Қапшағай тоғандарының Lymnaeidae, Planorbidae және Physidae моллюскаларын лабораториялық жағдайда өсіру және олардың морфо-биологиялық сипаттамалары


РЕФЕРАТ
Диплом жұмысы 60 бетте жазылған, зерттеу нәтижелері 8 кесте және 17 сурет арқылы берілген. Қолданылған әдебиеттер тізімі 50.
Кілттік сөздер: құрсақаяқты моллюскалар, гермафродиттік без, даму, көбею, синкапсула, жұмыртқа, бақалшақ.
Диплом жұмысының мақсаты: Қапшағай уылдырық шашу және шабақ өсіру шаруашылығының тоғандарында кең таралған моллюскалардың Lymnaeidae, Planorbidae және Physidae тұқымдастарының өкілдерін лабораториялық жағдайда өсіру.
МАЗМҰНЫ
Б.
1. 1. 2
1. 1. 3
1. 1. 4
1. 1. 5
1. 1. 6
1. 1. 7
1. 1. 8
1. 2
1. 3
Құрсақаяқты моллюскалардың бұлшықеттеріне сипаттама . . .
Құрсақаяқты моллюскалардың асқорыту жүйесі . . .
Құрсақаяқты моллюскалардың тыныс алу жүйесі . . .
Құрсақаяқты моллюскалардың қанайналу жүйесі . . .
Құрсақаяқты моллюскалардың зәр шығару жүйесі . . .
Құрсақаяқты моллюскалардың жүйкежүйесі . . .
Құрсақаяқты моллюскалардың көбею жүйесі . . .
Құрсақаяқты моллюскалардың экологиясы . . .
Моллюскалардың маңызы . . .
11
11
15
17
18
18
20
23
27
2. 1
2. 2
2. 3
Зерттеу материалдары . . .
Құрсақаяқты моллюскаларды жинау әдістері . . .
Құрсақаяқты моллюскаларды өңдеу әдістері . . .
30
31
32
3. 1
3. 2
3. 3
Кең таралған моллюскалардың Lymnaeidae, Planorbidae және Physidae тұқымдастарының өкілдерінің табиғи жағдайда өсу ерекшеліктері . . .
Кең таралған моллюскалардың Lymnaeidae, Planorbidae және Physidae тұқымдастарының өкілдерінің тіршілік циклдерінің ерекшеліктері . . .
Вегетационды кезеңде популяция құрамындағы моллюскалардың жастық өлшем көрсеткіштері . . .
34
37
44
3. 4
3. 5
Құрсақаяқты моллюскаларды лабораториялық жағдайда өсіру ерекшеліктері . . .
Лабораториялық жағдайда өсірілген Planorbidae тұқымдасының өкілі Planorbarius corneus - тың морфологиялық ерекшеліктері . . .
45
47
КІРІСПЕ
Құрсақаяқты моллюскалар табиғатта өте кең таралған жануарлар тобы. Теңізде, тұщы суларда және құрлықта мекендей отырып, табиғи биоценоздардың маңызды компоненті болып табылады. Құрсақаяқты моллюскалар, тіпті, өздері өсіп-өндіру объектісі ретінде қолданылуы мүмкін, алайда ауылшарушылық зиянкестері ретінде де белгілі немесе кейбір ауланатын құнды балықтардың қорегі бола тұра үй жануарлары мен адамға қауіпті құрттардың тасымалдаушысы болуы да мүмкін. Сонымен қатар моллюска бақалшақтары биостратиграфиялық жер қойнауын бөлшектеуде маңызды рөл атқарады. Ал бұл қазба байлықтардың көзін табуда негізгі элементтердің бірі.
Біздің елімізде кең таралған және де әр түрлі типті су қоймаларда кездесетін, көптеген тұщы су бассейндері экосистемасының міндетті компоненті - өкпелі моллюскалар болып табылады. Оларды меңгеру практикалық және ғылыми қызығушылықты тудырады.
Жануарлардың бұл тобы өздерінің популяциясының тығыздылығымен, түрлік құрамының алуантүрлілігімен, көптеген организмдермен арақатынаста болуымен ерекшеленеді. Осындай ерекшеліктеріне қарай көптеген экосистеманың құрам бөлігіне жатқызады.
Суқоймаларда құрсақаяқты моллюскалардың биомассасы барлық су түбілік жануарлар биомассасының 70-80%-ын құрайды. Көпшілік құрсақаяқты моллюскалар тұрақты компонент ретінде көптеген балық түрлерінің қоректік рационының құрамына кіреді. Моллюскалар кейбір балықтардың, құстардың және сүтқоректілердің қоректік рационының негізгі компоненті болып табылады [1, 2, 3] .
Моллюскалардың практикалық маңызы зор. Адамдарға шаруашылық жағынан моллюскалардың пайдалы және зиянды жақтарын ескеріп отыру қажет. Моллюскалардың әртүрлі типті суқоймалардағы өнімділік процессі олардың дамуы мен көбеюіне, және де экологиялық ерекшеліктеріне негізделу керек. Алайда, моллюскалардың биоөнімділігіне және Қазақстан тұщы су бассейндеріндегі энергия мен жалпы зат айналымындағы рөліне арналған зерттеулер толық емес.
Диплом жұмысының мақсаты:
Қапшағай уылдырық шашу және шабақ өсіру шаруашылығының тоғандарында кең таралған моллюскалардың Lymnaeidae, Planorbidae және Physidae тұқымдастарының өкілдерін лабораториялық жағдайда өсіру.
Диплом жұмысының міндеттері:
- кең таралған , өкілдерінің табиғи жағдайда өсу ерекшеліктерін анықтау;
- кең таралған , өкілдерінің тіршілік циклдерінің ерекшеліктерін анықтау;
- вегетационды кезеңде популяция құрамындағы зерттелінген моллюскалардың жастық өлшем көрсеткіштерін анықтау;
- құрсақаяқты моллюскаларды лабораториялық жағдайда өсіру ерекшеліктерін анықтау;
- лабораториялық жағдайда Planorbidae тұқымдасының өкілін Planorbarius corneus - ты өсіру және морфологиялық ерекшеліктерін анықтау [1, 3] .
НЕГІЗГІ БӨЛІМ
- ӘДЕБИЕТТЕРГЕ ШОЛУ
Моллюскалар типі түрлерінің саны жағынан буынаяқтылар типінен кейін екінші тұрған кең таралған жануарлар тобы. Түрлі мекен орталарын игеріп, омыртқасыз жануарлардың бұл тобы популяцияларының тығыздылығымен белгілі. Көптеген тірі организмдермен қатынаста, байланыста болуының нәтижесінде күрделі биологиялық тізбектердің құрамына енеді. Сондықтан да олардың көптеген экожүйелердің құрамдас бөлігі ретінде маңызы зор.
Моллюскалар немесе былқылдақ денелілер, олигомерлы көпқылтанды буылтық құрттардан тараған, өз алдына жеке оқшауланған тип. 130 000-нан аса түрі бар. Мұхиттарда, теңіздерде, тұщы су қоймаларында және құрлықта тіршілік етеді.
Моллюскалардың денесі кутикула қабатымен қапталынған, ал оның сыртын тек моллюскаларға тән, бір қабатты эпителийден құралған, мантия деп аталатын үлкен тері қатпаршағы жауып тұрады. Мантия мен дененің аралығында мантия қуысы пайда болып, онда желбезектері, кейбір сезім мүшелері (осфрадиялар) орналасады және бүйрек пен жыныс бездерінің, артқы ішектің тесіктері ашылады. Осы құрылымдар, жүрек пен бүйректерді қоса мантия жиынтығы немесе мантия комплексі деп аталады.
Моллюскалардың тағы бір ерекшелігі - бақалшақтың болуы. Ол мантия жабынында орналасқан көптеген түрлі бездердің шығарған секреттерінен түзіледі. Дененің сыртқы жағында орналасып, қорғаныш қызметін атқарады. Бақалшақ үш қабаттан құралады: органикалық заттан тұратын сыртқы қабат немесе конхиолин қабаты; ізбесті - ортаңғы қалың қабат және жұқа, жылтыр түсті - ішкі інжу немесе перламурт қабаты. Мантия жабыны бақалшақты астарлап жатады [2, 3] .
Басқа омыртқасыздардан ерекшеліктері - ас қорыту жүйесіндегі, жұтқыншақ бөлімінде, қоректі ұсақтайтын ерекше аппарат - радула деп аталатын үккіштің болуы тән [4] .
- Құрсақаяқты моллюскалардың сыртқы және ішкі құрылысы
Құрсақаяқтылардың немесе ұлулардың 90 000-нан астам түрі бар, көпшілігі теңіздерде, сондай-ақ тұщы сулар мен құрлықта және паразиттік тіршілік ететін түрлері де бар.
Құрсақаяқтылардың дене тұрқы 2 - 3 мм-ден бірнеше сантимертге дейін барады. Ең ірісі Hemifusus proboscidiferus , оның ұзындығы бақалшағымен қоса 60 см. Сол сияқты теңіз Aplysia -ның ұзындығы 25 см, Африкада мекендейтін құрлық ұлулары Achatina да ірі ұлулар қатарына жатады.
Құрсақаяқтылардың денесі үш бөлімге бөлінген: бас, аяқ, тұлға. Басы тұлға бөлімінен айқын бөлініп тұрады. Жақсы жетілген аяғында кең жорғалағыш табаны болады. Тұлғасы дененің жоғарғы жағына қарай қап тәрізді өсіндіні түзеді, бұны ішкі қапшық деп те атайды. Барлық құрсақаяқтыларға тән ерекшелік, ол құрылысының асимметриялы болуы.
Бұлардың бақалшағы жақсы дамыған, кейбір кезде редукцияға да ұшырауы мүмкін.
Құрсақаяқты моллюскалардың асимметриясы келесі белгілермен сипатталады: оң жақ мантия комплексінің мүшелері жойылып сол жағы мүшелерінің күшті дамуы; ішкі қапшықтың немесе тұлғаның спираль тәрізді оралуы, бұл ерекшеліктері бақалшақтың спираль тәрізді оралуына байланысты.
Тұщы және тұзды суларда мекендейтін құрсақаяқты моллюскалардың денесі бас, аяқ және құрсақ бөлімдерінен тұрады, асимметриялы, бақалшағы спиральді ширатылған және бунақталмаған болып келеді. Денесі сыртқа қарай ізбесті бақалшақ бөлетін мантиямен қапталған. Жалпы моллюскалар бір-біріне мүлдем ұқсамайтын құрсақаяқты, қосжақтаулы, басаяқты және т. б. моллюскалардың ортақ құрылымы мүшелердің мантиялық кешенінің (комплекс) болуымен анықталады. Дене мен мантияның ішкі бетінің арасында желбезек, асқорыту, зәр шығару және жыныс мүшелерінің тесіктері ашылатын мантия қуысы орналасқан. Атмосфералық ауамен тыныс алатын моллюскалардың желбезектік тыныс алуы өкпелік тыныс алумен алмастырылады. Оларда өкпенің функциясын өкпелік қуыс деп аталатын түрөзгерген мантия қуысының бөлігі атқарады.
Өкпемен тыныс алатын моллюскалардың көбісінде аналь, жыныс тесіктері, өкпе қуысы дененің сол жағына ашылады. Мұндай жағдайда моллюскалардың денесі асимметрия құрылысты немесе лейотропты деп атайды, ал жоғарыдан қарағанда бақалшақ сағат тіліне қарсы бұралады. Оң жаққа бұралған жағдайда бақалшақты гиперстрофты деп атайды. Lymnaea және Acroloxus туыстарына жататын моллюскалар денесіне оң симметрия тән. Кейде жұмыртқада қалыптасып келе жатқан бақалшақтың (эмбриональді бақалшақ) бір бөлігі басқа иірімдерге қарама-қарсы бұралады. Бұл кезде эмбриональді бақалшақтың гетерострофтылығы байқалады [1, 3, 4] .
Құрсақаяқтылардың дене пішіні алуан түрлі, көпшілігінің денесі ұзынша және арқа жағы томпайып шығыңқы болып келеді. Жақсы жетілген басының бауыр жағында аузы, арқа жағында 1-2 жұп қармалауышы мен жұп көздері орналасқан. Басының алдыңғы бөлігі кейде ұзын тұмсықша ретінде созылып тұрады (1-сурет) .
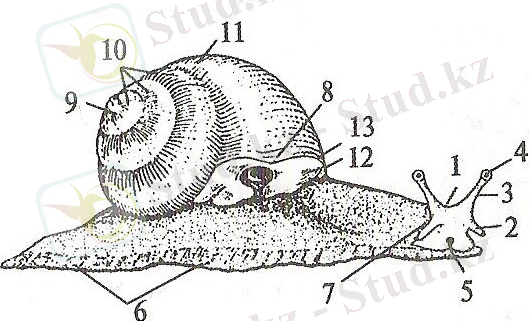
1 сурет - Құрсақаяқты моллюсканың сыртқы құрылысы: 1 - басы, 2 - ауыз қармалауышы, 3 - көз қармалауышы, 4 - көзі, 5 - ауыз тесігі, 6 - аяқ, 7 - жыныс тесігі, 8 - тыныс алу тесігі, 9 - бақалшақтың төбесі, 10 - бақалшақтың спиральді айналымы, 11 - соңғы айналым, 12 - саға, 13 - сағаның шеті
Аяғы бұлшықетті және жорғалаушы табаны бар, ол дененің бауыр бөлімінің өсіндісі. Сондықтан да бұл ұлуларды құрсақаяқтылар деп те атайды. Аяғының жиырылу нәтижесінде жануар субстрат бетінде баяу қозғалып жүреді. Әр алуан тіршілік етуіне байланысты аяғы да алуан түрлі модификацияға ұшыраған. Ең күшті өзгеріске ұшыраған жүзіп тіршілік ететін Құрсақаяқты класының кейбір өкілдерінің аяқтары. Мысалы, қырлыаяқтылардың аяғы бүйірлерінен қабысып жіңішке вертикальді жүзгіш қанатқа айналған, кейде бүйір жиектерінде екі жағына өсетін табанының рудименті байқалады. Аяқтың артқы бөлігі ұзын құйрық тәрізді тұтқаға созылады. Ал, қанатаяқтылардың аяғының бүйірлік бөлігі күшті өсіп кетеді де, қанат тәрізді қалақша түзеді [2, 4] .
Тұлғасы немесе ішкі қапшығы, төменгі сатыдағы құрсақаяқтыларда симметриялы және , өкпелілерге тән аяқ бөліміне айқын бөлінбеген. Ал, Құрсақаяқтылар класының көпшілік түрлерінде керісінше, тұлғасы аяғының үстінде спиральді оралған қапшық түрінде бөлінген. Тұлғаның төменгі жерінде жабын қатпарлары немесе мантия қатпары, ал оның астында мантия комплексінің мүшелері бар мантия қуысы түзіледі. Қатпар да, мантия қуысы да тұлғаның алдыңғы және оң жағында дамыған. Мантия бақалшақты бөліп шығарады, көбінесе спиральді оралған бақалшақты түзейді (2 сурет) .
Бақалшақтың төбе жағы тұйық біткен - ол оның төбесі деп аталады, ол оган қарама-қарсы екінші ұшында саңылауы орналасқан. Саңылауы арқылы жануардың басы мен аяғы сыртқа шығып тұрады. Тек сирек жағдайда ғана бақалшақтарының иірімдері бір жазықтықта жатады.
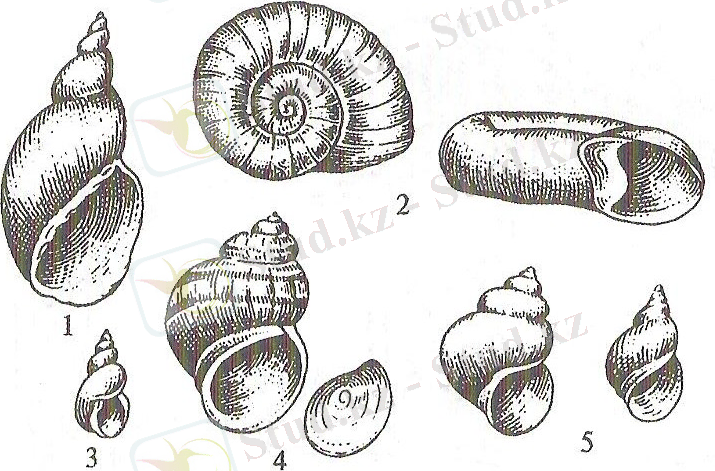
2 сурет - Тұщы суда тіршілік ететін құрсақаяқты моллюскалардың бақалшақтары: 1 - кәдімгі тоспа ұлуы, 2 - катушка, 3 - кіші тоспа ұлуы, 4 - тірі туатын шалғын ұлуы (лужанка), 5 - битиния
Бақалшақтың спираль иірімдері конус тәрізді (турбоспиральді) . Иірімдерінің диаметрі жануарлардың өсуіне сәйкес төбесінен қарама-қарсы саңылауына дейін үлкейеді. Әрбір жаңа түзілген ірілеу иірімдері алдыңғыларды орап орналасады. Мұндай бақалшақтар инволютты деп аталады. Көбінесе соңғы орамдар алғашқысына жақын тұратындықтан олардың барлығы көрініп тұрады. Құрсақаяқтылардың арасында бақалшақтың спиральді оралуының бағыты бойынша бақалшағы екі типті болады: үстінен қараған спираль, сағат тілінің бағытымен бұралса, онда мұндай бақалшақты оңға бұралған немесе дексиотропты деп атайды. Мұндай бақалшақ көпшілігінне тән. Егер, бақалшақ солға қарай бұралса, онда оны солға қарай бұралған немесе лейотропты бақалшақ деп атайды. Осьтің айналасында спираль тәрізді бұрамдары болса оны тығыз бағана деп атайды. Ал, иірімдері ішкі қабырғалары арқылы бір-біріне жанаспаса, ода бағана орнына қуыстар немесе кіндік түзіледі. Спиральді бақалшақтың кеңдігі сонша, ұлулар қауіпті жағдайда барлық денесін соған жинап алады.
Алдыңғы желбезектілер класс тармағының өкілдерінде аяғының артқы бөлігінің арқа жағында мүйізді немесе ізбесті қақпақшасы дамыған. Ол, моллюсканың денесі бақалшақтың ішіне кіргеннен кейін, бақалшақтың тесігін жауып тастайды.
Құрсақаяқтылардың кейбір түрлерінің бақалшағы біршама қарапайым құрылысты және қалпақ тәрізді болып келеді, себебі олардың дернәсілдерінің дамуында алдымен бұралған бақалшағы пайда болады да, кейінірек ол қалпақ пішінге көшеді.
Құрсақаяқтылардың бақалшағы сыртқы жұқа органикалық қабаттан тұрады, оның астыңғы бақалшақ бетіне перпендикуляр орналасқан, ізбесті тақташалардан түзілген фарфор тәрізді қабат жатады. Кейбір құрсақаяқтыларды, мысалы Haliotis, Turbo-да жылтыр перламурт қабаты бар.
Құрсақаяқтылардың барлық өкілдерінде бақалшақтың редукция байқалады. Редукция мынадай кезеңдер арқылы жүреді: бақалшағы жақсы жетілген, бірақ жануардың денесі жиырылғанда бақалшаққа толығымен орналаспайды; бақалшақ одан әрі кішірейе береді де екі мантиялы қатпарлармен көмкеріледі; мантия қатпарлары бір-бірімен бақалшақтың үстінде ортаңғы сызық бойымен бірігіп кетеді, соның нәтижесінде сыртқы бақалшақ ішкі нәзік жұқа тақтаға айналады; ішкі тақталы бақалшақ жойылып, онын орнын терідегі шашылып жатқан ұсақ ізбесті денешіктер басады. Мысалы, шырыш - Arion -да, жалағашжелбезектілер - Nudibranchia -да, ал бақалшақтың барлық ізі Pterotrachea ұлуларында мүлдем жойылған. Бақалшақтың жойылуы көбінесе жүзіп өмір сүретін, құрлықта және паразит құрсақаяқтыларда байқалады, себебі жүзу және жорғалау үшін бақалшақ кедергі жасайды, яғни олардың қажеті жоқ [2, 3, 5] .
- Құрсақаяқты моллюскалар бақалшағының ерекшеліктері
Тұщы және тұзды суларда мекендейтін моллюскалардың бақалшағы асимметриялы, спиральді ширатылған немесе қалпақша тәрізді болып табылады. Спиральді ширатылған бақалшақтың иірімдері бір жазықтықта (жалпақ спиральді) немесе бірнеше жазықтықтарда (турбоспиральді) болады, сонымен қоса иірімдердің ұлғаюына қарай бақалшақтардың пішіні де әртүрлі болуы мүмкін. Бақалшақ, әдетте, сары, мүйізді сары, қоңыз, қызыл-қоңыр, қара немесе жасыл және т. б. түсті болып келеді. Кейбір түрлерде бақалшақта әртүрлі жолақтар, дақтар, нүктелер немесе т. б. пішіндер кездеседі [1, 3, 5] .
Бақалшақтың ең үстіңгі нүктесі оның шыңы, ал моллюскалардың жұмсақ денесі жиналатын қуыс саңылау деп аталады. Шыңы жоғары қарай қарап тұрған бақалшақтың саңылауы ширатылу осінің қай жағында болуына байланысты, оңға немесе солға бұратылған бақалшақ деп бөлінеді. Саңылауға қарама-қарсы соңғы иірімнің беті желке деп аталады. Бақалшақтың пішініне қарай соңғы иірім ұзындығы әртүрлі болады.
Бақалшақтың сыртқы беті тегіс, жылтыр немесе әрең байқалатын сызықтар және терең емес ішке қарай майысқан шұңқырлары болады. Кейде иірімдерде сүйір бұрыш (киль) байқалады және ол соңғы иірімнің ортаңғы, астыңғы немесе үстіңгі бөлімдердің бойымен өтеді, сиректеу барлық иірімдерде болады. Турбоспиральді бақалшақтың ішкі қабырғалары бір-бірімен бірігіп кетіп кішкентай каналы бар бағана құрайды. Бұл каналдың бақалшақтың астыңғы жағына ашылатын жерін кіндік деп атайды. Ол үлкен, тар, саңылау тәрізді болып келеді немесе толығымен өсіп кеткен саңылау жиегімен жабылып тұрады. Кейде бақалшақтың астыңғы бетінде сифональді өсінді болады. Осы сифональді өсінді судың мантия қуысына кіруімен қамтамасыз етеді [5] .
Бақалшақ саңылауы домалақ, сопақша, алмұрттәрізді, құлақтәрізді және т. б. болуы мүмкін. Саңылаудың бос шеті, бағанаға қатысты шеті, бағанаға қатысты беті, сонымен қатар соңғы иірім қабырғасынан пайда болған бөліктер палатальді, колумеллярлы және париентальді деп аталады. Кейде палатальді бөліктің төменгі жағын базальді деп те атайды. Ал моллюскалардың кейбір түрлерінде колумеллярлы және париентальді бөліктер ұлғайып, колумеллярлы алаң құрайды. Кейде саңылау шеттерінде қалыңдаған қатпарлар пайда болады. Бақалшақ қабырғалары жұқа, сынғыш, қалыңдығы орташа немесе өте қалың және мықты болуы мүмкін [1, 3, 6] .
- Құрсақаяқты моллюскалар бұлшықеттеріне сипаттама
Құрсақаяқты моллюскалардың бұлшықеттері колумеллярлы бұлшықеттен тұрады. Моллюскалардың денесінің бақалшақ ішіне жиырылуы осы бұлшықет көмегімен жүзеге асады. Бұлшықеттің жоғарғы ұшы бақалшақ бағанасына бекітіледі, сол жерден аяққа қарай бағытталады да көптеген бөлек талшықтарға шашырайды. Дене жиырылған кезде аяқтың табаны бір-біріне сәйкес екіге бүгіледі.
Аяқ бұлшықеті қимыл-қозғалыс функциясын атқарады. Құрсақаяқтылардың кимыл-қозғалысы негізінен белгілі бір заттың үсті немесе су қабығының тартылу бетімен сырғуы болып табылады. Сырғу табан бетіндегі бұлшықеттің толқын тәрізді жиырылу нәтижесінде жүзеге асады, сонымен қоса аяқтың эпителийі және педальді бездер шырыш бөліп тұрады.
Бір немесе екі ретрактордан тұратын, жұтқыншақтың жиырылуын қамтамасыз ететін бұлшықеттер дистальді ұшында колумеллярлы бұлшықетпен қосылады [1, 6] .
Жыныс мүшелерінің бұлшық еттері бұл аталық шағылысу мүшесінің ретракторы және алдыңғы мен артқы протракторлар болып табылады. Аналық жыныс мүшелерінде арнайы бұлшықеттер жоқ.
Жоғарыда айтылған бұлшық еттерден басқа тері бұлшықеттері және әртүрлі мүшелер қабырғаларындағы бұлшықеттер бар [1, 3, 7] .
- Құрсақаяқты моллюскалардың асқорыту жүйесі
Құрсақаяқтылар басының алдыңғы ұшында ауыз тесігі орналасқан, ол ауыз қуысына ашылады да әрі қарай жұтқыншаққа жалғасады. Ауыз қуысы мен жұтқыншақ шекарасында кейде жақсы дамыған жақтар орналасады. Жұтқыншақ алдыңғы ілгектің кеңейген, бұлшықетті бөлігі болып табылады. Оның вентральді бөлігінде бұлшықетті тіл - одонтофор орналасады. Одонтофордың алдыңғы ұшы жұтқыншақ қуысына кіріп тұрады [2, 4, 7] .
Жұтқыншақ қуысын астарлап жататын жабынды эпителийде бөлек кешендерге топтасқан кутикулалы, кірпікшелі және шырышты клеткалар байқалады [4, 8] .
Тіл кутикулярлы қабықшамен қапталған және үстінде көлденеңінен орналасқан тіс қатарлары - радула бар. Радула - қоректі ұсақтайтын, радулярлы тістердің пластинкалары орналасқан склеропротеинді таспа, артқы ұшында үнемі қалыптасып, алдыңғы ұшында тозып, жойылып тұрады. Радулярлы тіс - радулярлы мембранадағы кутикулярлы тақташа, базальді тақташа, дене және коронкадан құралған.
Радулярлы аппарат - радула мен одонтофордан тұратын мүшесі моллюскалардың қоректі табу және жинақтау оны қорыту болып табылады [8] .
Өкпемен тыныс алатын моллюскаларда радуланың көлденең қатары бір-біріне бірте-бірте ауысатын орталық тістен тұрады. Осы орталық тістің екі жағынан латеральді тістер орналасады, ал одан әрі қарай аралық және маргинальді немесе шеткі тістер орын алады. Әрбір тісте оның негізі және бір немесе бірнеше тісшелері бар кутикулярлы бөлігі болады [1, 3, 6, 8] .
Радула тістерінің химиялық құрамы моллюсканың жасына қарай өзгеріп отырады. Алғашқы кезеңінде тістер белоктан және хитиннен (мүмкін, насекомдардың хитиніне ұқсас гликопротеиннен) құралады. Кейіннен амин топтарының және күміске реакция беретін материалдардың пайда болатыны анықталды. Соңында тіс денесінде темір тұздары мен кремний жинақталады [4, 9] .
Жұтқыншақтың артқы бөлігіне қалақшалы немесе бұтақталған бір, кейде екі жұп сілекей бездерінің түтікшелері ашылады. Әрі қарай жұтқыншақ өңешке жалғасады. Өңештің өзі гистологиялық құрылысы жағынан алдыңғы, ортаңғы және артқы бөлімдерге дифференциацияланған. Бұл бөлімдердің жабын эпителийі негізінен қылаулатқыш призмалық клеткалардан құралған. Өңештің ортаңғы бөліміндегі клеткалар ерекше жоғары сектеторлы қасиетке ие. Кейбір моллюскаларда өңеш әрі қарай артқы бөлігінде кеңейіп, қарынның алдыңғы бөлігін құрайды. Қарынның басқа бөлігі ортаңғы ішектің кеңейген қалта тәрізді құрылымы болып табылады. Қарынның осы бөлігіне асқорыту безі - бауырдың өзегі ашылады. Лимнеидтерде қарынның бұл бөлігі қалақшалы бұлшықеттік және созылыңқы пилоритикалық бөлімдерден тұрады [1, 4, 5, 9] .
Құрсақаяқтылардың бауыры көлемді, қоңыр түсті, әдетте, жұп емес немесе егер жұп болса, симметриясыз, қарынмен бір-екі текіспен байланысқан. Сонымен қатар, бауыр бұтақталған түтікшелердің күрделі жүйесі мен екі негізгі клеткалар типінен: секреторлы мен абсорбты заттары бар асқорыту клеткалардын және экскреторлы заттарды қоңыр түсті конкрециялар түрінде жинақтайтын бөліп шығарғыш клеткалардан құралады. Қарастырылып отырған асқорыту безі - бауыр сіңіру, асқорыту және заттарды бөлу қызметтерін атқарады [1, 3, 9, 10] .
Алдыңғы, ортаңғы және артқы ішек бөлімдерінде өздеріне тән қатпарлануы және секреторлы, кірпікшелі мен сіңіру қызметін атқаратын клеткалар ішектің барлық бөлімдерінде кездеседі, тіпті тік ішекке дейін. Осыған орай сіңіру процессі ішектің барлық бөлімдерінде жүреді [4, 9, 10] .
Аузы басының алдыңғы төменгі жағында орналасқан, кейбір түрлерінде аузы ұзарып тұмсық түзеді. Ал, жыртқыш түрлерінде басының алдыңғы ұшы одан да бетер ұзарып, жиырылатын және қорегін ұстауда атыла алатын бұлшықетті тұмсықшаға айналған.
... жалғасыСіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz