Липидтердің алмасуы және биохимиялық реттелуі

әл-Фараби атындағы Қазақ ұлттық университеті Биология факультеті Биотехнология мамандығы
Тақырыбы: Липидтердің метаболизмі
Тексерген: б. ғ. д., профессор Бейсембаева Р. У.
Орындаған: І курс магистранты Карибозов Ерғұлан
Алматы, 2009
Мазмұны
Липидтердің алмасуы
Липидтердің биологиялық қызметі ең алдымен олардың энергия көзі болатындығымен анықталады. Бұл функцияны майлар ыдырағаннан кейін бөлініп шығатын май қышқылдары атқарады. Фосфолипидтер, гликолипидтер және холестерин клетка мембранасының түзілуге қатысады. Полиқанықпаған май қышқылдарының кейбір туындылары (простогландиндер) реттегіш қызмет атқарады, бұл май қышқылдары ауыстырылмайтын тағамдық факторлар болып табылады. Холестерин мембрананың құрылымдық компоненті, сонымен қатар өт қышқылдары мен стероидтық гормондардың алғы заты.
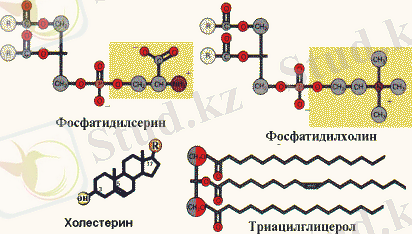
Май қышқылдары, майлар, стероидтар мен фосфолипидтердің құрылымы
Сонымен қатар, организмге майлармен бірге майда еритін витаминдер (А, Е, D, К) түседі. Айта кететін жайт, гидрофобтылық - барлық липидтердің ортақ қасиеті. Бірақ, кейбір липидтер (гликолипидтер, фосфолипидтер, өт қышқылдары) амфифильді, өйткені оның құрамында гидрофильді және гидрофобты аймақтары бар.
Липидтердің асқазан - ішек жолында ыдырауы
Липидтердің ыдырауы 12 елі ішекте жүзеге асады, онда ұйқы безінің сөлі бар липаза және өт құрамындағы коньюгацияланған өт қышқылы түседі. Эмульгацияланған майлар - астың қорытылуына қажетті жағдай, өйткені гидрофобты субстратты гидролитикалық ферменттер - липазалардың әсер етуін қолайы жағдай жасайды. Эмульгациялану өзінің амфифильдік қасиетіне байланысты, май тамшысын қоршайды және беттік натяжение төмендетіп, майдың бөлшектенуіне әкелетін май қышқылдарының қатысуымен жүзеге асады.
Майлардың гидролизі майдың тамшысының бетінде сорбцияланып, триацилглицериндердің (ТАГ) эфирлі байланыстарын ыдырататын панкреатикалық липазалардың қатысуымен жүреді. Май қышқылдарының ыдырауы ең алдымент α-жағдайынан басталады. Нәтижесінде диацилглицерин, кейін гидролиздің негізгі өнімі болып табылатын β-моноацилглицерин түзіледі:
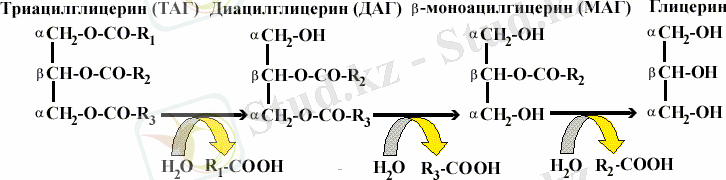
Сіңірілу сонымен қатар моноацилглицериндер, холестерин және май қышқылдарымен бірге гидролиз өнімдерінің шырышты ішектің клеткаларына өтуін қамтамасыз ететін ерігіш комплекс - аралас мицеллалар түзетін өт қышқылдарының қатысуымен жүреді. Өт қышқылы қан арқылы бауырға жеткізіледі, кейін өт қайта ішекке секреттеледі, яғни қайта пайдаланылады, айналым жүреді: бауыр - ішек - бауыр. Бірақ, тәулік бойы өт қышқылының шамамен 0, 3 г сіңірілмейді, калмен шығарылады. Жоғалған өт қышқылы бауырда холестерин көмегімен синтзделеді. Өт түзілуінің немесе ішекке өттің түсуінің бұзылуы майлардың ыдырауы мен олардың кал құрамына бөләнуінің - стеаторреяның бұзылуына әкеледі.
Триацилглицериндердің ыдырау өнімдерінен ресинтезделуі шырышты ішек клеткаларында жүреді:
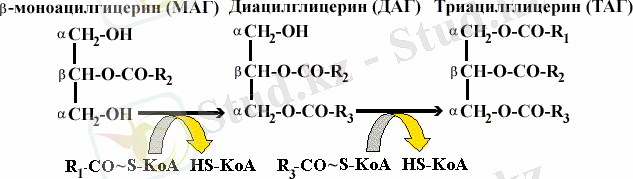
Ресинтезделген майдың лимфа жүйесі және қан айналымы арқылы тасымалдануы, оның липопротеиндердің құрамына енгеннен кейін мүмкін болады.
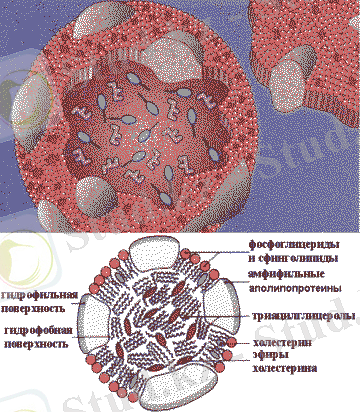
Липопротеиндердің құрылымы
Ішекте липопротеиндердің екі түрі түзіледі: хиломикрондар - ХМ және аз мөлшерде тығыздығы өте төмен липопротеиндер - ТӨТЛ. Хиломикрондардың құрамындағы экзогенді майлар органдар мен ұлпаларға жеткізіледі.
Липопротеинлипаза (ЛП-липаза) - экзогенді майларды ұлпаның рпайдалануын қамтамасыз ететін фермент. тамырлардың эндотелиінде ыдырайтын ЛП-липаза қан айналымының хиломикрондарымен әсерлесіп, триацилглицериндерден клеткаға түсетін глицерин және май қышқылдарын гидролиздейді. Хиломикрондардан ТАГ бөліп алғаннан кейін, хиломикрондар қалдық хиломикрондарға айналады да бауырға түседі. Майларды қоректену сипаты мен энергетикалық шығындарға байланысты тәулігіне 50-100 г. қажет етеді.
Майлардың депонирование және мобилизациясы
Майлар гликоген сияқты энергетикалық материалдың депонирленген формалары болып табылады. Сонымен қатар, майлар - ең ұзақ және эффективтілігі жоғары энергия көзі. Ашыққан кезде адамдағы май қоры 5-7 күнде бітеді, ал гликоген бір тәулікте толығымен пайдаланылады. Егер майлардың түсуі энергияның организмдегі қажеттілігінен жоғары болса, онда иай май ұлпаларының арнайы клеткаларында - адиипоциттерде депондирленеді. Сонымен қатар, егер организмдерге түсетін көміртегі мөлшері гликоген түрінде депондирования үшін шамадан тыс болса, онда глюкозаның бір бөлігі майға айналады. Осылайша, май ұлпаларында май үш процесс нәтижесінде жинақталады:
- ішектен экзогенді майларды әкелетін хиломикрондардан түседі
- бауырда глюкозадан синтезделіп, эндогенді майларға тасымалданатын ТӨТЛ түседі
- май ұлпаларының өз клеткаларында глюкозадан түзіледі
Бірінші және екінші жағдайда липопротеиндердің құрамындағы майлар ЛП - липаза көмегімен гидролизденеді және клеткаға кейін ТАГ синтезі үшін пайдаланылатын май қышқылдары түседі. Май қышқылдары ТАГ түсу үшін алдымен А коферменттерінің тиоэфирлерінің түзілу жолымен активтеледі, кейін глицеролфосфатпен әсерлеседі. Глицерол адипоциттерде фосфорлана алмайтындықтан (бұл клеткаларда глицеролкиназа болмайды), глицеролфосфат тоттықсыздануы кезінде түзіледі.
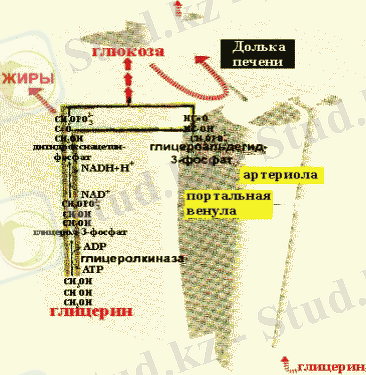
Бауырдағы глицерол метаболизмі
Сәйкесінше, ТАГ синтезі глюкоза бар жағдайда ғана жүзеге асады. Одан гликолиз процесінде диоксиацетонфосфат түзіледі. Инсулин ТАГ синтезін стимулдейді, өйткені ол бар ортада май ұлпалары клеткаларының мембраналарының глюкоза үшін өткізгіштігін жоғарлатады.
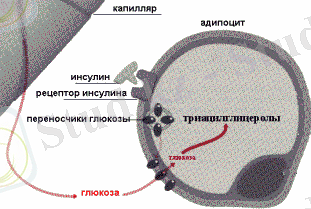
Майлардың депондированиясындағы инсулиннің ролі
Депонирленген ТАГ мобилизациясын (липолиз) ұлпалық липаза катализдейді. Нәтижесінде майлар глицерин мен бос май қышқылдарына ыдырайды.
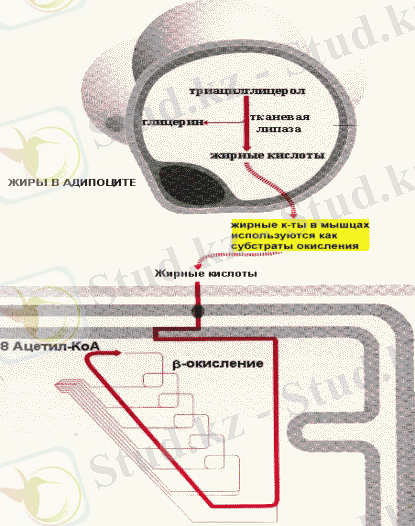
Майлардың мобилизациясы және оның бұлшықеттердегі тотығуы
Адреналин мен глюкагон клеткаішілік липазаны активтендіреді. Бұл гормондардың әсер етуі аденилатциклаза каскадының реакциясымен опосредовано. Ол осы кезде белсенді формаға өтіп, ТАГ эфилі байланыстарды ыдырататын аденилатциклазаның активтенуінен басталып, липазаның фосфорлануымен аяқталады. Глицерол плазмада еритін зат ретінде бауырға тасымалданады, ол онда глюконеогенез реакцияларына пайдаланылады. Май қышқылдары қан арқылы комплекс түрінде сарысу альбуминімен бірге әр түрлі органдар мен ұлпаларға тасымалданады, ол онда тотығу процесіне қатысады.
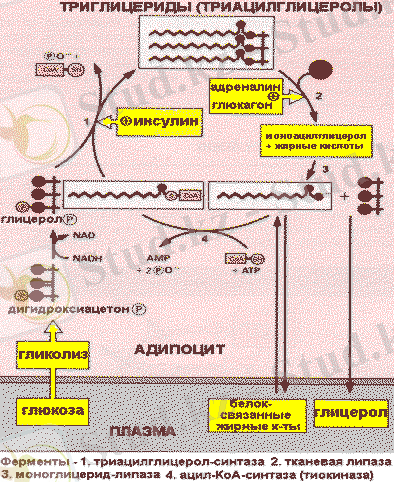
Триацилглицеридтердің синтезі мен мобилизациясы
Май қышқылдарының тотығуы
Май қышқылдарының тотығуы екі сатыдан тұрады:
- β -тотығу
- цитратты айналым
Бұл екі саты да тыныс алу тізбегімен жанасқан. β - тотығу клетканың митохондрияларында жүреді, ал май қышқылы қан айналымынан цитозольға түседі, ол онда А коферментінің конденсациялану жолымен тиоэфир түзе отырып активтенеді.
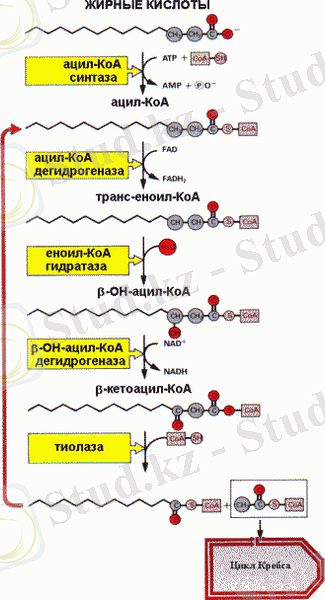
Май қышқылдарының тотығуы
Май қышқылы митохондриялық мембранадан өту үшін оларға арнайы тасымалдағыш - карнитин қажет. Сондықтан да, ацильді топ бірінші ацил-СоА-дан карнитинге тасымалданады. Ацил-карнитин мембранадан өтеді де, өзінің ацильді фрагментін А коферментіне қайта береді. Бұл қайтымды реакцияның ферменті ретінде ацил-карнитинтрансфераза қызмет атқарады. β - тотығудың келесі төрт реакцияның нәтижесі екі көміртекті фрагменттің ажырауы мен оның ацетил - СоА түзілуімен А коферментіне тасымалдануы. Ол кейін толығымен тотығу үшін цитрат айналымына енуі мүмкін. Қысқарған ацильді тізбек β - тотығу айналымына екінші рет енеді. β - тотығу айналымының қайталанатын итогы барлық ацил тізбегінің ацил - СоА дейін тотығуы болады.
Май қышқылдарының тотығу энергетикасы
β - тотығудың бір айналымы ішінде ацетил-СоА бір молекуласы түзіледі, оның цитрат айналымында тотығуы 12 моль АТР түзілуін қамтамасыз етеді. Сонымен қатар, 1 моль FADH 2 және 1 моль NADH түзіледі, олардың тыныс алу тізбегінде тотығуы кезінде сәйкесінше 2 және 3 моль АТР синтезделеді.
Осылайша, мысалы, пальмитин қышқылының (С16) тотығуы кезінде β-тотығудың 7 айналымы жүреді, нәтижесінде 8 моль ацетил-СоА, 7 моль FADH 2 және 7 моль NADH + түзіледі. Сәйкесінше, β-тотығу нәтижесіндегі АТР шығымы 35 АТР құрайды және цитратты айналым нәтижесінде 96 АТР түзіледі, барлығы 131 моль АТР.
Бауырдағы кетонды денелердің түзілуі (кетогенез)
Бауырдағы май қышқылдарының β-тотығуы бауыр клеткаларының энергетикалық қажеттілігіне пайдаланылатын тотықсыздандыру коферменттерінің түзілуіне әкеледі.
Бауырдағы β - тотығу нәтижесінде түзілген ацетил-СоА-ның кейінгі негізгі пайдаланылатын жолы - кетон денелерінің синтезі. Ол келесі ретпен жүреді: екікөміртекті молекула бір бірімен кейінгі реакцияларда ацетоацетат және β -гидроксибутират түзілуімен конденсацияланады.
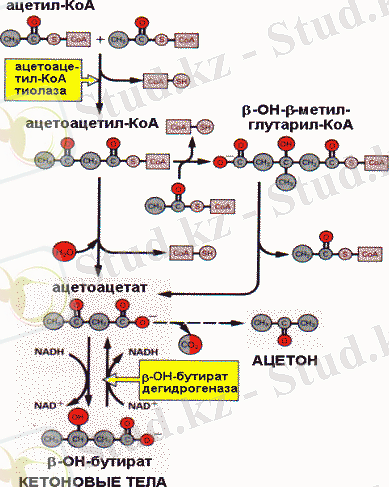
Кетон денелерінің синтезі
Бұл екі қышқыл кетонды денелер деп аталады. Кетонды денелер қанға диффундирлейді және бауыр ұлпаларынан басқаларымен энергия көзі ретінде пайдаланылады. Кетон денелерінің қандағы қалыпты концентрациясы 2 мг/л. Кетонденелерінің синтезін катализдейтін ферменттер митохондрияда орналасқан. Бауырда май қышқылдарының интенсивті тотығуы арнайы метаболитикалық жағдайларда жүрген кезде, біршама мөлшерде кетон денелері түзіледі. Бұл көмірсуарды пайдалану қиын жағдайда, анығырақ айтқанда организм үшін энергия көзі май қышқылдары (бұлшықеттің ұзақ жұмыс істеуі, ашығу, қант диабеті) болған кезде жүреді. Кетон денелерінің синтезделу жылдамдығы оларды ұлпаның пайдалануынан жоғары болып, қорға жинақталуына әкелуі мүмкін. Кетон денелерінің концентрациясының біршама жоғарылауы кетоацидоз (кетоз) деп аталатын қауіпті жағдай тудырады. Патологиялық кетоацитоз кезінде ацетоацетат ферментсіз декарбоксилденеді, ол ацетонның түзілуімен жүреді, сондықтан кетоз үшін ацетонның иісі тән.
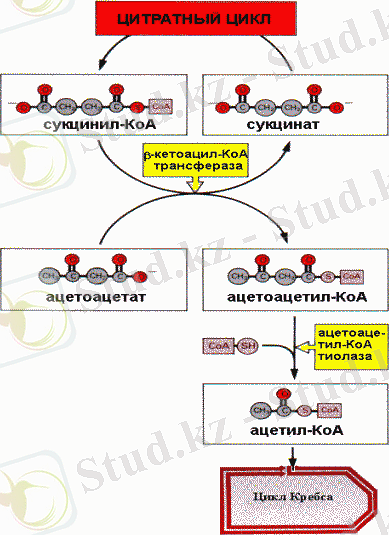
Кетон денелерінің тотығуы
Май қышқылдарының биосинтезі
Процесс клетканың әр түрлі бөліктерінде өтеді. Цитоплазмада С16 дейінгі (пальмитин) көміртегі тізбегі бар қаныққан май қышқылдары синтезделеді.
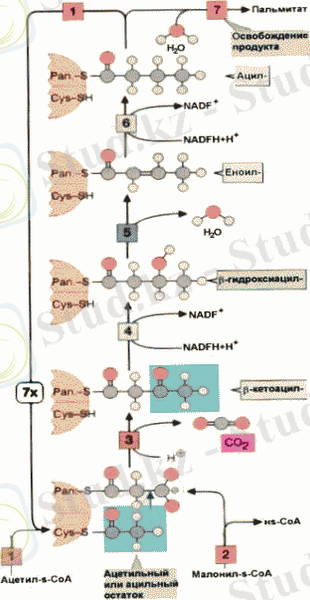
Жоғары май қышқылдарының синтезі
Митохондрияда тізбектің кейінгі ұзаруы жүреді, ал ритикулумда қаныққан май қышқылдары қанықпаған май қышқылдарына айналады, снымен қатар, тізбектің ұзарады.
... жалғасыСіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz