Бидай алейрон қабатындағы плазматикалық мембрананың Н+-АТФ-азасының активтілігі және цитокинин медиаторының әсері

ӘЛ-ФАРАБИ АТЫНДАҒЫ ҚАЗАҚ ҰЛТТЫҚ УНИВЕРСИТЕТІ
Биология факультеті
Биотехнология, биохимия, өсімдіктер физиологиясы кафедрасы
БІТІРУ ЖҰМЫСЫ
Бидайдың плазматикалық мембранасындағы Н + АТФ-азаның активтілігін анықтау
Орындаған:
4 курс студенті Спанкулова З. Б
Ғылыми жетекшілері:
ҚР ҰҒА-ның
академигі Гильманов М. Қ
“__” 2008ж
х. ғ. д., профессор Шойынбекова С. Ә
“__” 2008ж
Норма бақылаушы: Акимбекова А. Б.
“__” 2008ж
Қорғауға жіберілді:
Кафедра меңгерушісі,
б. ғ. д., профессор Иващенко А. Т.
“__” 2008ж
Алматы, 2008
РЕФЕРАТ
Бітіру жұмысы: 29 беттен, 35 әдебиеттен, 4 кесте, 3 суреттен тұрады.
Бітіру жұмысын Спанкулова Зере Бақтыбекқызы М. А. Айтхожин атындағы биохимия және молекулалық биология институтының ферменттердің құрылымы мен реттелу лабораториясында өткізді.
Өзектілігі: Мембранамен байланысқан АТФ тәуелді ферменттердің биохимиялық қасиеттерін анықтап, оларға цитокинин медиаторының әсерін зерттеу ферменттердің белсенділігін арттырып, ауылшаруашылығы дақылдарының өнімділігін жоғарылатады.
Негізгі сөздер: бидай дәні, ұрық бөлігі, алейрон қабаты, Н + АТФ-аза НАДФ глютаматдегидрогеназа, цитокинин, цитокинин медиаторы, фузикокцин
Алдына қойылған мақсаты: Бидайдың плазматикалық мембранасындағы Н + АТФ-азаның биохимиялық қасиеттерін анықтау және оған цитокинин медиаторының әсері мен оның физиологиялық, биохимиялық қасиеттерін сипаттау және биотехнологияда қолдану жолдарын қарастыру.
Қолданылған әдістер: Октил сефароза (CL-4B) гидрофобты хроматографиясы және RP-18 колонкада жасалынатын жоғары қысымды кері фазалы хроматография, гомогенизациялау, центрифугалау.
Қысқартылған сөздер
НАДФ-ГДГ- глютаматдегидрогеназа
ЦББ- цитокинин байланыстырушы белоктар
АТФ- аденазин үш фосфат
АДФ- аденазин екі фосфат
ГДГ- глютаматдегидрогеназа
ФК- фузикокцин
6-БАП- бензоламинопурин
МАЗМҰНЫ
КІРІСПЕ
М. А. Айтхожин атындағы молекулалық биология және биохимия институтының ферменттердің құрылымы мен реттелуі лабораториясында ең алғаш рет фузикокцинге ұқсас екінші реттік цитокинин гормоны ашылды. Ол клеткадағы сигналды трансдукцияда маңызды роль атқаратындығы анықталды. Медиатор екінші сатыдағы сигналды беретін зат, себебі гормон тікелей өзі емес медиаторлар арқылы клеткаға әсер ете алады. Сол себептен цитокинин медиаторын зерттеу ғалымдардың қызығушылығын тудырады. Себебі осы цитокинин медиаторы клетка ішіндегі мембранамен байланысқан ферменттерге әсері өте жоғары [1] .
Бұл медиатор бидай тұқымының өніп жатқан ұрығында цитокинин әсерінен пайда болады. Фк мен цитокинин медиаторы арасында жақын қатынасты дәлелденген қосыша мәліметтер алуға болады. Бұл жоспарда ФК-нің өсімдік клеткасының плазматикалық мембранасының Н + АТФ- азасының активтілігіне қасиеті бар екені белгілі. Осы себепті бидай өскіндері тамырының плазматикалық мембранасының Н + АТФ- азасына цитокинин медиаторының әсерін зерттеу мүмкіндігі туындайды [2] . Екінші реттік цитокинин гормоны бидай дәніндегі ұрықтық емес бөлігінің плазматикалық мембранасындағы Н + АТФ-азасын активтендірмейтіндігі белгілі болды. Осыған байланысты осы екінші реттік цитокинин гормонының бидай дәнінің алейрон қабатындағы плазматикалық мембрананың Н + АТФ-азасына әсерін зерттеу қажет болды.
Осы себепті зерттеудің мақсаты: бидай дәнінің алейрон қабатындағы плазматикалық мембрананың Н + АТФ-азасының активтілігін жоғарылату.
Осы мақсатқа жету үшін алға қойылған міндеттер:
- Цитокинин медиаторын бөліп алу;
- Бөліп алынған цитокинин медиаторының бидай дәнінің алейрон қабатындағы плазматикалық мембрананың Н+АТФ-азасының активтілігіне әсерін қарастыру;
1. ӘДЕБИЕТКЕ ШОЛУ
Гормондар- тірі организмдердің тіршілігі үшін маңызы өте зор. Организмдердің дамуы, өсуіндегі ең негізгі процестер гормондармен реттелуге тәуелді. Гормондар жетіспеген жағдайда немесе артып кететін болса онда организмді ауруға ұшыратады [3, 4] .
Яғни осы қызметтеріне қарап гормондарды-организмді реттеуші патшалық деп айтуға болады. Өсімдік тіршілігін реттеуде өсімдік гормоны цитокининнің маңызы зор [5] . Цитокининнің функциясы өсімдік онтогенезінің барлық кезеңдерінен көрінеді. Цитокининді зерттеу аймағындағы жаңа жетістіктер мен көріністерге кейінгі жылдары ғалымдардың қолы жетуде.
Цитокинин- клетка бөлінуін реттейді, хлоропластардың дифференцациясына қатысады. Клетка бөлінуі үшін цитокининнің маңызы зор. Осы гормонның сигналды трансдукция механизмі әлі анықталмаған. Кейінгі зерттеулердің барлығы дерлік өсімдік фитогормондарының өсу процесіне, клетка бөлінуіне әсерін зерттеу бағытында жүргізіледі. Фитогормондар клеткамен клетка, ткань мен ткань, мүше мен мүше байланысқанда көптеген морфогенетикалық және физиологиялық программаларды атқарады [6] .
Фитогормондар өсімдік организміндегі заттардың мөлшеріне, маңыздылығына, көбею ерекшелігіне, клетканың бөлінуіне, тыныс алуға, қартаюға, зат алмасуға қатысады. Өсімдік тіршілігін реттеуде өсімдік гормоны цитокининнің маңызы зор [7] .
Цитокининнің функциясы өсімдік онтогенезінің барлық кезеңдерінен көрінеді. Цитокининді зерттеу аймағындағы жаңа жетістіктер мен көріністерге кейінгі жылдары ғалымдардың қолы жетуде.
1. 1. Фузикокциндердің ашылуы мен химиялық құрылысы
Фузикокцинді (ФК) 1964 жылы италияндық ғалым Alessandro Ballio Fusicoccum amygdale фитопатогенді саңырауқұлағының фитоитоксині ретінде ашты. Бұл фитоксин жас миндаль ағаштарының жапырақтарының саңылауларын ашқан да, осыдан соң олар жабылмаған [7, 8] . Артық транспирация және тамыр жүйесінің әлсіздігінің нәтижесінде жас ағаштар тез кеуіп кетеді, яғни фузикокцин табиғи дексикант болып табылады. Профессор A. Ballio лабораториясында осы фитотоксиннің құрылысы анықталынды (1-суретте көрсетілген )
Фузикокциннің гибберлиндер сияқты табиғи терпеноидтарға жататындығын Gronevald авторларымен бірге ескере отырып, гибберлиндер мен фузикокциннің туыстығы туралы гипотезаны шығарды. Бұл екеуі де ауру саңырауқұлақтардың дитерпеноидтары, метаболиттері болып табылады. Уақыт өткен сайын гиббереллиндердің саны артуда. 1960-1980 жылдардын бері сол сияқты фузикокциндердің де 15-ке жуық топтары белгілі болды. Олар А, В, С символдарымен ажыратылады [9] .
Жоғары сатылы өсімдіктерде фузикокцин тұқымдасына жататын заттардың бар болуы туралы алғашқы мәліметтерді 1980 жылы Муромцев қызметкерлерімен бірге тұңғыш рет баспадан басып шығарды. Кейін фузикоцинді табу үшін жоғары қысымды сұйық хроматографта материалды алдын ала фракционерлейтін қолданды. Бастапқыда авторлар Fusicoccum amygdale саңырауқұлағының културальды сұйықтығында фузикокцинді метаболиттерді анықтаудың тәсілдерін ойлап тапты, ол газды хроматография мен трисилил туындыларының масспектрометриялы аралас әдісі, мұнда детектирлеу бірнеше таңдалған сипатталған иондармен жүзеге асады [10] .
Авторлар фузикокцинді алу үшін жүгері собығын және орамжапырақ жапырақтарын қолданды. Оларды этил спиртінде гомогенизирледі. Спиртті экстрактыларды концентрлеп, концентратты хлороформмен экстрагирледі. Хлороформды экстрактыны сұйық хроматографиядан өткізді. Алынған пикті ізінше масспектрометриямен жүретін газ сияқты хроматографияға сынады.
Өсімдіктегі экзогенді А фузикокциннің құрамы 10 -10 -10 -12 М құрады, бұл эндогенді гибберлиндердің құрамынан 2-3 есе төмен. Бұл жерден жоғары сатылы өсімдіктерден табиғи фузикокциндердің препаратты мөлшерін бөліп алу мәселесі күрделі екендігі анық болып отыр [11] .
Кейінірек жоғары сатылы өсімдіктерден табиғи фузикокцинді бөліп алудың тағы да бір мүмкіндігін осы авторлар қолға алды. Бұл жерде авторлар фузикокцинді бөліп алудың обьектісі ретінде қарқынды өсуімен ерекшеленетін трансформацияланған қына тамырларының культураларын пайдаланды. Бұл обьектіні таңдап алу себебі төмендегідей, біріншіден фузикокциннің көзі болып табылатын микроорганизмдер мен саңырауқұлақтардың зақымдану мүмкіндігі толығымен болмады. Екіншіден, олардың қарқынды өсуіне байланысты тамыр клеткаларының культурасын көп мөлшерде алуға болады [12] .
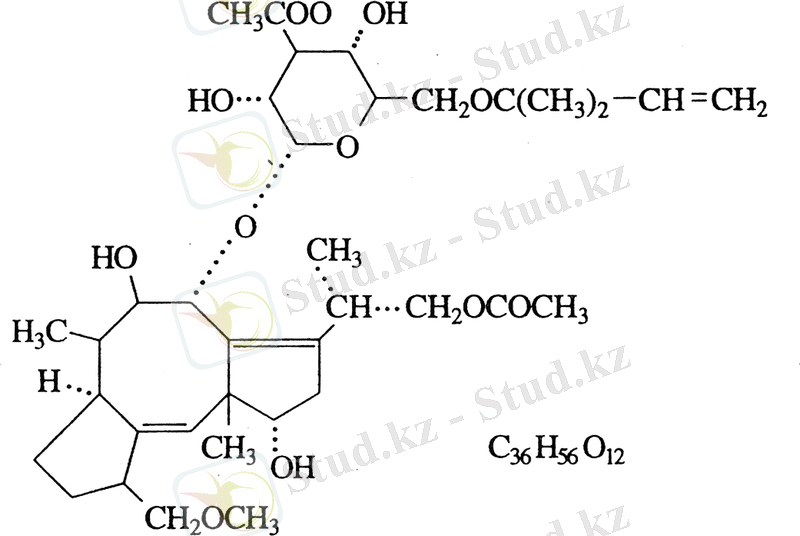
1 сурет Fusicoccum amygdali Del. Фитопатогенді саңырауқұлағының фузикокцин - фитотоксин формуласы (1964 жылы Ballio et all сипаттаған)
Тамыр клеткаларының культураларынан авторлар спиртті экстрактыларды алды. Экстрактыларды вакуумның астында буландырып, су қалдықтарын хлороформмен экстрагирледі, одан соң сулы және хлороформды фазаларда фузикокциннің болуын анықтауды иммунды тәсілмен жүргізді. Фузикокцин хлороформда болды, ал сулы фазада тек іздің мөлшері ғана болды, ал бұл белок және қант қосылыстары сияқты рецепторлар мен антиденелердің тиімді байланысуын тежейді. Фузикокцин сияқты заттарды тамырдың барлық культивирлену кезеңінде тауып отырды (14 тәуліктен бастап) . Тазартылған тамырда өсірілген фузикокциннің максимальды құрамы 1кг тамырға 150н моль-ге дейін құрайтынтындығы анықталды [11, 13] .
Алынған фракциялардың масспектрометрлі анализі мынаны көрсетті: негізінен А типті фузикокцин болды, сондай-ақ берілген анализдер басқа анықталынбаған фузикокциннің болу мүмкіндігін көрсетті. Зерттеулер нәтижесінде эндогенді фузикокцинді идентификациялау қиын деген қорытындыға Муромцев қызметкерлерімен тоқталды.
Алғашқы және негізгі идентифицирленген фузикокциндердің бірі А фузикокцині болып табылады. Ол өз алдына гликозидті үш корбооксилді дитерпендер, молекулалық салмағы 680 кД және формуласы С 36 Н 56 О 12
ФК молекуласының агликонды бөлігі үш циклді жүйе болып табылады. Бұл жүйе сегіз бұрышты және бес бұрышты сақинаны біріктіреді, сонымен қатар ФК -нің молекуласының агликонды бөлігі тотығу кезінде ацетилді топпен байланысады. ФК-нің 10 шақты түрі моно, ди, үш ацетаттар. ФК басқа қатары өзгеше құрылысты болып келеді. Олардың 20-ыншы көміртегі атомы тотықпаған. Әдебиеттегі мәліметтер бойынша фузикокциннің молекуласы табиғатта кең таралған дициклопентан және циклооккандардың байланысының туындысы болуы мүмкін деген тұжырымдар бар [14] . Мұндай құбылыстар саңырауқұлақтарда, балдырларда, кейбір жоғары сатыдағы өсімдіктерде, тіпті жануарларда (жәндіктерде) байқалған. Фузикокциндердің көптеген түрлері әртүрлі организмдерден бөлініп алынған, мысалы: фузикоплагин С, анаденсин, эпоксидилтимен, т. б. Циклопентанды, циклооканды байланыстары бар терпеноидтар номенклатурасы бойынша фузикоккандар қатарына кіреді. Мұндай фузикоккандардың байланысы транс-син-транс С 20 -көмірсутек ретінде белгілі болған [14] .
Фузикокцинді ары қарай зерттеу олардың физиологиялық, биохимиялық қасиеттерінің алуан түрлі екенін көрсетеді. Бұл жағдай көптеген өсімдіктердің өсуінде табиғи реттеуші екендігіне мүмкіндік береді. Көптеген жоғары сатыдағы өсімдіктердің клеткалары, мүшелері, ұлпалары ФК көлемінің ұлғаюына жауап беруге байланысы бар, сол сияқты саңылаулардың ашылуы, ұрықтың өсуінің қарқыны, көбеюі, т. б. процестерге байланысы бар. ФК және ФК заттар тұқымның өсуіндегі маңызды эндогенді реттегіш болып табылады деген мәліметтер бар. Г. С. Муромцев лабораториясында ең бірінші рет фузикокциндердің тұқымның ризогенезіне активті әсер ететіні анықталған. Фузикокцин асбұршақ тұқымының өсуіне өте қызықты әсер көрсетті. Осылайша ол котилидон клеткаларының өнуін стимулдап отырды да, бір жағынан осьтің ұрықтарының өсуін тежеп отырды, бұл фузикокциннің полярлы әсері бар екендігін көрсетеді [11, 14] . Микроорганизммен зақымдалуына маңызды роль атқарады, яғни фузикокцин бұршақ тұқымдастарда түйнектердің түзілуін активтейтіні анықталды.
Фузикокцин жапырақтың бағаналы клеткаларымен көмірқышқыл газын меңгеруін активтейді. Сондай-ақ фузикокцин Vigna angularis калеоптилінің клеткалар апопластарындағы аскорбат концентрациясын реттейді.
Фузикокциндердің гормоналдық қасиеттерінің бірі-оның антистресстік активтілігі. Г. С. Муромцев ФК-нің әртүрлі орта жағдайларында тұқымға әсер етуін дәлелдеген. Өсімдіктер үшін ФК еш күмәнсіз маңызды да, күшті антистресстік байланыс болып табылады . Муромцев қызметкерлерімен бірге өсуге қолайсыз жағдайда тұқымдардың ұқсастығын фузикокцин арттыруға қабілетті екенін көрсетті, мысалы: жоғары және төмен температураларда, артық ылғалдануда, тұздануда және т. б. РАН өсімдіктер физиологиясы институтының қызметкерлері тұқымдарды малу (0, 68 мг/л фузикокцин), қыстық бидай, арпа, сұлыға шашырату (0, 34 мг/л фузикокцин) арқылы өсімдіктердің аязға төзімділігін арттыратындығын көрсетті. Аязға төзімділіктің артуы фотосинтетикалық аппараттың даму деңгейімен қанттың жиналуымен жақсы коррелирленеді, сонымен қатар, клеткада эндоплазматикалық ретикулумның дамуын жылдамдатумен де ерекшеленеді. Фузикокцин күрішті тұзданудан қорғайтындығы, әртүрлі ауруларға картоп түйнектерінің тұрақтылығын арттыратындығы көрсетілді. Фузикокцин өсімдік үшін ең күшті әсер ететін антистресті қосылыстардың бірі болып есептелінеді [14, 15] . Фузикокциннің осмотикалық стреске бейімделу кезіндегі жағымды ролі анықталды.
Біз үшін фузикокциннің биохимиялық активтілігін зерттеу бойынша деректер қажет. Тұңғыш рет 1977 жылы Alessandro Ballio қызметкерлерімен бірге жүгері өскіндерінің ұштарынан бөліп алынған плазматикалық мембранасынан фузикокцинді рецепторларды тапты. De Boer қызметкерлерімен бірге Фк байланыстыратын белоктарды тазалауда арнайы аффинді сорбентте тазалау әдісін ойлап тапты, бұл жерде активті топ ретінде биотинилирленген фузикокцин тігілген [16] .
Осы авторлар фузикокцин байланыстырушы белоктарды бөліп алуда жұмыстар жүргізді. Richard G. Stout және Robert E. Cleland фузикокцинді рецепторды зерттеу үшін фузикокцинге қарсы моноклональды антиденелерді алды.
Фузикокцин үшін мүмкін рецепторлар берілген, олар фузикокцин байланыстырушы белоктартың (ФКББ) екі типі. Бірінші типінің константты диссоциациясы (Kg) шамамен 10 -11 М (жоғары аффинді ВА) болатын фузикокцинге жоғары ұқсастығы бар, ал екінші типінің константты диссоциациясы (Kg) шамамен 10 -7 М (төмен аффинді НА) болатын төмен ұқсастығы бар рецепторлар. Тұңғыш рет осы рецепторлар жүгері калеоптилінің плазматикалық мембранасы бар функциялардан табылды. Жұмыста белгіленген мембраналарда рецептордың жоғары аффинді сайттары мөлшерінің төмен аффинділірге қатынасы - [ВА] / [НА] шамамен 1-ң 2-ге қатынасын құрайтындығы көрсетілген. Авторлар фузикокцин әсеріннен тек жоғары афиндік сайттың байланысуы қатысады деген тұжырымға келді. Мұндай көзқарастар кейінгі жылдары басымдылық таныта бастады. Фузикокцин рецепторы ретінде жоғары аффинді сайттың қатысуы фузикокцин байланыстырушы белоктан және Н + АТФ-азадан құралатын in vitro жүйесінің реконструкциясы бойынша жұмыс істеген авторлармен болжамдалады [17] .
Көптеген оқшауланған мембраналарда төмен аффинді сайттардың аз ғана мөлшері немесе жоқтығы бөліну процесінде олардың инактивациясымен байланысты болуы мүмкін. Бұл жоғары аффинді сайтпен салыстырғанда төмен аффинді сайттар байланысының үлкен лабильділігімен түсіндіріледі. Қазіргі уақытта жоғары және төмен аффинді сайттардың қызметтік ролі туралы сұрақтар зерттеу обьектісі болып отыр.
Жоғары аффинді фузикокцин байланыстырушы белоктарды зерттеуде жақсы нәтижелерге қол жеткізді, олар бөлініп алынып сипатталынды. Бір жағынан, бұл белоктар гетеродимерлер болып табылатындығы көрсетілді (әрбір мономердің молекулалық массасы 30 кДа жақын) Бұл полипептидтердің сиквенс бойынша мәліметтері сұлының тамырының, Commelina communis жапырақтарының және жүгерінің калеоптилінің ФКСБ құрылымы бойынша белоктардың 14-3-3 класына жататын эукариоттардың көпфункциональды реттегіш белоктарымен гомологты болып келетіндігі көрсетілді. Фузикокцин байланыстырушы белоктар 14-3-3 белоктар тұқымдасына жататындығы туралы De Boer-дің жұмысында көрсетілген. 14-3-3 белоктар редуктаза нитратын ингибирлейтіндігі көрсетілді, алайда бұл белоктардың фузикокцинмен байланысуынан кейін редуктаза нитратының активациясы байқалды.
ФК Н + АТФ-азаның бірден бір активаторы болып саналады. Ол 14-3-3 белок регуляторымен байланысқан. Сондықтан да Н + АТФ-аза фузикокциндердің үш құрылымды 14-3-3 белогымен байланысты болып келеді. Бұл белоктар эукариотты организмдердің барлығында, ашытқы саңырауқұлақтардан бастап жоғары сатыдағы өсімдіктерден және жануарларға дейін табылған. Бұл белоктардың қызметі ДНҚ мен байланысқан транскрипция факторындағы протеинкиназаның активтенуіне қатысады. 14-3-3 белогының рецепторлық қызметі әлі белгілі емес. Басқа жағынан қарағанда, ФКСБ мембранада молекулалық массасы 90 кД болатын полипептид ретінде, ал комплексте молекулалық массасы 30кДа болатын полипептид екендігі туралы мәліметтер бар [17] .
Фузикокциннің мембраналық эффектілері тез жауаптарға жатады әдетте протондар шығуының стимуляциясы фузикокцинді қоса салысымен басталады және лаг фазасы болмайды. Тез мембраналық эффектілермен қатар Фк өсімдіктерде пролингирленуші, генерализдеуші болып әсер ете алады, ал бұл оның гормональды қасиеттері үшін ұтымды. Басқа фитогормондар мен өсуші регуляторға қарағанда (гибберлин) фузикокциннің 2-3 қатарға төмен дозасы (10-20мг/га) назар аудартады. Бабаков жұмысында ФК протеинкиназа активациясындағы ролі қарастырылуда [18] .
Фузикокциннің клеткаларға әсері қызығушылық тудырады. Ең алдымен ФК әсерінен клетка құрамының қышқылдануына қатыстығы туралы айтуға болады. ФК сұлы калеоптилінің клеткасында протондардың шығуын реттейді деген дерек зерттеу жұмыстарында тұңғыш рет көрсетілді. Аталған процестер Н + АТФ-азаның қызметінің реттелуі кезінде немесе плазмалемманың ионды каналдары арқылы иондар ағуының төмендеуі кезінде активтеледі. Шынындада кейінгі мәліметтерге жүгінсек, ФК Н + АТФ-азаның активтілігіне де плазмалемманың калий каналдарының өткізгіштігіне де әсер етеді [19] .
Claudio Olivari қызметкерлерімен тұңғыш рет Н + АТФ-азаның Фк мен активтелуі үшін тағы да басқа белоктың болуын қажет ететіндігін бекітті. Claudio Olivari жұмысында фениарасин оксид Н + АТФ-азаның фузикокцинмен активациясында спецификалық ингибитор болып есептелетіндігі дәлелденген.
Фузикокцин 14-3-3 реттеуші белокпен байланысу арқылы плазматикалық мембранасындағы Н + АТФ-азаның активаторы екендігі анықталды, осылайша Н + АТФ-азаның активациясы тек үштік комплекстің фузикокцин, 14-3-3 белок, Н + АТФ-азаның түзілуінде ғана мүмкін болады. Мүмкін, фузикокцинмен активтенетін Н + АТФ-аза кальций ионына спецификалық болады да, кальций ионының цитоплазмаға тасымалдануын жүзеге асырады. Бұл басқа жағдайлар үшін өте маңызды [18, 20] .
Сұлтанбаев Б. Е. қызметтестерімен жүргізген А 23187 антибиотигімен жүргізген жұмысы бидай дәнінің ұрықсыз бөлігіндегі НАДФ - ГДГ активациясы үшін цитозольды Са 2+ -ң деңгейін реттеп отыруға болады деп тұжырым жасады. Авторлардың цитозольды Са 2+ -ді реттеу бойынша жұмыстары келесідей түрде өткізілді: ионоформен өңдеген ұрықсыз жартыларын 10мМ CaCL 2 бар ерітіндіге орналастырды. Бұл кезде кальций ионофор түзген кальций каналдары арқылы клетканың ішіне еркін өтті. Осылайша цитозольды Са 2+ -ң деңгейі артып отырды. Басқа жағдайларда ионоформен өңделген ұрықсыз жартыларды кальций байланыстырушы оселат ДГТА (этилен гликоль тетраацетат) бар ерітіндіге орналастырды, цитозольды кальций клеткадан ағып шыға бастады да цитозольды кальцидің деңгейі жасанды түрде төмендеді. НАДФ-ГДГ активациясын эмбриональды фактордың көмегімен толығымен басып тастауға болады, егер де ЭГТА-ң көмегімен цитозольды Са 2+ -ң деңгейін жасанды төмендетсе, сол сияқты жасанды түрде цитозольды Са 2+- ң деңгейін арттырса, онда авторлар НАДФ-ГДГ активациясы ешбір факторсыз жүргендігін байқады.
Осылайша, бұл нәтижелер цитозольды Са 2+ деңгейі ғана НАДФ-ГДГ активациясы үшін анықтаушы екендігін көрсетеді. Алайда Сұлтанбаев қызметкерлерімен клеткадағы кальций ионы деңгейінің жоғарылауын қамтамасыз ететін механизмдерді зерттемеді.
Кальций ионының клетка ішіне келуін Са 2+ спецификалық транспортты АТФ-азаның көмегімен плазматикалық мембрана арқылы активті ионды транспорт қамтамасыз етіп отыру керек екендігін дұрыс деп болжады. Сол себепті плазматикалық мембраның протонды кальцилі АТФ-азасының активациясына эмбриональды фактордың эффектілерін зерттеу маңызды тәжірибелердің бірі болып отыр [21] .
Барлық тірі клеткалар ішкі бөліктерді сыртқы ортадан бөлетін жұқа қабықтан тұрады. Ондай қабық плазмалық мембрана деп аталады. Клетка ішіндегі ядро, митохондрия, хлоропластар, гольджи аппараты, эндоплазмалық тор, лизосома сияқты органеллаларды да жеке мембрана қоршап тұрады. Бұл аталған органеллаларды плазма бөліктерінен (ферменттерден, метаболиттерден, т. б. ) мембрана бөліп тұрады.
Клетка өзінің тіршілігін қамтамасыз ету үшін үнемі сыртқы және ішкі мембраналар әр түрлі заттарды тасымалдайды. Мембраналар заттар үшін таңдаулы өткізгіш барьер болып табылады. Мембрана жартылай өткізеді, ол арқылы молекулалар мен иондардың өтуі таңдамалы түрде іске асырылады және ол белсенді түрде реттеліп отырылады. Көптеген заттар белсенді түрде тасымалданып мембрана арқылы ішке енеді. Мембрана липидтерден, белоктардан және көмірсулардан құралады, дегенмен оның негізгі бөлігі липидтер мен белоктар. Клетка мембранасының құрамында белоктар бар. Олардың кейбіреулерінің молекулалары липидтік биқабаттың сыртына орналасады, ал белок молекулалары бүкіл мембрана құрамына енеді [19, 20, 22] .
Плазматикалық мембраналар әр түрлі иондарды іріктеп тасымалдау нәтижесінде клетканың ішіндегі сұйықтардың және сыртқы сұйықтардың иондарының концентрациясының градиенті пайда болады. Бұл иондық концентрация градиентінің нәтижесінде потенциал энергия пайда болады .
Тірі клеткаларда пассивті және активті транспорт болады. Пассивті транспорт дегеніміз концентрация градиенті бойы немесе электрохимиялық градиенті бойы заттардың тасымалдануын айтамыз. Пассивті транспорт липопротеинді комплекс нәтижесінде түзілген арнайы каналдар арқылы (Na, K және басқа каналдар) жүреді. Қанттар, амин қышқылдар және т. б субстраттар арнайы тасымалданушылармен өсімдіктерде, бактерияларда және саңырауқұлақтарда Н + ионымен, ал жануарларда Na + ионымен тасымалданады. Бұл жердегі қозғаушы күш субстрат градиенті емес, иондардың градиенті болып саналады.
... жалғасыСіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz