Әртүрлі бидай генотиптеріндегі құрғақшылықтан туындаған физиологиялық-биохимиялық өзгерістер

ӘЛ - ФАРАБИ АТЫНДАҒЫ ҚАЗАҚ ҰЛТТЫҚ УНИВЕРСИТЕТІ
БИОЛОГИЯ ФАКУЛЬТЕТІ
МАГИСТРАТУРА
Биотехнология, биохимия, өсімдіктер физологиясы кафедрасы
МАГИСТРЛІК ЖҰМЫСЫ
БИДАЙДЫҢ ӘРТҮРЛІ ГЕНОТИПТЕРІНДЕГІ ҚҰРҒАҚШЫЛЫҚТЫҢ ӘСЕРІНЕН ПАЙДА БОЛҒАН ФИЗИОЛОГИЯЛЫҚ - БИОХИМИЯЛЫҚ ӨЗГЕРІСТЕР
Орындаған Бериккайрова Г. М. «»2010 ж.
(қолы)
Ғылыми жетекшісі б. ғ. д., профессор Кенжебаева С. С.
(қолы)
«»2010 ж.
Қорғауға жіберілді:
Кафедра меңгерушісі,
б. ғ. д., профессор Карпенюк Т. А. «»2010 ж.
(қолы)
Алматы 2010
РЕФЕРАТ
Магистрлік жұмысы 39 беттен, 14 кесте, 7 суреттен тұрады, 31 әдебиет қолданылды.
Түйінді сөздер: Құрғақшылыққа төзімді өсімдік, пролин, сусыздану.
Зерттеу объектісі: Зерттеу объектісі ретінде астық тұқымдасы бидайдың жаздық 8 генотипі алынды. Астана, Актюбе 39, Байтерек, Волгоуральская, Ертис 97, Казахстанская 25, Надежда, Саратовская 29 сұрып таңдап алынды. Бұлардың сулы және құрғақ массалары алынды.
Зерттеу әдістері:
- Бидай тұқымының сулы, құрғақ массаларын және су мөлшерлерінің көрсеткіштерін алу.
- Өсімдіктердің құрамынан пролиннің мөлшерін анықтау.
Зерттеу жұмысының мақсаты: Бидайдың әртүрлі генотиптеріндегі құрғақшылықтың әсерінен пайда болған физиологиялық - биохимиялық өзгерістер.
Жұмыстың мақсатына жету үшін қойылған негізгі міндеттер:
- Физиологиялық көрсеткіштері бойынша: құрғақшылықтың әсерінен болатын сусыздану дәрежесіне талдау жасау.
- Биохимиялық көрсеткіштері бойынша: пролиннің осмолиттердің қасиеттерімен болатын дегидратация әсерінен бидай генотиптеріндегі мөлшерлерін анықтау.
Тақырыптың өзектілігі:
Құрғақшылық Қазақстанда бидай өнімділігінің жақсы дамуына аса қауіпті стресті фактор болып табылады, сондықтан бидайдың қуаңшылыққа төзімділігін арттыру үшін бірнеше сұрыптарын алып, олардың физиологиялық және биохимиялық қасиеттерін анықтайды.
Қазіргі кезде адаптациялық морфофизиологиялық қасиеті бар негізінен бидайды жаңа формаларын және генотиптерін қуаңшылыққа төзімділігін артып, сұрыптау көптеп жүргізіледі. Бірақ әртүрлі жаздық бидайдың генотиптерінде физиологиялық - биохимиялық қуаңшылыққа төзімділігіне байланысты физиологиялық және биохимиялықтың көрсеткіштерді дұрыс зерттелмеген. Құрғақшылық күрделі фактор болып табылады және ол үш компонентерден түрады: су тапшылығы (дегидратация стресс), күн радияциясының интенсивтілігі, жоғары ауа температурасы. Осы компонентердің ішінен су тапшылығын әсерін зерттеледі.
МАЗМҰНЫ
КІРІСПЕ
1 ӘДЕБИЕТКЕ ШОЛУ . . . 5
- Құрғақшылық ұғымы туралы . . . 5
- Өсімдіктердің бейімделу қасиеті . . . 6
- Өсімдікке ылғал жетіспеушілігімен жоғары температураның
әсері . . . 7
- Құрғақшылыққа АБҚ-ның бейімделу рөлі . . . 9
- Өсімдіктерден қуаңшылыққа төзімділігін анықтау және арттыру әдістері . . . 11
1. 6 Өсімдік клеткасы потенциялының су мөлшерінің төмендеуінен құтылу стратегиясы . . . 11
1. 6. 1 Осмолиттер. Қасиеттері мен функциялары. 12
1. 7 Дигидратация кезіндегі өсімдік клеткаларындағы пайда болатын ақуыздар . . . 13
1. 8 Қорғаныш және су тапшылығына индуцирленетін регуляторлы ақуыздар функциясы . . . 14
1. 9 Бидай және оның жалпы сипаттамасы15
- ЗЕРТТЕУ ОБЪЕКТІЛЕРІ ЖӘНЕ ӘДІСТЕРІ . . . 17
2. 1 Зерттеу объектілері17
2. 2 Зерттеу әдістер . . . 17
- ЗЕРТТЕУ НӘТИЖЕЛЕРІ ЖӘНЕ ОЛАРДЫ ТАЛДАУ . . . 22
3. 1 Тұқымға құрғақшылықтың әсері. 22
ҚОРЫТЫНДЫ . . . 36
ПАЙДАЛАНЫЛҒАН ӘДЕБИЕТТЕР ТІЗІМІ . . . 37
КІРІСПЕ
Соңғы он жыл ішінде біздің құрғақшылыққа төзімді өсімдіктермен болған зерттеу жүмыстарымыз едәуір қарқынды жүргізілуде. Себебі, жыл сайын болатын жауын-шашын мөлшерінің тым аз болуы, температураның көтерілуі, сусыздану көрсеткішінің жоғарылауына байланысты құрғақшылықтың туындауы себеп болып отыр және өсімдіктердің физологиялық және молекулалық тұрғыдан бейімделу дәрежесін өзгертеді. Осыған байланысты ауылшаруашылығында төзімді, түсімі мол өнімді алу кеңінен қолданылып отыр [1] .
Су тапшылығы (дегидратация стресс) . Өсімдіктің тамыр жүйесі суды сіңіру жылдамдығы транспирацияға қарағанда тез өтеді, соған байланысты өсімдіктер су тапшылығының зардабын шегеді. Бұндай жағдай тек қуаңшылықтың әсерінен емес, сонымен қатар топырақтың тұздылығының, төмен температураның әсеріне де байланысты болады. Бұл кезде дегидратация стресті “сусыз” табиғаттың компоненті өсімдікте жақсы жүреді, ал топырақта су мөлшері жеткілікті болса да дегидратация кезінде өсімдікке су тапшы болып жатады [2] .
Су тапшылығы кезінде клеткаларда су құрамы өте аз болады және су тапшылығының әсерінен цитоплазмадағы иондардың концентрациясының ұлғаюы нәтижесінде құрылысы мен биополимерлердің функциялары өзгереді. Сондықтан, әртүрлі генотипті 8 бидай сұрыпын алып, олардың дегидратацияға жауап реакциясын зерттейміз.
- Бірінші физиологиялық көрсеткіштерді бақылаудағы және әртүрлі дегидратация кезеңінін әсерінен өсімдіктердің жер бетіндегі бөліктерінде су мөлшерлерін анықтау.
- Екінші физиологиялық көрсеткіштерді құрғақ және сулы биомасса жиынтығы (жерасты мүшелері мен тамыр) анықтау. Ортадағы сорттың құрғақшылыққа төзімділігін анықтау.
- Биохимиялық көрсеткіштері - пролиннің осмолиттердің қасиеттерімен болатын дегидратация әсерінен бидай генотиптерінде мөлшерлерін анықтау.
1 ӘДЕБИЕТКЕ ШОЛУ
1. 1 Құрғақшылық ұғымы туралы жалпы сипаттама
Өткен 10 жылдың ішінде, біздің өсімдіктердің қуаңшылыққа қарсы әсері туралы түсінігіміз тез арада жоғарылады. Назар аударатын жағдай ол сигналдық процесс. Судың тапшылығы осы процестегі негізгі бөлім болып табылады. Ол өте қажетті яғни өсімдіктің қысымға деген төзімділігі, бұған өсімдіктің басқару және бағыну техникасы туралы жан-жақты түсінік керек [1] .
Құрғақшылық қазіргі кездегі ауылшаруашылық өндірістің маңызды мәселелерінің бірі, зерттеулер көрсеткендей, құрғақшылық пен шөлге айналған жерлер жыл сайын әлемдік масштабта ауылшаруашылығына 42 млрд доллар шығын әкелуде. Алдыңғы 1998, 2000, 2001, 2002, 2003 жылдарында ТМД барлық териториясындағы региондарда немесе басқа бөліктерінде құрғақшылық көп болған. Бұл жағдайдың қиын екендігін көрсетеді. Ауылшаруашылық культуралар құрғақшылықтан және басқада әсерлер салдарынан келген зардап тек бір Ресейде 2000 жылы 20 млрд рубль болған. Ауа - райын балжаушы мамандардың айтуынша әлемдік климаттың өзгеруі, сонымен қатар орташа температураның да ауытқуы кенеттен суып кетумен бірден ысып кетуге әкелуде.
ТМД елдері үшін құрғақшылық қалыпты жағдай болып табылады. ТМД территориясында ылғлдылыққа төзімсіз жерлерде бар, олар 250-500 мм ылғалдылықты тез қабылдап ұшып кетеді [1] .
Құғақшылыққа тұрақтылық - өсімдіктің ұзақ уақыттық құрғақшылыққа төзімділігі, суды аз сіңіруі, клеткалар, ұлпалар мен мүшелердің суды үнемдеп жұмсауы болып табылады.
Топырақтағы құрғақшылық жаңбырдың ұзақ уақыт жаумауы мен атмосфералық температураның жоғары болуымен және күннің инсоляциясымен, топырақтың беткі қабатының құнсыздануымен трансперациямен күшті жолмен пайда болады. Осының бәрі топырақтың тамырлы қабаттарына зиян келтіріп, ауа ылғалдығы төмендегенде су қорын азайтады [2] .
Атмосфералық құрғақшылық жоғары температурамен және төменгі ауа ылғалдығымен (10-20%) сипатталады. Қатты атмосфералық құрғақшылық құрғақ және ыстық ауа массасының ауысуымен пайда болады [2] .
Атмосфералық құрғақшылық, судың тез арада жұтылуына яғни транспирацияға әкеледі, бұл топырақтағы су көлемінің азаюына және өсімдіктердің солып қалуына әкеледі. Бірақта өсімдіктің тамыр жұйесі берік болса, құрғақшылық қатты әсер ете алмайды, онда температура өсімдік көтере алатындай болу керек. Ұзақ уақыттық атмосфералық құрғақшылық және жаңбырдың аз жаууы топырақтың құрғақшылығына әкеледі, ал ол өсімдіктер үшін өте қауіпті [2, 3] .
1. 2 Өсімдіктердің бейімделу қасиеті
Біз айтып отырған бейімделу қасиеті дегеніміз ол өсімдіктердің жоғары температураға, су тапшылығына, жоғары жарыққа бейімделу сияқты үш факторын қамтиды [3] .
Мақсаты тірі организімнің әртүрлі сыртқы ортаның стресс факторларына бейімделе алуы туралы жаңа теориялар мен ғылыми зерттеулерге түсінік беру. Бейімделу потенциалы деген және жалпы бейімделу қабілеті деген ұғым енгізді. Осы мәселе төңірегінде маңызды мәселелерге және оларды шешу жолдарына тоқталып кету керек. Өсімдіктердің тұрақтылық қасиетін қалай қалыптастыратынын, оның ішінде тұрақтандыру және қалпына келтіру қасиеттеріне тоқтала кету, тірі организімнің өмір сүруіне болатын артық заттарды иллюстрация жасай отырып мысал келтіреміз [3] .
Өсімдіктің су тапшылығына деген жауабы жайлы зерттеудің маңызы ерекше, жер-шарының көп қуаңшылық жерлері климаттың ауысуы жайлы көп кеңес сұрайды. Жоғары температурадағы концидент және радияция мен байланыстыра отырып, ең көптеген қоршаған орталық өсімдіктің және бір рет өнім беретін өімдердің тірі қалуына еріксіз көндіру проблемасын тудырады. Ауыл шаруашылық аймақтарының қолданатын су ресурсының мөлшері, қуаңшылықтың артуы және өсірудің жалпылануына байланысты. Келешекте су үлкен қамбаға айналса, тіпті ерекше жағыдайда өмір сүре алатын ауылшаруашылығының өзі қатал су тапшылық жағыдайын өзгерте алады. Егістіктегі көп жұмыстар өсімдіктің стратегиясынан бастап қуаңшылықтағы су статусының бақылауы физиологиялық және биохимиялық процесс. Негізгі өсімдік су тапшылығына деген жауабы жапырақтың құрамы және функциясымен тамыр морфологиясы мен кең қысым арасындағы байланыстың интерпретациясында жүреді. Өсімдіктің әртүрлі ортада бәсекелесіп өмір сүретін қабілеті жайлы түсінік ең негізгі мәселе болып отыр. Бірақ бұл молекула процесін қамтыуы және қысымға деген төзімдігі жайлы зерттеу. Қазіргі таңда, көптеген гендердің қуаңшылықта пайда болғаны әлде (анықталғаны), көптеген құралдар (ген бейнелеу үлгісі, трансгендік өсімдіктер тағыда басқа) олардың ерекше функциясын және ролінің өсімдіктің су тапшылығына әрі өнуін пайдаланады. Себебі, өсімдіктің қысымға деген жауабы өте күрделі, қандай да болмасын, осы гендердің көптеген функциялары әлі белгісіз. Керісінше, барлық қысымға жауапты гендер клетканың бейімделу процесіне қатысады. Кейбіреулері қысқа мерзімді көптеген зиянды жауапқа қатысады [4] .
Бейімделу потенциалы (БП) - бұл түсініктің астарында өсімдіктің өмір сүруге, өнуге, өздігінен дамуға және өздігінен регуллциялану қасиеті арқылы түсіндіріледі. Бұл өзгерістер сыртқы ортаның әртүрлі стресс факторы кезінде жүреді. Ауылшаруашылық әдебиеттерінде бұл термин жалпы бейімделу қажет і ( ЖБҚ ) ретінде қарастырылып жүр.
Д. М. Гродзинский еңбегінде бұл мәселе арнайы түрде атап көрсетілген. Организімнің өміріндегі, негізгінде оның бүкіл организімінде жүйелердің тұрақтылығына сөзсіз байланысты. Ал жұйелер: молекулалық, клеткалық, ұлпалық, организімдік, популляциялық, тұрақтылық әлсіз толқын арқылы бейнеленсе, өсімдіктің факторларының кез-келген түрінде өмір сүрумен сипатталады.
Беріктілік - бұл өсімдіктің көп жағдайда ішкі күйінің бейнеленуші шекке жетуі сияқты сипатталады. Өсімдік беріктілігі екі жағдайда қамтамасыз етіледі. Жалпы организм немесе клетка жүйесі өздігінен функцияларын атқаруы, бірінші әдісті тұрақтандырушы, ал екінші беріктілікті қамтамасыз етушы деп атайды [4, 5] .
1. 3 Өсімдікке ылғал жетіспеушілігі мен жоғары температураның әсері
Құрғақшылық біріншіден өсімдіктің су режиміне, содан кейін басқа да оның физиологиялық функцияларына әсер етеді.
Атмосфералық құрғақшылық пен жоғары температура немесе күн сәулесі қатар жүрсе өсімдіктің сабағы мен жапрақтарының өсуі тежеледі, өсім төмендейді. Ал кей кезде өсімдік “ыстық соққыдан” аз уақытта өлімге ұшырауы да мүмкін [5] .
Өсімдік ұлпаларына су жетіспеушілігі оның транспирация кезінде көп кетуімен байланысты. Бұл ыстық күнді ауа райында күннің ортасында байқалады. Осы кезде жапырақтағы су көлемі таңғы көлеммен салыстырғанда 25-28% төмендейді, ал өсімдік тургорлығын жоғарлатып, сола бастайды. Нәтижеде жапырақтың су потенциялыда төмендейді, бұл топырақтан өсімдікке қажетті су көлемін арттырады.
Өсімдіктің солыуының екі түрі бар уақытша және жоғары уақытша солу негізінде атмосфералық құрғақшылыққа байланысты. Онда топырақтағы су өсімдікке уақытында жетпегендіктен өсімдік сола бастайды. Уақытша солу кезінде жапырақ тургоры кешкі және түнгі уақытта қалпына келеді, уақытша солу өсімдіктің өнімділігін төментетеді, себебі саңылау тургоры жоғалғандықтан саңылау жабылады және фотосинтез бірден тоқтап қалады [6] .
Тереңдік солу, топырақтың тамырдан суды мүлдем ала алмайтын кезінде болады. Құрғақшылық осы кезде болса, онда ол өсімдіктің өлуіне әкеп соғады. Солу кезінде өсімдік әртүрлі көлемде су жоғалтады. Қараңғы сүйгіштер 3-5%, ал тұрақтылары 20-30% [7] . Кейбір мәліметтер бойынша суықпен өңдегенде микроспоралардың плазматикалық мембранасының жағдайы өзгереді. Суықпен өңдеу режимі өсімдік түрлеріне байланысты болады: арпа - 28 күн, 4℃ - та, 14 күн 7℃ - та; бидай - 5-7 күн 3℃ - та, 8 күн 4 - 5 ℃- та.
Арпа мен бидайдың тозаңқаптары және бөліп алынған микроспораларының культурасы үшін 3 орта қолданылады: N
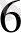 , Rotato 11, MС - модификация. Ұлпалар культурасында қолданылатын қоректік қоспалардың минералды компопненттерінің негізін MС орта құрайды. N
, Rotato 11, MС - модификация. Ұлпалар культурасында қолданылатын қоректік қоспалардың минералды компопненттерінің негізін MС орта құрайды. N
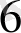 және Rotato 11 орталары тек дәнді дақылды өсімдіктердің тозаңқаптарын культивирлеу үшін жасалған.
және Rotato 11 орталары тек дәнді дақылды өсімдіктердің тозаңқаптарын культивирлеу үшін жасалған.
Күріш тозаңқаптары культурасында ондрогенезге нитраттың жоғары концентрациясы мен аммонийдың аз мөлшері ықпал етеді. Қоректік ортадағы Fe
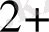 иондары микроспоралардың in vitro дамуындағы эмбриогенезді бақылайды.
иондары микроспоралардың in vitro дамуындағы эмбриогенезді бақылайды.
Көмірсуды қажет ету деңгейіне қарай өсімдіктерді шартты түрге 2 топқа бөледі: көмірсуды аз қажет ететін (2 - 4%), жоғары концентрацияда қажет ететін (6 - 12%) .
Көптеген зерттеушілердің айтуынша, картоп экстрактін, әртүрлі амин қышқылдарын ортаға қосқанда кейбір өсімдіктердің тозаңқап культурасына оң әсер етеді. Картоп экстрактінің құрамы анықталмаса да, дәнді дақылдардың тозаңқаптарын культивирлеуде кең қолданылады. Табиғи крахмал тек қоректік компмнент емес, сонымен қатар гель түзуші тасмалдағыш ретінде де қолданылады. Арпа микроспораларының культурасында эмбриоидтардың индукция жиілігі ионозитті 1000 - 5000 мг/л концентрацияда глутаминмен ( 160 -180 мг/л) бірге пайдаланғанда артты. Бидай тозаңқаптары культурасында ортаның кейбір қосымша компоненттерінің эффективтігі глутамин + зеатин + картоп экстракті қосылғанда байқалды.
Су тапшылығы мен солу өсімдіктің физиологиялық процестерінде әсер етеді. Клеткада бос көлемі төмендейді, концентрация жоғарлайды және вакуулдер шырынының pH төмендейді, ал цитоплазма ферменттердің тез шыға бастайды.
Ұзақ мерзімді солу кезінде ферменттердің активтілігі төмендейді, олар синтез процестерін қамтамасыздайды және гидролитикалық процестерді қамтамасыздайтын ферменттер көтеріледі, сонымен қатар протеолиз немесе аммиак, полисахарид, тағыда басқа биополимерлер протеолидті жүреді. Көптеген өсімдіктер жинақталып бұзылады. Егер су тапшылығының көлемі артса, нуклеинді алмасу, синтез және DNK синтезінің жүруі тоқтамайды. Жапырақтарда синтез төмендейді. Ыдырауы жүре DNK полимеразаның суббірліктерінің ыдырауы жүреді. Митоз тоқтамайды, белок синтезі тоқтағандықтан өсімдік өлімге ұшырайды [6, 7] .
Соңғы кездерде өсімдіктердің қолайсыз жағдайларына, соның ішінде қуаңшылыққа төзімділігі белоктардың биологиялық ырықтығы мен құрамындағы SH және S-S топтардың мөлшеріне де байланыстылығы туралы пікірлер қалыптасты. Топырақ құргақшылығының әсерінен тамыр жүйесінде фосфорлы қосылыстардың алмасуы бұзылып, АТФ, нуклеин қышқылдар, белоктар мөлшері азайып, қанттардың мөлшері 7-10 есе көбейеді. Ұзаққа созылған қуаңшылық ауылшаруашылық дақылдарының өнімділігін төмендетеді [8] .
1. 4 Құрғақшылыққа АБҚ - ның бейімделу рөлі
Алдыңғы жұмыстарда АБҚ-ның синтезі изопреноидтық метаболизмнің C 5 буының конденсациясынан кейінгі бір тармағы ретінде жүретіні болжанды.
АБҚ - нің тікелей синтезі қарапайым алғы заттардан түзілеті болжанды, бірақ АБҚ - нің тікелей синтезі тек фитопотогендік саңырау құлақтарда болатыны көрсетілді. Мысалы: (сұр шіріктің патогенінде - Bovytis)
АБҚ - нің тікелей синтезі 1990 жылдары негізгі ғылыми болжам болды. Бірақ сонан кейін өсімдіктерде ерекше мутацияларды каратиноидтың биосинтезі бұзылуы ашылған. Осы мутациялармен бірдей АБҚ - нің биосинтезі бұзылды. Осы ашылымнан шыққан қортынды өсімдіктерде АБҚ күрделі алғы заттар синтезделеді [9] .
АБҚ - нің ақуыз рецепторлары дұрстап зерттелген жоқ. Бір ақуыз 1 RPK (receptor - like proteinkinase) деп аталатын рецептордың рөлін атқарады. Ол протеин киназдық цитоплазмалық дәрмені бар таныс мембраналық ақуыз АБҚ мен байланысады.
АБҚ - дабылдың трансдукция жүйесінің кейбір компаненттері белгілі. Мысалы: АБҚ - нің әсері басталғанда, саңылаулардың тұйықтаушы клеткаларында НАД + - тан циклдік АДФ - рибоза синтезі басталады. Осы месенжерлер тонопластың Са 2+ - дің йондары вакоулдері мен басқа ішкі клеткалық деподан цитоплазмаға шығады. Цитозолдың рН ұлғаяды. Барлық аталған реакциялар протеин-фосфотаздарды 1ABI және 2ABI активтенеді. Олардың рөлі АБҚ дабылдың ақуыз рецепторлары, дефосфорлау репрессор диферилдеу әсерімен инактивилдейді және содан клетка АБҚ - ға жауап береді. АБҚ репрессор әрқашан жауап реакцияны тежейді және өсімдік осы гармонның әсеріне сезімтал болады, АBA (insensitive) саңылаулары жабылуы АБҚ - нің әсерінен өте жылдам жауап реакциясы болады. Сондықтан геном деңгейінде олардың қосу мүмкіндігі болмайды [9, 10] .
Ядродағы гендердің процессорлары ыдырату 3ABI реттеуші ақуыздың маңызы көп есептеледі.
АБҚ - осмотикалық стресс әсерінен гормоны АБҚ клеткада судың мөлшері өзгергенде пайда болады. Қалыпты жағдайда судың мөлшері өзгеруі үшін факторға тәуелді болып келеді.
- Кебу
- Клеткада қосындылардың концентрациясының ұлғаюы
- Суықтың әсері
Барлық аталған жағдайда су клеткаға жетпейді, яғни судың жетіспеуі байқалады. Қалыпты тіршілікте клетканың барлық молекулалары сумен байланысады. Мысалы: ақуыздармен нуклеин қышқылдар сутегі байланыстарымен суды ұстайды, егер макромолекулалар сумен байланысқан күйінде болса, су қыйншылықпен буланады, сонымен қатар клеткада белгілі йондық күш қолдайды. Ол белоктардың және нуклеин қышқылдардың конформациясын қолдау керек [11] .
Судың тапщылығымен йондардың концентрациясы ұлғаю нәтижесінде биологиялық активті молекулалардың денетурациялану мүмкіндігі болады, сондықтан клетка деңгейіндегі АБҚ - нің әсерінен осмотикалық белсенді қосындылардың концентрациясы, яғни оксиполин, амино қышқылдың, сахарозаның және тағыда басқа заттар ұлғаяды. Төмен молекулалық қосындылардың концентрациясы ұлғаяды, осмотикалық күші ұлғаяды, ал ол судың жоғалуына кедергі жасайды.
АБҚ - ның әсерінен 10 - 15 минут ішінде әртүрлі өсімдіктерде саңылаулары жабылады. Егер судың тапшылығы тамырда болса, онда тамыр АБҚ арқылы жапырақтарға дабылды тапсырады, саңылаулары жабылады. өте күшті қуаңшылықта өсімдік төменгі жапырақтарды тастайды, содан булануы тез арада төмендейді.
АБҚ ауксин әсерінен клеткалардың созылуын тежейді және ол ауксин тасымадануын тоқтатады. Сонымен өсімдіктің қалыпты физиологиясы бұзылыды [11] .
Төмен температура әсерінен су кристалл күйіне аусады, соның нәтижесінде мембраналардың құрылымы бұзылады. Сондықтан, суықтың әсері клетканың ең манызды міндетті - судың кристалл күйіне ауысуын тоқтату, яғни суы бар қосындылармен байланасу керек.
- Клеткада суықтың әсерінен судың тапшылығы сияқты бірдей жауап реакциялар байқалады. Белоктардың, ДНК - ның, РНК-ның синтезі тоқтайды. Клеткада оксипролин, қанттар, полиаминдер мен осмотин жиналады. Суықталған клеткада қанттар мен осмотин мұздың қалыптасуына кедергі жасайды.
- Бояушы қосындылардың, яғни антоциандердің биосинтезі ұлғаяды. Өсімдіктерде қызыл бояу пайда болады. Бірақ, осы физиологиялық жауап реакциялардың манызы әлі түзінксіз болады [12] . Өсімдіктердің қуаңшылыққа төзімділігін анықтау және арттыру әдістері
Өсімдіктердің қуаңшылыққа төзімділігін алдын-ала болжаудың немесе анықтаудың тікелей және жанама әдістері бар.
Табиғи жағдайда атмосфера мен топырақ қуаңшылығы жыл сайын бола бермейтін құбылыс. Сондықтан қуаңшылықты қолдан жасау мақсатында құрғатқыш шатырлар қолданылады. Оның айналасынан тереңдігі 80 см, ені 50-60 см ор қазылып, төбесі су өткізбейтін материалмен жауын-шашын болардан бұрын жабылады да, басқа уақытта ашық қалдырылады. Осындай жан-жағынан ылғал түспейтін шатырдың ішінде зерттелетін өсімдіктер егіп, тиісті бақылаулар жүргізіп, түсімі есептелініп, қалыпты жағдайда өсірілген өсімдіктермен салыстырылады. Алынған салыстырмалы нәтижелер арқылы зерттелген өсімдік сорттарының қуаңшылыққа төзімділігі туралы тиісті қорытынды жасалады.
Осы әдіске жақын солдыру әдісінде зерттелетін өсімдіктер топырақ салған ыдыста өсіріліп, әлсін-әлсін жасанды қуаңшылық жағдай туғызылады.
Қысқа мерзімді тәжірибелерде өсімдіктерді суда өсіру әдісі қолданылады. Ол үшін өсімдік өз алдына бір ыдыста өсіріліп, ал ыдыс ішінде қоректік орта бар екінші ыдысқа орнатылады. Керек кезінде өсімдік өсіп тұрған ішкі ыдыс белгілі мерзімде суарылмайтын жағдайда ұсталады.
Соңғы кездерде өсімдіктердің қуаңшылыққа және ыстыққа төзімділігі тікелей талдау-протоплазманың тұрақтылығы, синтездік қабілеттілігі, гидрофилділігі, белоктардың коагуляциялануы, тағыда басқа көрсеткіштер арқылы анықтау әдістері енгізілді.
Өсімдіктердің қуаңшылыққа төзімділігін арттыру мақсатында шынықтыру, әртүрлі тұздар ерітіндісімен өне бастаған тұқымды өндеу әдістері қолданылады [12, 13] .
1. 6 Өсімдік клеткасы потенциялының су мөлшерінің төмендеуінен құтылу стратегиясы
... жалғасыСіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz