Бидай алейрон қабаттарындағы альдегидоксидаза ферментінің гиббереллин мен абсциз қышқылы арқылы реттелуі

Мазмұны
Кіріспе 3
1 Әдеби шолу 5
1. 1 Бидай дәніне жалпы сипаттама 5
1. 1. 1 Дәннің құрлысы 5
1. 1. 2 Алейрон қабаты 7
1. 2 Фитогормондарға сипттама 10
1. 2. 1 Абсциз қышқылы 11
1. 2. 2 Гиббереллиндер 12
1. 3 Альдегидоксидаза ферментіне жалпы сипаттама 16
3 Пайдаланған әдебиеттер тізімі 18
.
Кіріспе.
Өсімдік клеткалардың программаланған өлімінің немесе апоптоздың генетикалық механизмін зерттеу қазіргі заманғы молекулалық биология және генетиканың өзекті проблемаларының бірі болып табылады. Клетканың дифференцияциясың фундаментальді механизмін ашуға, өсімдіктің дамуы мен өсуін сонымен қатар өсімдіктің өнімділігін жоғарылату әр түрлі ортаның биотикалық және абиотикалық факторларына төзімді және өнімділігі жоғары ауылшаруашылық өнімдерін алу жолында Өсімдік клеткалардың программаланған өлімінің молекулалық және генетикалық негіздерін зерттеу өте маңызды. Жоғарыдағы мәселені зерттеу соңғы жылдарда интенсивті түрде дамып келе жатқан және биологиялық өнімнің қалыптасуын шешуші факторлардың бірі. Қазіргі заманғы молекулалық әдістер сигналдық жүйелерін реттеуші гормондар-Гиббериллин (ГҚ) және абсциз қышқылының (АБҚ) нюанстарын толығымен зерттеп ашуға болады. Астық тұқымдастардың Алейрон клеткаларының ультра құрылымы олардың қызметін көрсетеді. Дәндегі алейронның көптеген көрсеткіштері оның ерекше және жан-жақты тәжірибелік жүйе болуын қамтамасыз етеді. Астық тұқымдастарының алейрон қабаты біртекті, жоғары дифференциалданған клеткалардың бір қабатынан (сұлы, жүгері, қара бидай, бидай) немесе бірнеше қабатынан (күріш, арпа) тұрады.
Алейрон қабатының онтогенетикалық программаланған клеткалар өлімі (ПКӨ) апоптотикалық сипатта жүзеге асатыны анықталған. Бұл процестің белсенділігінің артуымен, геномдық ДНҚ молекуласының фрагментациялануымен үйлесіп, фитогормондар - гиббереллин және абсциз қышқылдарымен бақыланылатыны көрсетілген Жоғарыда айтылғандай, алейрон клеткалары - жоғары деңгейде дифференциацияланған ұлпа. Бұл ұлпаның негізгі қызметі - гидролитикалық ферменттерді (альфа-амилазаны қоса) синтездеп, крахмалды эндоспермге секрециялау. ГҚ және АБҚ гормондарының Клетканың программаланған өліміндегі рөлі анықталған. ГҚ клеткалардың өлімін жеделдетеді, ал АБҚ бұл әсерді тежейді. Клеткада О2 радикалының көзі ретінде плазматикалық мембрананың NADPH оксидаза болады. Н2О2өсімдіктерде клетка қабырғасының пероксидазасы және щавель қышқылының оксидазасы көмегімен пайда болады. Н 2 О 2 плазматикалық мембрананың NADPH оксидазасы экспрессиясының индукторы болып табылады. Н 2 О 2 суперорксиддисмутаза көмегімен бір электронды О2 қалпына келтіріп, О2 супероксидті анион радикалының пайда болу нәтижесінде Н2О2 клеткада генерацияланады. Альдегидоксидаза ферменттер оттегі радикалдарының негізгі көздері болып табылады Альдегидоксидаза өсімдіктерде әр түрлі спецификалық субстраттары бар ферменттер топтарығ және өсімдік организмінде метаболиттік рөл атқарады. Құрамында молибдені бар фермент альдегидоксидаза каталитикалық реакция үшін оттегін пайдаланып, нәтижесінде супероксид радикалы пайда болады. АО АБК синтезінің кілтті ферменті, ал АБК бастамасы ксантофилдерден көрінеді. Бұл заттардың фитогормонға айналуына 5 ферменттер қатысады. Оның екеуі 9- (ЭКДО) және альдегидоксидаза каталитикалық реакция үшін оттегін пайдаланады. Нәтижесінде, аралық өнім суперорксид анион пайда болады. Жоғарыдағы мәліметтерге сүйенетін болсақ, оттегінің белсенді түрлері митохондриальді тізбектегі электрондардың тасымалдануы нәтижесінде және оксидазалардың көмегімен жүреді.
Мақсаты: Бидай алейрон қабаттарындағы альдегидоксидаза ферментінің гормондар арқылы реттелуін зерттеу
Міндеті:
Бидай алейрон ұлпаларындағы альдегидоксидаза ферментінің АБҚ гормондар арқылы реттелуін зерттеу
Бидай алейрон қабаттарындағы альдегидоксидаза ферментінің ГҚ гормондар арқылы реттелуін зерттеу
1. Әдеби шолу.
1. 1. Бидай дәніне жалпы сипаттама.
1. 1. 1. Дәннің құрлысы.
Дәннің формасы: шар тәрізді(горох), ұршық тәрізді және көп гранулалы болады.
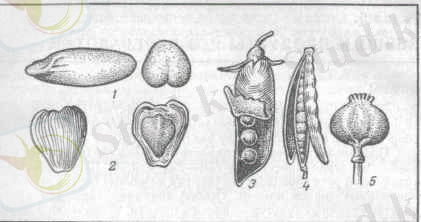
Қара бидай, сұлы, арпа секілді дәнді-дақылдарды еккенде аударылған жерге құрсақ бөліміне қарама-қарсы бөлігі жерге түсірілген болады.
Дән - сыртқы қабықтан, алейрон қабатынан, ұрықтан, эндоспермадан тұрады.
Бидай, қара бидай және жүгері жалаңаш дәнді-дақыл куьтурасына жатқызылады.
Дәннің жемістік және ұрықтық қабыршақтары ұрықты механикалық бұзылудан сақтайды. Олардың құрамына дәннің бояуын көрсететін клетчатка, пентозандар, минералдық және пигменттік заттар кіреді.
Дәннің ұрығы клетканың негізгі ұрық қалқанынан айырылған тірі клеткадан тұрады. Оның химиялық құрамы белок, қантқа, липидке, минералды тұздарға сонымен қатар, активті заттарға(витамин, фермент) бай болып келеді. Дән салмағының 1, 5-10% ұрық бөлігі құрайды.
Дәннің ең үлкен бөлігін эндосперм құрайды, шамамен 75-80%. Белок пен крхмалдан тұратын ірі клеткалар ұнды бөлікке жатқызылады.
Қабық пен эндосперм арасында, реттелген клеткалардан құралған алейрон қабаты болады. Алейрон қабатына қиын қорытылатын белок пен майлар кіреді. Дән салмағының 7-12% алейрон қабаты құрайды.
Ұзыннан кесілген бидай дәні:
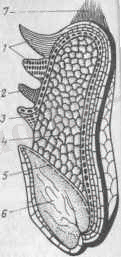
1-сурет
1-Жемісті қабық; 2-тұқымдық қабық; 3-алейрон қабаты; 4-эндосперм; 5-ұрықтың қалқаны; 6-ұрық; 7-шапқысы(бородка) .
Дәннің химиялық құрамы:
Дәннің дақылдық қоректік құрамы түрге, сортқа, дәнді жинау уақытымен әдісіне, жетілдіру тәсіліне, сақталуына және тағы басқа факторларға байланысты.
Көмірсулар - дәннің қоры және құрғақ заттың көп бөлігін құрайды. Олар крахмал, клетчатка, гемицелюлоза, қант түрінде кездеседі.
Крахмал - дәннің 36-59%-ға деиін кездеседі. Эндоспермде және тұқыдықта ұрық тәрізді болып келеді. Дәннің қалған анатомиялық бөліктерінде крахмал табылмаған. Крахмал формасы және көлемі дәннің түрі мен дақылға байланысты.
Клетчатка - дәннің 1, 5-11%дейінгі арлықта болады. Ол гүлденген жұқа қабықта, өнім және де аз мөлшерде алейрон қабатында, ұрықта, эндоспермде кездеседі.
Гемицелюлоза - дән клетчаткасында 4-10%-ға дейін кездеседі.
Қант - дәнде 1, 2-9, 0% болады. Негізгі жиналу ұрықта, жемісте, ұрықтық қабықта жиналады.
Азоттық заттар - 9:1 қатынаста белок және белоктық емес заттар күйінде болады.
Бидайдың көп бөлігі белокпен байланысқан, сәйкесінше ұн да суда ерімейді, сондықтан қамыр да сумен байланысып, ол ашуға және ұлпа құруға қабілетті, және оны сумен жуу арқылы тез бөліп алуға болады. Ол созылмалы ақ, сары немесе сұршыл түрінде масса болады.
Липид бидайда 1, 6% болады.
Дәнді дақылдың май молекуласының құрамына қанықпаған май қышқылының көп бөлігі кіреді, соның ішінде жоғары биологиялық құндылыққа ие ауыстырылмайтын (линол, линолен) май қышқылдары.
Сонымен қатар қышқылдың көп мөлшерде болуы дәндегі майлардың тез қышқылдануына әкеледі, және өнімнің қайта өңдеуінің сақталу кезінде оның ашылануын тудырады. Дәнде май біркелкі орналаспаған, ол негізінде ұрықта және алейрон қабатында болады.
Дәнде ферменттер гидролаза түрінде кездеснді. Гидролаза - ақуыз, май, көмірсулардың еруіне қатысады, және ұрық пен алейрон қабатында табылатын фермент. Зиянсыз дәнде ферменттер активсіз, бірақ оның дымқылданған және өсу кезінде, ферменттердің активтілігі кенеттен өсіп, бұзылуға алып келеді.
Минералдық заттар гүлденген жұқа қабаттың, жемісті және ұрықтық қабаттың, алейрон қабатының құрамына кіреді. Олар эндосперманың құрамында да болады, бірақ өте аз мөлшерде. Дәндегі минералдық заттардың мөлшері 1, 2-5%-ға дейін. Олар кальций, калий, натрий, магний, темір, фосфор және тағы басқа түрлерде кездеседі.
В(В1, В2, В6), РР витаминдері кездеседі. Май қышқылдары Е (токоферол) витамині кездеседі және кариотин витамині аз мөлшерде кездеседі. Ұрық пен алейрон қабаты витаминдерге бай.
Таблица 1.
Дәннің химиялық құрамы:
1. 1. 2. Алейрон қабаты
Алейрон қабаты ұрықтың дәнге жетілу кезеңінде қоректік қор ретінде пайдаланылады. Дәннің пісіп жетілу кезеңінде, судың жетіспеушілігін жойып, вакуоль протопластына өту арқылы қатты белоктардың өнуін қамтамасыз етеді. Алейрондық қабат майда дәнге толы бір реттік ірі қабатты клеткалардан тұрады. Алейрон қабатының құрамына шамамен 40% белок (альбумин, глобулин), 10% май, 6% қант, 10% клетчатка, 10-13% зол, суда ерйтін витминдер, гемицелюлозалар, пентозандар кіреді. Дәннің таза салмағының 4-9% құрайды. Алейрон дәндері - протеин дәндері, дәнді-дақылды өсімдіктерде шар тәрізді, түссіз болып кездеседі.
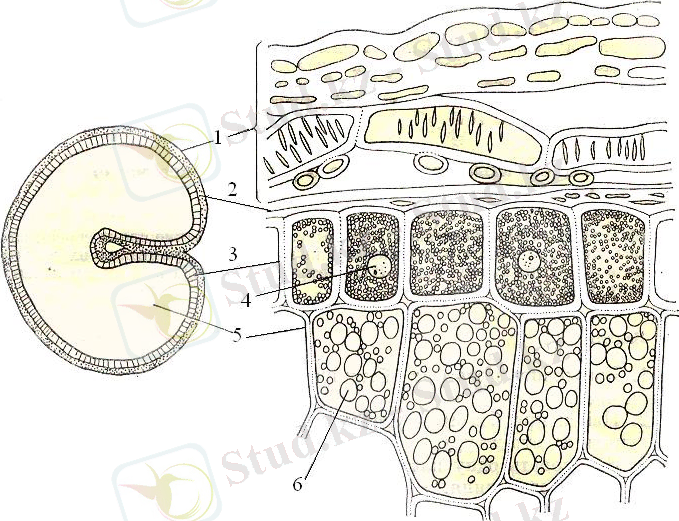
Алейрон қабаты - бұл эндоспермнің сыртқы қабаты, бірақ ұн тартқыштар (мукомолы) оны қабықшаның бір бөлігі деп қарастырады. Бидайдың алейрондық қабаты бір қатарға орналасқан ірі қалың қабатты клеткалардан тұрады және дәннің крахмалды эндосперм мен ұрықтың бетін толығымен жауып тұрады. Сыртқы жағынан алейрондық қабат гиалинді қабатпен тығыз байланысқан; ішкі жағынан ол крахмалды эндосперммен байланысқан. Алейрон қабатының клеткалары ұзынынан және көлденеңінен кесіндісінде төртбұрышты немесе сәл созылған пішінде болады. Көптеген мәліметтер бойынша, алейрондық қабаттың қалыңдығы шамамен 65-70 μ болады. Алейрон қабатының клетка қабырғасы негізінен клетчаткадан тұрады. Алейрон қабатының әрбір клеткаларында ірі ядросы болады (сурет-3) .
Алейрон қабаты негізінен бір қабат клеткалардан тұрғанымен, кейде бір клетканың орнына екі клетка орналасуы мүмкін. Фогольдің мәліметтері бойынша алейрондық қабаттың ішкі клеткаларында крахмалдың біршама мөлшері болады. Клеткалардың әртүрлі қалыңдығына байланысты алейрондық қабаттың ішкі беті тегіс емес болады және бұл жағдай қабықтардан эндоспермді тазалау кезінде бірқатар қиындықтар тудырады [10] .
Бидай дәнінің эндоспермі тозаң қапшығында, орталық клетканың аталық жыныс клеткасымен ұрықтану нәтижесінде пайда болатын, триплоидты ұлпа. Тозаңдану процесінен кейін шамамен 8-10 тәуліктен соң, дән эндоспермі крахмалды эндоспермге және алейрон қабатына дифференциацияланады [7] . Алайда, ұрықтанған бір клеткадан пайда болатынына қарамастан, алейрон ұлпасы мен крахмалды эндосперм клеткаларының өлуі әртүрлі уақытта жүзеге асады. Крахмалды эндосперм клеткалары бидай дәнінің пісіп-жетілу барысында өліп, дән қуысын белокты және крахмалды гранулалар түрінде толтырып тұрады деген болжам бар. Ал алейрон клеткалары, дәннің пісіп-жетілу сатысында еш өзгеріссіз сақталып, тек қана оның өніп-өсу сатысын, гидролитикалық ферменттерді синтездеп, секрециялау арқылы
индукциялағаннан кейін ғана элиминацияланады [11] .
2-сурет.
1-ұрыққа жақын бөлігі, 2-дән қабығы, 3-алейрон қабаты, 4-ядро, 5-крахмал дәндері бар эндосперм клеткалары, 6-крахмал дәндері
Алейрон қабатының онтогенетикалық программаланған клеткалар өлімі (ПКӨ) апоптотикалық сипатта жүзеге асатыны анықталған [12-14] . Бұл процестің белсенділігінің артуымен, геномдық ДНҚ молекуласының фрагментациялануымен үйлесіп, фитогормондар - гиббереллин және абсциз қышқылдарымен бақыланылатыны көрсетілген [15] .
Алейрон клеткаларының ультра құрылымы олардың қызметін көрсетеді. Дәндегі алейронның көптеген көрсеткіштері оның ерекше және жан-жақты тәжірибелік жүйе болуын қамтамассыз етеді. Астық тұқымдастарының алейрон қабаты біртекті, жоғары дифференциалданған клеткалардың бір қабатынан (сұлы, жүгері, қара бидай, бидай) немесе бірнеше қабатынан (күріш, арпа) тұрады [16] . Пісіп-жетілген құрғақ алейрон клеткалары қалың геммицеллюлозалық қабырғамен қоршалған және олар ешқандай да клеткалық бөлінуге ұшырамайды. Алейрон қабаттарын жабысып қалған өлі крахмалды эндоспермнен ажыратып алуға болады және ажыратып алынған алейрон ұлпасынан ферменттердің көмегімен протопластардың біртекті популяциясын дайындауға да болады [17] . Осындай жолмен жекеленіп алынған протопластар, тұтас алейрон ұлпалары сияқты гиббереллинге тәуелді α-амилаза изоферментінің активтенуі және синтезін қамтамасыз ететіні көрсетілген. Жекеленген протопластар алу әдістемесі сұлы және арпа алейрон ұлпасына қатысты жасалынған [18] . Біздің лабораторияда бидай алейрон ұлпаларынан жекеленген протопластарды бөліп алудың және тазалау, оларды концентрлеу үшін фракциялаудың модификацияланған әдістемесі ұсынылған [19] . Ферменттік өңдеу арқылы алынған бидай алейрон протопластарының шығымы шикі ұлпаға шаққанда -35х10 6 клетка/1 г болды. Протопластарды фиколл градиенттінде центрифугалау әдістемесі тіршілік дәрежесі жоғары клеткалар фракциясын алуға мүмкіндік берді (шамамен 80-85 %) .
Алейрон клеткаларының цитоплазмасына белоктар қорынан тұратын көптеген вакуольдер тән, оларды көбінесе алейрондық түйіршіктер деп те атайды, олар бүкіл клеткаларды алып жатады. Бұл органелла алейронның гормондар әсеріне қайтаратын реакциясында негізгі қызмет атқарады. Протеиндердің қоры бар вакуольде гиббереллинмен өңделгеннен кейін гидролизге ұшыраған протеиндерді секреторлық белоктардың түзілуіне қажетті аминқышқылдармен қамтамасыз ету үшін қор ретінде сақтай бастайды [20] . Протеиндердің қоры бар вакуольдер, сондай-ақ, астық тұқымдастардың дәндеріндегі минералдардың негізгі қоймасы болып табылады. Олардың құрамынан минералдар фитин түрінде бөлініп алынған. Фитин К + , Mg 2+ , және Ca 2+ иондары мен фитинді қышқылынан (гексофосфат) түзілген кристалды, ерімейтін комплекс болып табылады. Бұл кристалды қосынды, глобоид деп те аталады. Рентгендік микроанализ нәтижелері бойынша дән құрамындағы РО 4 3+ , К + , Mg 2+ және Ca 2+ иондарының шамамен 75% алейрон қабаттарында сақталады [21] . Алейрон немесе протопластардың инкубациялық кезеңінде вакуольдер көлемдері ұлғайып, бір-бірімен орталық үлкен бір вакуоль түзілгенше қосыла бастайды. Инкубациялық ортада гиббереллин болған жағдайда клеткалар мен протопластардағы алейрон қабаттарының вакуольдену процесі жылдамдай түседі. Бейтарап майларды сақтайтын липидтік қосындылар немесе олеосомдар да алейрон клеткаларының маңызды органеллалары болып табылады, олар клетканың бүкіл кеңістігінің 30% алып жатқан үшглицеридтік негізден тұрады [22] . Алейрон клеткаларындағы олеосомдар эндоплазмалық торда пайда болып, кейіннен эндоплазмалық торға және протеиндердің қоры бар вакуольдер бетіне бекінген күйде болады. Олеосомдардың вакуольдер бетіне бекініп жатуы вакуолдердің кейбір түрлері тікелей эндоплазмалық тордан пайда болады деген болжамды растайды. Қор ретінде сақталған бейтарап майлар алейрон клеткаларын көміртегі көзімен де қамтамасыз етеді, эндоспермді мобилизациялау басталғанға дейін клетка көміртегіні осы жерден жұмсай алады, сонымен бірге мұндағы май қышқылдары мембрананы түзуге пайдаланылады. Бұл май қышқылдарының алейрон клеткалары арқылы метаболизденетініне бірнеше дәлелдер бар. Алейрон клеткаларында малатсинтетазадан және изоцитрат лиазадан тұратын көптеген глюкоксисомалар да болады [23; 24] және оқшауланып алынған алейрон қабаттары сахарозаны синтездей алады.
Жоғарыда айтылғандай, алейрон клеткалары - жоғары деңгейде дифференциаланған ұлпа. Бұл ұлпаның негізгі қызметі - гидролитикалық ферменттерді синтездеп, крахмалды эндоспермге секрециялау. Олай болса, алейрон ұлпаларында оның секреторлық қызметіне байланысты эндоплазматикалық тор, Гольджи аппараты, митохондриялар өте жақсы дамыған.
1. 2. Фитогормондарға жалпы сипаттама
Фитогормондар - төменгі молекулалы, өсімдіктерді өндіруші және реттегіш қызметі бар органикалық заттар. Фитогормон төмен концентрацияда (10-11М) әсер етіп, өсімдіктің сезу және қозғалысына әртүлі морфологиялық және физиологиялық өзгерістер әкеледі.
Жануарлармен салыстырғанда өсімдіктерде арнайы гормон синтездеуші органдары болмағандықтан, кейбір органдар фитогормондармен қаныққан күйінде болуын ескеру керек. Сабақтың жоғары меристемасының бөлігі ауксинге бай, жапырақтары гиббереллинге, тамыр және жетілуші дән цитокининге бай. Фитогормондар кең спектрлі қызмет атқарады. Дәннің өсуі, бойының өсуі, ұлпа және органдардың дифференциялануы, гүлденуі, жемістің жетілуі сияқты өсімдіктің тіршілік әрекетін фитогормондар реттейді. Өсімдіктің белгілі бір органында фитогормон түзіліп, басқа органға тасымалдануға қабілетті.
Жалпы қасиеттері . Химиялық бйланыстар белгілі бір өсімдік бөлігін және қызметін басқа органда өңдеп, өзінің әсерін аз ғана концентрацияда көрсетеді, өсу даму процесіне спецификалық қызмет атқару қасиетіне ие, осы тұжырым «клетка - гендік модельді қабылдаушы» дегенді растайды. Сонымен қатар фитогомон әсері белгілі бір мөлшерде ішкі және сыртқы факторлардың шиеленісуімен анықталады.
Классификациясы және структурасы . Жалпы классификациясы, өсімдіктер арасында гормондарды 5 негізгі топқа бөлінеді. Әртүрлі өсімдік гормондары бір-бірімен химиялық структурасы бойынша ажыратыллады. Сондықтанда әсеріне байланысты өсімдік физиологиясы және жалпы химиялық құрлысына негізделіп гормондарды топтастыруға болады. Бірақ, кейбір физиологиялық активті заттар бірде бір класқа жатпайды. Әр класс өзіне стимулятор қосып, әртүрлі функцияның ингибиторы сияқты жұп жұбымен жұмыс жасайды. Бұл жағдайда бір немесе бірнеше заттардың концетрациясы анықталынады, өсімдіктің соңғы өсу әсерінен.
Негізгі топтар:
Абсциз қышқылыЦитокининдер Этилен қышқылыГиббереллиндер
1. 2. 1. Абсциз қышқылы
Абсциз қышқылы( таблица-2 ) - абсцизин, дормин - бұл өсімдік гормоны.
Қызметі. Алғашқы кезде жапырақтың түсуінде АБҚ рөлі байқалған, бірақ қазіргі кезде мұндай рөл кейбір өсімдіктерде стреске және патогенді жауап беретін сигналды жолдармен байланысқан АБҚ жауапты.
АБҚ биоситезі жүретін қайсыбір өнімде ген және секвенирленген ген табылған. Өсімдіктерден өзгеше жолмен кейбір патогенді саңырауқұлақтарда АБҚ синтезделеді.
Жапырақтың түсуіндегі АБҚ рөлі көрсетілген қысқа дайындық кезеңінде АБҚ өсімдік бүршігінің ұшында синтезделеді. Бұл период өсуді тежейді, жапырақшылардан қорғаушы қабыршақ түзіледі, суық периодта ұйқылы бүршікте жабылады. АБҚ комбий клеткаларының бөлінуін бірінші және екінші өсуді тоқтатады.
Сонымен қатар АБҚ өсімдіктерінің тамырында, стрестік факторларда түзіледі. Одан кейін саңылау клеткалары осмотикалық потенциалды өзгертіп және саңылаудың жабуын тудыратын күйде жеткізетін АБҚ жапыраққа енеді. Саңылаудың жабылуы транспирацияны тежеп және жапырақ арқылы судың жойылуын тоқтатады.
АБҚ түзілуінің орны және уақыты :
өсімдіктерді кептірудегі бүршіктің тығыздалу кезеңінде; жасыл жеміс жидектер және қысқы период алдындағы дәнде; тамырдан жапырақ ксилема жолдарынан тез тасмалданады; қоршаған ортаның стрестік факторларына синтездік жауап береді; өсімдіктің барлық органдарында тамырда, гүлде, жапырақта, сабақта синтезделеді. АБҚ әсері:
1. 2. 2. Гиббереллиндер.
... жалғасыСіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz