тетрануклеотидтер (ААТТ және ТТАА) жалғанды. Нәтижесінде, осы генді бактериофаг арқылы Е. Со1і клеткасына енгізгенде олар өз функциясын көрсетеалды.
Қазіргі уақытта Г. Корана өңдеген әдістер бірқатар жасанды гендерді синтездеу үшін қолданылады. Осындай жолмен адамның - инсулин және интерферон, адам мен жануарлардың - энкефалин және бардикинин гормондарының функциялы гендері синтезделді.
Қазіргі кезде гендерді автоматты синтездейтін құралдар бар. 1980 ж. Итакура алғаш өзінің құрамында алдын ала берілген тізбек бойынша 6 сағ. ішінде 12-мүшелік олигонуклеотидті синтездеуге қабілетті автоматтың жұмысын компьютер бақылады. Мұндай жабдықтардың 1982 жылғы бағасы 36000-39500 долларға тең болатын. Егер 1979 жылы 120 н. ж. генді синтездеу үшін екі жыл керек болса, ал 1981 ж. бұл мақсатқа үш-аң күнде жетуге болады.
3. Генді синтездеудің ферменттік (энзимдік) әдісі. Генді синтездеудің үшінші әдісі кеңінен таралған және кейін рекомбинанты ДНҚ күйінде бір, кейде көп клеткалы организмдерде көбейетін гендердің негізгі шығу көзі болып саналады. Оның мәні ферменттік синтез арқылы генді алудан тұрады және төмендегіге саяды. Ең алдымен, клеткалардан (мұнда, қажет геннің активтілігі жоғары болуы керек) информациялық (матрицалық) РНҚ (иРНҚ) молекулаларын бөліп алады, олардың арасында ген коделейтін иРНҚ бар, міне осы иРНҚ-ны бөлу қажет. Бұдан кейін кері транскриптаза (ревертаза) ферментінің көмегімен бөлінген иРНҚ-да комплементарлы ДНҚ (кДНҚ) синтезделеді. Кері транскрипция процесінде матрицалық иРНҚ-да кДНҚ-ның синтезі басталуы үшін иРНҚ-ның 3’- ұшына комплементарлы «ашытқы, »-олигонуклеотид (қысқа тізбек) қажет. Жаңа кДНҚ тізбегі синтезделуі үшін тағы Мg иондары мен керек.
Жаңа кДНҚ синтезделіп біткеннен кейін, оны тазалап, ДНҚ-ның екінші тізбегін синтездеу үшін матрица ретін де пайдаланады. Ол үшін ортаға ревертазаны немесе ДНҚ-полимеразаны (бактериялардан алынған) қосады. ДНҚ-ның екінші тізбегінің синтезі басталуы үшін олигонуклөотид (ДНҚ-ның 3’- ұшына комплементарлы) керек, әйтпесе кДНҚ-ның 3’- ұшы белгісіз себептермен шпилькалы кұрылым түзейді, осы құрылым екінші ДНҚ-ның синтезін инициациялай алады. ДНҚ-ының екінші тізбегі синтезделіп болғаннан кейін, оның алғашқы кДНҚ-ымен қосылған бөлігі (шпилькалы) нуклеаза 51 ферментінің әсері арқылы ажырайды, нәтижесінде қос тізбекгі ген алынады, оңы қос тізбекті комплементарлы ДНҚ (қт-кДНҚ) деп атайды. РНҚ жіпшелері сілтінің әсерімен гидролизденеді, ал олигонуклеотидтер аталған нуклеазаның әсерімен ыдырайды.
Кері транскриптаза ферменті арқылы синтезделген гендерді бактерия клеткасына енгізбестен бұрын, оларға реттеуші элементтер жалғайды. Прокариоттарда сплайсинг өтпейтінін ескерсек, онда генді ферменттік жолмен синтездеу үшін матрица ретінде гяРНҚ-ны емес, жетіл-ген иРНҚ пайдалану керек.
Рестрикциялық ферменттер (эндонуклеазалар)
1. Рестриктизалалық ферменттердің ашылуы
2. Рестриктизалалық ферменттердің типтері
3. Рестрикция -модификация жүйесі
1. Ген инженериясының немесе рекомбинантты ДНҚ технологиясының қалыптасуы ерекше ферменттер класы - эндонуклеазалардың немесе рестриктазалардың ашылуымен және қолданылуымен байланысты. Рестрикция ферменттерін 1972 ж. В. Арбер бактерия клеткасында ашты. Барлық бактериялар қос тізбекті ДНҚ-ның белгілі нуклеотидтер бөлігін үзе алатын бір немесе бірнеше рестриктазаларды синтездейді. Рестрикция ферменттерінің клеткадағы қызметі - бактерияға енген бөтен ДНҚ молекуласынан қорғау, рестрикция тоқтату деген мағынаны білдіреді. Рестрикция ферменттері фагтың нуклеин қышқылын үзіп, оның бактериялық клеткада кебеюіне жол бермейді.
Рестрикция ферменттерін синтездейтін клетканың ДНҚ молекуласы эндонуклеазалардың әсерінен үзілмейді, өйткені бұл клеткалар модификациялау ферменттерін синтездейді, олардың әсерінен ДНҚ-ның құрылымы модификацияланады (өзгереді) - ДНҚ-ның репликациясы кезінде рестриктаза танитын бөліктерге метил тобын (СНз) - жалғайды. Метилденген ДНҚ-ны рестриктаза үзе алмайды. Мұны бактериялардың өзіндік бір иммундьқ жүйесі деп санауға болады. Дегенмен, кейде бактерия клеткасына енгең кейбір бактериофагтардың нуклеин қышқылы метилденіп кетеді де, бактериялар осы фагтың әсерінен лизиске ұщырайды. Осы құбылысты-Нобель сыйлығының 1973 ж. лауреаттары - X. Смит, Д. Ңатанс және В. Арбер ашты. Рестрикциялау және метилдеу фөрменттері клетканың модификация-рестрикция жүйесін (МR) - құрайды. 2. Рестриктазалардың негізгі үш типі: I, II және, III белгілі. Барлық рестриктазалар қос спиралды ДНҚ-ның белгілі нуклеотидтер қатарын дәл тани алады. Алайда, рестриктазалардың I типі белгілі нуклеотидтер қатарын танығанымен, оны әр қилы нүктелерде үзеді, ал рестриктазалардың II және III типтері ДНҚ-ның нақты бөліктерін дәл үзе алады. Эндонуклеазалардың I және ІІI типтер құрылымы күрделі және екі модификациялық және АТФ-ға тәуелді эндонуклеазалық активтіліктері бар. Рестрликтазалардың II типі екі жеке белоктардан құралған: рестрициялаушы эндонуклеаза жэне модификациялаушы метилаза. Аталған себептерге байланысты ген инженериясында тек қана рестриктазалардың II типі пайдаланылады. Екінші жағынан; рестриктазаның II типі ғана бірдей нуклеотид тізбектерінің фрагменттері бар ДНҚ препараттарын алуға және әр түрлі геномдардан алынған фрагменттерден химерлі ДНҚ молекуласын құрастыруға мүмкіндік туғызады.
Рестриктазалардың ІI типінің басым көпшілігі 4-6 н. ж. құралған палиндромды ДНҚ тізбектерін тани алады. Мысалы, Вас. subtilis бактериясының ВsuR1 деп белгіленетін рестриктазасы ДНҚ бөлігін төмендегідей үзе алады:
Әр тізбекті 3’- ұшына 5′ бағытына қарай көз жүгіртсе, олар бірдей оқылады (ЦЦ ГГ), міне осындай тізбек палиндром деп аталады. Натижнсінде ДНҚ-ның қос тізбегі симметриялы үзіледі. (сілтемемен (стрелка) көрсетілген), сақиналы ДНҚ түзу құрылымға айналады. ДНҚ молекуласының. Соңында жабысқақ ұштар пайда болады, сондықтан олар ешқандай қосымша өңдеусіз бір-бірімен қайтадан қосылуға қабілетті болады. Қазіргі кезде рестриктазалардың II типінің 500-ден астам түрлері белгілі және олар ДНҚ-ны бір-бірінен өзгеше 120 жерден үзе алады. 1973 ж. X. Смит және Д. Натанс рестрикция-модификация жүйе (МК) ферменттерін белгілеу номенклатурасын ұсынды. Көпшіліктің мақұлдауы бойынша туыстың бірінші әрпі және түрдің алғашқы екі әрпінен құралған үш әріп ферменттің шығу көзін көрсетеді. Эндонуклеазаларды - R, метилазаларды - М символдарымен белгілейді. Егер клетканың бір типінен екі және одан көп рестрикция ферменттері бөлінген болса онда оларды рим цифрларымен (I, II, ІІІ т. с. с) нөмірлейді. Осы номенклатура сүйеніп, Е. Соlі рестриктазасы ЕсоRІ, ЕсоRІІ және т, т., ал Вас. subtilis рестриктазасы ВsuRI т. с. с. болып
белгіленеді.
3. ДҢҚ тізбегінің үзілуі не симметрияның өсі бойынша жүруі мүмкін, онда «доғал» ұшты фрагменттер түзіледі (мысалы, Ваll немесе ВsuRI рестриктазалары), не симметрия өсінен біршама қашықтықта өтуі мүмкін, онда шығыңқы «жабысқақ» 5′ (ЕсоRІ, Ніnd III- және т. б. рестриктазалар) немесе 3′ (РstІ рестриктазасы) ұштары бар фрагменттер түзіледі.
Әр түрлі рестриктазалардың II типінің ішінен ДНҚ-ның бір нуклеотидтер қатарын танитын екі немесе бірнеше ферменттер кездеседі. Осындай жұп немесе топ эндонуклеазалар изошизомерлер деп аталады. Изошизомерлердің екі түрін ажыратады: нағыз изошизомерия - ферменттер белгілі бір нуклеотидтер қатарын танып, ДНҚ-ны бір нүктелерде үзеді және жалған изошизомерия - ферменттер бір нуклеотидтер қатарын тани алға-нымен, оларды әр түрлі үзеді.
Рекомбинантты ДНҢ техникасы үшін алуан түрлі рестрикция ферменттерінің ішінен өздеріне комплементарлы «жабысқақ» ұштары бар фрагменттер түзейтін рестриктазалар бағалы болып саналады.
Ген инженериясының векторлық (тасымалдаушы) молекулалары
1. Векторлар және оларға қойылатын талаптар
2. Вектор түрлері1. Ген инженериясының маңызды мақсаты синтезделген немесе бөлінген генді клеткаға тасымалдау саналады. Бұл мақсат ДНҚ-ның векторлық молекулалары немесе қысқаша векторлар (тасымалдаушы немесе тасығыш) көмегімен іске асады.
Вектор деп бөтен генетикалық материалды (ДНҚ фрагментін) клеткаға (реципиенттің) тасымалдауға қабілетті ДНҚ молекуласын айтады. ДНҚ молекуласын векторсыз, мысалы, бактериялық клеткаға енгізсе, онда оларды бактериялық ферменттер ыдыратып жібереді. Кейбір жағдайда ДНҚ сақталуы мүмкін, бірақ клетканың бөлінуінде олар тұқым қуаламайды. Осындай жағдай болмас үшін векторлық молекулалар қолданылады. Век-торлар - ген тасығыштар, ал вектор деген сөздің өзі бағыттағыш деген мағынаны білдіреді. Сонымен бірге, векторларды рекомбинантты ДНҚ-ның міндетті бөлігі деп түсіну керек. Векторларды эксперименттік жолмен құ-растырады және оларға мынадай талаптар қойылады: 1) вектор клетка ішіне өзімен бірге бөтен генді алып кірген соң, репликациялана (көбейе) алатын болуы керек, сонда ғана ол келесі ұрпаққа тұқым қуалай алады, ол үшін векторға ДНҚ репликациясын бастайтын ерекше нуклеотидтер тізбегін енгізеді; 2) құрамында рестриктазалар танып, үзе алатын нуклеотидтер тізбегі болуы керек, әйтпесе векторға ДНҚ фрагментін енгізу мүмкін емес; 3) оның, бір немесе бірнеше таңбаланған гендері (маркерлері) болуы керек, сол таңбалар бойынша қажет геннің қайсы клетканың (реципиенттік) ішіне енгенін анықтайды; 4) вектордың, клетка ішіндегі копиялары (көшірмелері) жеткілікті мөлшерде болуы керек. Ген инженериясында векторлық молекуланың шығу козі болып бактериялық плазмидалар мен вирустар (әсіресе фагтар) саналады. Плазмида және фагтар негізінен бірінші талапқа сай келеді, ал, оларды келесі талаптарға сай келтіру үшін қосымша генетикалық өндеу жүмысын жүргізу керек.
Жоғарыда аталған векторларға қойылған талаптарға сәйкес кез келген генетикалық құрылым, басқа жағынан кез келген плазмида мен фагтар ген тасығыш бола алмайды. Ген инженериясында қолданылатын векторларды үш топқа бөлуге болады: 1) плазмидалар; 2) бактериофагтар; 3) космидтер және фазмидтер. Қазіргі кезде рекомбинантты ДНҚ технологиясында алғашқы екі векторлар типі жиі пайдаланылады, егер вектор ретінде плазмида қолданылса, онда ген клеткаға трансформация типімен енеді, ал бактериофаг (фаг лямбда) қолданылса, онда ген клеткаға трансдукция типімен енеді.
2. Плазмидалық векторлар. Вектор ретінде мөлшері 15- 20 мың н. ж. дейінгі, көбінесе 2-ден 10 мың н. ж. құралған кішігірім плазмидалар қолданылады. Плазмидалар бактериялардың хромосомадан тыс генетикалық элементі екендігін және олардың сипаттамасын біз қарастырған болатынбыз. Плазмидалық векторлар генді бактерияларға тасымалдап, оның тиімді жұмысын қамтамасыз ете алады.
Е. Соlі бактериясы үшін көптеген векторлық плазмидалар құрастырылды. Олардың ішінен әсіресе СоlЕ1 плазмидасының туындылары кең таралды. Ф. Болвар және Р. Родригес құрастырған осындай плазмида - рВR322 бірнеше мың жұп негіздерден (4362 н. ж. ) құралған. Бүл плазмидада антибиотиктер - ампициллин мен тетрациклинге төзімділіктің гендері бар және бірнеше рестриктазалар үзе алатын сайттары (нуклеотидтік нүктелері) бар рВR322 плазмиданың құрамында екі плазмида (рМВ1 және рSС101) және Тn З транспозоны бар. Ампициллинге тұрақтылықтың генінде Pst1 рестриктазасына арналған сайтқа және тетрациклинге тұрақтылық генінде ВаmН1 және SaL1 рестриктазаларына арналған қос сайтқа мән беріңіз. Осындай рестрикциялық сайттарда екі антибиотиктерге төзімділік гендерінің болуы қажет ДНҚ-сы (бөтен ДНҚ-сы) бар плазмидаларды сұрыптап алуға мүмкіндік береді.
Плазмиданы Ваm1 рестриктазасымен үзеді де, босаған орынға бөтен ДНҚ молекуласын (олар да, алдын ала Ваm1 рестриктазасымен үзіледі) жалғайды, нәтижесінде рекомбинантты плазмида тетрациклинді ортада өсе алмайды, өйткені плазмиданың tet генінің біртұтастығы бұзылған. Керісінше, плазмида ампицилинді ортада өсе алады, міне, осындай ортада көбейетін бактериялардан қажет генді оңай табуға болады. Бұл арада, вектор антибиотиктерге төзімділік гендері бойынша таңбаланды.
pBR322 векторынан басқа СоlЕ1 плазмидасы негізінде бірнеше векторлар құрастырылған. Басқа плазмидалар (рSC101 т. б. ) негізінде алынған векторлар да белгілі.
СolЕ1 плазмидасы негізінде құрастырылған вектордың кемшілігі де бар: бөтен ДНҚ-ның молекулалық массасы артқан сайын рекомбинанттардың саны (копиялары) азая түседі. Осы себептен ұзын ДНҚ фрагменттерінің (10 мың. н. ж. асатын) клондарын алу үшін фагтық векторларды, космид және фазмидтерді пайдаланады.
Фагтық векторлар. Вектор ретінде бактериялармен қатар вирустар да қолданылады. Олардан вектор алу әдістемесі лямбда (λ) фагында жете зерттелген. Лямбда фагтың ДНҚ-сы қос тізбекті және формасы түзу, оның геномы 48, 5 мың н. ж. тұрады. Фаг ДНҚ-сының 12 н. ж. құралған комплементарлы жабысқақ ұштары бар. Олар, фаг клеткаға енгеннен кейін бір-бірімен бірігіп, ДНҚ сақиналы формаға ауысады. Сақиналы ДНҚ репликациялық форма болып саналады. Лямбда фагтың ДНҚ молекуласында 60-тай ген бар. Фагтың геномында лизистік даму мен ұрпағын көбейту үшін қажет гендері (маңызды) жоқ екі учаскесі бар. Бірінші учаске фаг геномының орталық бөлігінде (маңызды емес гендер) орналасқан, оның ұзындығы 22, 0 мың н. ж. тең, екінші учаске P мен Q гендері арасында орналасқан, ұзындығы 3, 0 мың н. ж. тең. Міне
Сіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz