Рибосома және белок биосинтезінің молекулалық механизмдері мен реттелуі

РИБОСОМА. БЕЛОК БИОСИНТЕЗІ
Клеткада белоктың синтезделуі өте күрделі де, көп сатылы процесс. Қазіргі кезде белок синтезінің өте нәзік механизмі анықталған. Теория жүзінде, мысалы, 12 амин қышқылының өзі ғана кезектесе қосылысып, ақыл жетпейтіндей 10 30 мөлшеріндс алуан түрлі белок молекулаларын кұрастырады. Мұндай санның қанша екенін атап айтудың өзі мүмкін емес. Іс жүзінде клеткада белоктың синтезделуі қалай болса солай тәртіпсіз жүрмейді, ол процесс генетикалық информацияға сәйкес, қатал тәртіппен заңды түрде жүреді екен.
Синтезделетін әр белок клетканың мұктажын өтеу үшін түзілетіні және дені таза организмдс белгілі бір биологиялық қызмет атқаратыны анықталды.
Белок биосинтезі үшін қандай компоненттер қажет? Ол үшін: барлық 20 амин қышқылы, ЛТФ, ГТФ, магний иондары, әр түрлі ферменттер, тРНҚ, иРНҚ барлық түрі, рибосомалар, инициадия факторы, элонгация және терминация факторлары, т. с. с. қажет.
Белоктың синтезделу процесін шамамен негізгі төрт кезеңге бөлуге болады. Олар: 1) амин қышқылдарының активтенуі және амииоацил-тРНҚ түзілуі, бұлар амин кышқылының активті молекулалары болып табылады; 2) белок синтездеуші комплекстің - рибосомалар мен иРНҚ-дан құралған комплекстің түзілуі; 3) полипептидтік тізбектің синтезделуі (трансляция) ; 4) белоктың кеңістіктік құрылымының (конформацияның) түзілуі.
Осы кезеңдердің әрқайсысын жеке-жеке қарастырамыз.
Бірінші кезең. Амин қышқылдарының активтенуі және аминоацил-тРНҚ түзілуі.
Белоктың синтезделу процесі, әдетте, амин қышқылдарының активтенуінен басталады. Олардың активтенуі АТФ энергиясының есебінен және магний иондарының қатысуымен жүреді. Дипептидтік комбинация түзілу үшін орта есеппен 2-3 ккал/моль энергия қажет. Амин қышқылдары карбоксил тобы бойынша амино-ациладенилаттар түзіп, активтенеді:
фермент
R―СН-СООН + АТФ →R-СН-CO-АМФ + пирофосфатNH 2 NH 2
Амин қышқылы Аминоацнладенилат
(активгснген амин қышқылы)
Әр амин қышқылын активтендіретін оның өзіне ғана тән арнайы ферменті бар. Ол фермент аминоацил-тРНҚ-синтетаза деп, метионинді активтендіретін фермент метионил-тРНҚ-синтетаза деп аталады. Бұл фермент процестің келесі сатысына − амино-ациладенилат пен тРНҚ өзара әрекеттесуіне қатысады. Активтелген амин қышқылы аденозиннің З 1 −OH тобы бойынша тРНҚ-ның акцепторлық шетіне жалғасады және аминоацил-тРМҚ түзіледі:
фермснт
аминоациладенилат + тРНҚ → аминоацил-тРНҚ + АМФАминоацил-тРНҚ-синтетаза ферментінің байланысатын екі учаскесі болады. Фермент молекуласының бір учаскесі тиісті амин қышқылын «таниды», екінші учаскесі тРНҚ-ны «таниды», осы тРНҚ-ға сол амин қышқылы ғана ковалентті байланысып жалғасуы тиіс. Бұдан кейін тРНҚ амин қышқылын белок синтездейтін комплекске ауыстырады.
Екінші к е з е ң. Рибосома мен иРНҚ-дан белок синтездеуші комплекстің түзілуі.
80S рибосоманың әрқайсысы екі суббөліктен - 60S және 40S бөліктен кұралады. иРНҚ молекуласы алғашында рибосоманың 40S кіші суббөлігімен байланысады, бұдан кейін оған алғашқы, ииициациялаушы аминоацил-тРНҚ қосылады. Осылайша бастапқы комплекс түзіледі, бұдан әрі қарай бастапкы комплекс рибосоманың 60S суббөлігімен берік байланысады да, белок синтездеуші комплекс түзеді. Әдетте иРНҚ-ға бірнешс рибосома қосылады, синтезделетін белоктың ұзындығына байланысты олар 5-100 шамасындай болады, осылайша полирибосома немесе полисома түзіледі.
Әр рибосоманың екі учаскесі бар (27-сурет) : оның біреуі полипептидтің өсіп келе жатқан тізбегімен байланысады және ол пептидильдік (П) учаске деп аталады. Ал екінші учаскесі жаңа амидоацил-тРНҚ-ны қосып алады және оған сәйкес аминоацил (А) учаскесі деп аталады.
Ү ш і н ш і к с з е ң. Полипептидтік тізбектің синтезі (трансляция) .
Бұл белок биосинтезіндсгі аса маңызды процестердің біреуі - трансляция үш сатыға бөлінеді: а) инициация немесе полипептидтік тізбек синтезінің бастамасы; б) элонгация немесе полипептидтік тізбектің ұзаруы; в) терминация немесе пептидтік тізбек синтезінің аяқталуы.
Инициация. Көпшілік белоктардағы пептидтік тізбек синтезінің инициация (басталуы) метионин амин қышқылынан басталатыны анықталған. Бактерия клеткасына қатысты инициациялаушы амии қышқылы формилметионин, ал эукариоттық клеткага қатысты белок синтезіне ең алдымен мотионин (формильдік топсыз) кіріседі. Мұнда Инициациялаушы тРНҚ активтенген ме-тионинді синтездеу орнына жеткізеді. Бұл амин қышқылына АУГ кодоны сәйкес коледі, ол кодон әр иРНҚ молекуласының бастапқы жағында орналасуы тиіс.
Инициациялаушы метионин-тРНҚ рибосоманың П-учаскесімеи былай байланысады: тРНҚ-ның антикодоны иРНҚ-ның инициациялаушы кодонымен үйлесімді байланысады, инициациялаушы комплекс осылайша пайда болады.
Бұл процесте инициациялаушы белоктың үш фактор іҒ 1 , іҒ 2 , іҒз және энергия көзі ретінде ГТФ қатысады. Инициация факторларының қызметі дәл анықталған жоқ. Синтез басталғаннан кейін инициация факторлары босайды да, жаңа инициация үшін қайтадан пайдаланылады. ГТФ гидролизденеді, бұл кездс инициация үшін энергия бөлініп шығады.
Элонгация. Инициациялаушы комплекс түзілгеннен кейін бірден-ақ пентидтік тізбек ұзара бастайды (элонгация) . Инициациялаушы комплексте метионинге жаңа амид кышқылдары кезектесіп қосылыса береді, оларды тРНҚ алып келеді.
иРНҚ молекуласы бойында амии қышқылдары қалай кезектесіп, белгілі бір тәртіппен орналасады? Бұл - белок биосинтезіндегі өзекті мәселелердің бірі. Көптеген ел ғалымдарының күш жұмсауы нәтижесінде бұл мәселе толық шешілген. Клеткада делдалдық қызмет атқаратын аралық молекулалар болады екен. Сондай молекулалардың көмегімен болашақ белоктың өсіп келе жатқан полипептидтік тізбегінде амин кышқылдары өз орнын дұрыс таба алады. Антикодон ілгегі бар тРНҚ осындай адапторлық делдал болады. Үйлесімді сәйкестік принципі бо-йынша тРНҚ антикодоны иРНК кодонын «таниды» да, сутектік байланыс арқылы жалғасады. Антикодонның кодонды осылай «тануы» генетикалық мәліметті трансляциялауда аса маңызды болып табылады.
Мұнда тРНҚ молекуласы екі бірдей қызмет атқарады: саны 20 амин қышқылдардың ішінен «өзінің» амин қышкылын таңдап алады және антикодонның көмегімен иРНҚ-дағы сәйкес кодонды табады.
Элонгация процесін елсстету үшін пептид синтезін қарастырып көрейік. Мұнда инициациялаушы метионил-тРНҚ кіші суббөлікке келеді. Ол бөлікпен иРНҚ байланысқан және антикодон-кодон байланысы қалыптасады. Осымен бір мезгілде метионин амин қышқылы үлкен суббөліктің П-учаскесімен байланысады. Бұдан кейін бірден-ақ АУГ кодоны бір адым жылжиды, яғни кіші суббөлікте бір триплет қозғалады. Осы кезде кіші суббөлікте антикодонмен байланысуға дайын келесі ГЦУ кодоны тұрады. ГЦУ кодонына аптикодон аланил-тРНҚ келеді және үлкен суббөліктің А-учаскесіне аланин енеді. Мет пен Ала екеуінің арасында пептидтік байланыс қалыптасады да, тРНҚ босап шығады. Екінші кодон ГЦУ тағы бір қадам жылжыған кезде келесі кодон үшін кіші суббөлік босанады. Осы кезде фенилаланиннің кодон-аптикодоны бірігіп кабысады. Үлкен суббөліктің А учаскесінде фенилаланин амин кышқылы келеді. Ала меп Фен екеуінің арасында пептидтік байланыс қалыптасады. Осылайша бір триплет соңынан екінші триплет жалғасып, поли-пептидтік тізбек ұзарады (элонгация) . Полипептидтік тізбек біртіндеп кезектесіп N-соңғы бос амин тобынан ( −NH 2 ) карбоксильдік соңғы топқа (−СООH) қарай өседі.
Полипептидилсинтетаза фсрментінің амин кышқылдарын өзара байланыстыруы және пайдаланылған тРНҚ-ның босануы П-учаскесіндс іске асады.
Тәжірибс жүзінде полипептид синтезі кезінде рибосомалардың біртіндеп бір қадамның соңынан екінші қадам басып, яғни әр жолы бір триплет ұзындығындай жылжып, иРНҚ молекуласының бойымси қозғалатыны дәлелденген. Мұндай процестің нәтижесінде үздіксіз полипептидтік тізбек түзіледі. иРНҚ молекуласының бойымсн рибосомалардың қозғалуы (транслокация) элонгация факторының εҒ 2 әсерінен және ГТФ молекуласының гидролизденуінен шыққан энергия есебінен іске асады. Клеткада элонгацияның белоктық екі факторы εҒ 1 және εҒ 2 болады. Оның біреуі А-учаскесінде аминоацил-тРНҚ фиксациясына қатысады, ал екіншісі жоғарыда айтылғандай, рибосома транслокациясына қатысады.
Терминация. Полипептидтік тізбек биосинтезінің соңғы сатысы
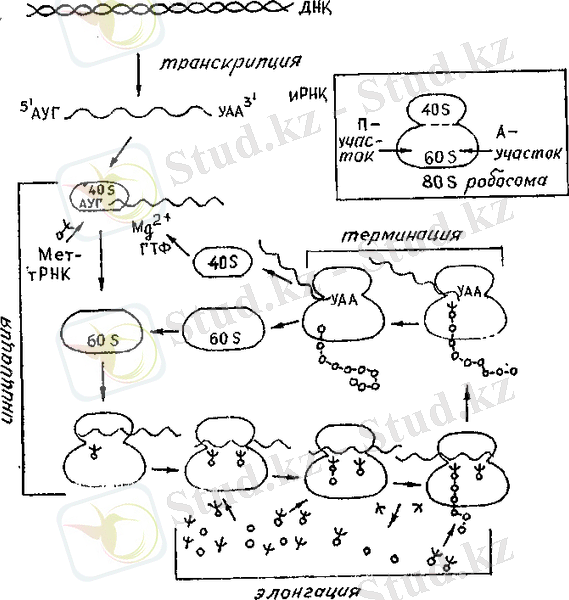
Белок биосинтезінің жалпы схемасы.
РНҚ молекуласында инициациялаушы кодон ЛУГ 5 1 - соңында және тер-миндеуші кодон УАЛ З 1 ―соңында көрсетілген. ІІІартты белгілер: ψ-тРНҚ, о-о-о - аминқышқылдары, ψ - аминоацил - тРНҚ о-о-о-о-о-о - полипептидтік тізбек.
Жоғарыда oң жақта 80S рибосоманың құрылым схемасы берілген.
терминация болып табылады. Рибосомалар иРНҚ молекуласының бүкіл ұзыны бойынан жүріп өтіп, ондағы терминация сигналына (белгісіне) жеткенше, генетикалық информация трансляциясы созыла береді. Полипептид синтезінің аяқталу белгісі УАА, УАГ немесе УГА сияқты мағынасыз кодондардың біреуі болады. Терминацияның белоктық факторы осы кодондардың біреуін «таниды» да, элонгацияны тоқтатады, сөйтіп полипептидтер синтезі доғарылады. Белок синтездеуші комплекстен бөлініп шығады, рибосомалар суббөліктерге диссоциацияланады да, иРНҚ ыдырап бұзылады. Полипептидтің С-соңы мен иРНҚ екеуінің арасындағы байланыстың ажырауын рибосомалық фермент пептидилтрансфераза катализдейді.
Клеткадағы белок синтезінід схемасы жоғарыдағы суретте берілгсн.
Т ө р т і н ш і к е з е ң. Белоктың кеңістіктік құрылымының (кон-
формациясының) қалыптасуы.
Біз организмдегі генетикалық информация полипептидтік тізбекте амин қышқылдарының орналасу ретін қалай дәл белгілейтініне қатысты мәселені, яғни белоктың бірінші реттік құрылымын қарастырдық. Егер ксз-келген белок молекуласының тиісті кеңістіктік кұрылымы - үшінші реттік немесе төртінші реттік құрылымы болса ғана ол өзіне тән міндетті атқара алады.
Белоктың бірінші реттік құрылымы мен оның конформациясы екеуінің арасындағы тығыз байланыс анықталған. Бұл жағдай мынаны көрсетеді: геннің құрылымы белоктың кызметін, оның биологиялық қасиеттерін және басқа да ерекшеліктерін алдын ала белгілейді.
Тәжірибе жүзінде дәлелденгендей, белок конформациясының қалыптасуы рибосомалардагы полипептидтер синтезі кезінде басталады. Өсіп келе жатқан полипептид тізбегі біртіндей бүктеледі, өзінің синтезделуі аяқталғанға дейін үш өлшемдік құрылымға ие болады. Полипептид синтезі аяқталғаннан кейін молекула энергетикалық тұрғыдан ең тиімді күйге көшіп, өзінен-өзі тұрақты да сәйкес конформацияға түседі.
Белок конформациясының қалыптасуында амин қышкылдарындағы бүйірлік топтар, олардың полярлығы және полярсыздығын айтарлықтай қызмет атқарады. Полярлық және полярсыз амин қышқылдарының caн қатынасына және тізбектегі олардың орнына байланысты, қалыптасқан белоктың үшінші реттік құрылымы, олардың ылғалды ортадағы ерекше беріктігін қамтамасыз етеді. Өйткені белок ылғалды ортада болады.
Простетикалық топтары бар күрделі белоктар конформациясының қалыптасуы да баяндалғандай болады. Углеводтық, липидтік, қышқылдық және басқа да простетикалық топтар синтездің барысында полипептидтік тізбекке қосылып жалғасады. Мұндағы ең маңызды құбылыс - синтез біткеннен кейін олардың қосылуы да тоқталады.
Белок биосинтезінің реттелуі
Белок биосинтезінің жылдамдығын бір жағынан генетикалық аппарат басқарса, екінші жағынан сыртқы орта факторлары басқарады. Эукариот клеткаларында белок синтезі реттелуінің екі механизмі болуы мүмкін. Олар: транскрипция деңгейінде реттелу және трансляция деңгейінде реттелу.
... жалғасыСіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz