Жоғары сатылы өсімдіктердің морфогенезіне цитокинин медиаторының әсері

әл-Фараби атындағы Қазақ ұлттық университеті
БИОЛОГИЯ ФАКУЛЬТЕТІ
Генетика және молекулалық биология кафедрасы
БІТІРУ ЖҰМЫСЫ
Цитокини медиоторының жоғары сатыдағы өсімдіктер морфогенезіне әсерін зерттеу
Орындаған: Тоқтасын Нүргул.
(қолы, күні)
Ғылыми жетекші:
ҚР ҰҒА - ның
академигі Гильманов М. Қ.
(қолы, күні)
Норма бақылаушы Сатылған И. А.
(қолы Күні)
Қорғауға жіберілді:
Кафедра меңгерушісі,
б. ғ. д., профессор Айташева З. Г.
(қолы, күні)
Алматы, 2008
РЕФЕРАТ
Бітіру жұмысы: 34 беттен, 38 әдебиеттен, 1- сызбадан, 9 -суреттен тұрады. Бітіру жұмысын Тоқтасын Нұргүл М. А. Айтхожин атындағы биохимия және молекулалық биология институтының ферменттердің құрылымы мен реттелу лабораториясында өткізді.
Өзектілігі: Цитокинин медиаторын бөліп алып, тазартып, медиаторының морфогенезге әсерін қарастыру
Неізгі сөздер: бидай дәні, ұрық бөлігі, алейрон қабаты, НАДФ глютаматдегидрогеназа, цитокинин, цитокинин медиаторы, ризогенез, морфогенез, каллус
Алдына қойылған мақсаты: цитокинин медиаторын бөліп алып, тазартып оның морфогенезге әсерін қарастыру.
Қолданылған әдістер: Октил сефароза (CL-4B) гидрофобты хроматографиясы және RP-18 колонкада жасалынатын жоғары қысымды кері фазалы хроматография, гомогенизациялау, центрифугалау, инекьция жасау, , Мурасиге - Скуг ортасында каллустарды өсіру
ҚЫСҚАРТЫЛҒАН СӨЗДЕР
ДАГ - диацилглицерол
PIP - фосфоинотидилинозитол
ИФ 3 - инозитол үш фосфат
ФИФ 2 - фосфоинотидилинозитол 2 фосфат
НАДФ-ГДГ- глютаматдегидрогеназа
РНҚ - рибонуклеин қышқылы
ЦББ - цитокинин байланыстырушы белоктар
ИУК - индонил сірке қышқылы
БАП - бензоаминопурин
ФК
-фузикокцин
МАЗМҰНЫ
Кіріспе. . . . 5
1. Әдебиеттерге шолу . . . 6
1. 1Цитокининнің ашылуы мен химиялық табиғаты . . . 6
1. 2. Фитогормондардың атқаратын ролі . . . 8
1. 3. Цитокинин медиаторының өсімдік морфогенезіне әсері . . . 11
1. 4. Цитокининнің медиаторы . . . 16
1. 5. Цитокинин синтездеуші гендердің трансфромациясы . . . 16
2. Тәжірибелік бөлім . . . 18
2. 1. Материалдар мен зерттеу әдістері . . . 18
2. 2. Алынған нәтижелерге талдау жасау. 19
Қорытынды . . . 30
Пайдаланылған әдебиеттер . . . 32
КІРІСПЕ
Қазіргі заманғы биологияның ең болашағы зор әдістерінің бірі өсімдік клеткасы мен ұлпасын жасанды жағдайда өсіру. Әсіресе гаплоидты технологияның тиімділігі зор. Гаплоидты организмде барлық рецессивті белгілері көрінеді. Аталық тозаңдарды өсіру арқылы гаплоидты организмдерді алуға болады. In vitro жағдайында өсімдік ұлпасын өсіріп, өсімдік жүйесінің бір клеткадан дифференциация гистогенез, морфогенез арқылы біртұтас өсімдік алу мүмкіндігінің күрделі механизмдерін зерттеуге болады. Клеткаларды In vitro жағдайында өсіру әдістерінің ең күрделі мәселесі каллус клеткаларының морфогенез процестерін реттеу болып табылады. өсімдік клеткасын жасанды жағдайда өсіргендегі технологияда қолданудың басты мақсаты біртұтас регенерант өсімдігін алу. Даражарнақты өсімдіктердің, әсіресе астық тұқымдастардың оқшауланған ұлпаларын In vitro жағдайында өсіруге өте қиын, себебі бұл өсімдіктердің тотипатенттілік қасиеті төмен. Сондықтан органогенез процесіне эффективті әсер ететін және каллус түзуге, өсу және морфогенез процестерін реттейтін жаңа биореттегіштерді іздеу қажет [1, 2] . Гормондар - тірі организмдердің тіршілігі үшін маңызы өте зор. Организмдердің дамуы, өсуіндегі ең негізгі процестер гормондармен реттелуге тәуелді. Гормондар жетіспеген жағдайда немесе артып кететін болса онда организмді ауруға ұшыратады. Яғни осы қызметтеріне қарап гормондарды - организмді реттеуші патшалық деп айтуға болады. Өсімдік тіршілігін реттеуде өсімдік гормоны цитокининнің маңызы зор. Цитокининнің функциясы өсімдік онтогенезінің барлық кезеңдерінен көрінеді. Цитокининді зертеу аймағындағы жаңа жетістіктер мен көріністерге кейінгі жылдары ғалымдардың қолы жетуде [3, 4] . Цитокинин - клетка бөлінуін реттейді, хлоропластардың дифференцациясына қатысады. Клетка бөлінуі үшін цитокининнің маңызы зор. Осы гормонның сигнальды трансдукция механизмі әлі анықталмаған.
Трансдукция дегеніміз - сигналдың сигналға қатарымен берілуі. Трансдукцияның ерекшеліктері бар. Сигнал иерархиясы - жоғары дәрежелі сигнал төменгі дәрежелі сигналды тудырады, ал төменгі дәрежелі сигнал жоғары дәрежелі сигналды тудыра алмайды. Трансдукция сигналды эстафета тәрізді өткізіледі. Эстафета тәрізді сигнал әр сатыда өзгереледі, басқаша айтқанда жоғары сатылы сигнал тек қана көршіге әсер етеді, ол ары қарай сигналды бере алмайды. Сигналды трансдукцияның механизмінде медиатордың ролі өте маңызды. Медиатор - екінші сатыдағы сигналды беретін зат. Себебі гормон тікелей өзі емес медиаторлар арқылы клеткаға әсер ете алады. Сол себептен цитокинин медиаторын зерттеу қызығушылық тудырады [5] . Біздің мақсатымыз цитокинин медиаторын бөліп алып, тазартып оның морфогенез процесіне әсерін зерттеу; Осы мақсатқа жету үшін алға қойылған міндеттер: Цитокинин медиаторын бөліп алу; цитокинин медиаторын in vitro жағдайында өсіп жатқан бидайдың каллустарының морфогенезіне әсерін тексеру, Практикада цитокинин медиаторының қолдану жолдарын іздестіру.
1. ӘДЕБИЕТКЕ ШОЛУ
1. 1. Цитокининдердің ашылуы мен химиялық табиғаты
Клетка бөлінуін қоздыратын, морфогенез, қартаю процестерін, тыныштықты реттеуде іске қосылатын және алуан түрлі биологиялық белсенділіктің белгілі бір жиынтығына ие болатын фитогормондар типінің бірі - цитокинин болып табылады. Өсімдіктерден цитокининді 1955 жылы алғаш рет АҚШ-тағы Висконсинс университетінде Миллер мен Скуг ашты [6, 7] . Цитокининнің ашылуына келесі жағдайлар көмектесті. Авторлар темекі сабағы өзегінен алынған каллустармен жұмыс істей отырып, белгісіз фактордың жетіспеуінен оның өсуінің тоқтап қалатынын байқаған. Сондықтан ғалымдар каллустардың өсуіне жеткіліксіз белгісіз факторды ашытқы экстрактысынан бөліп алуды қарастырды. Осымен бірге бұл активті заттың құрамына пуриннің кіретіндігін де анықтады. Сонымен қатар табиғи пуриндік негіздің РНК және ДНК гидролизатындағыдай темекі сабағы каллусының өсуіне жағдай туғыза алмайтындығын тексерді. Байқаусызда активті болып бұрыннан сақталып келе жатқан ДНК препаратының эфирлі экстрактысы болып табылды. Сонымен активті затты кез-келген ДНК препаратын қышқылдық ортада автоклавтау жолымен оны дегидрациялау арқылы алуға болатындығын анықтады. Бұл зат кристал түрінде және иденфикациялау арқылы бөлініп алынды [8, 9] . Ол 6-фурфурил-аминопурин болып табылады. Өте активті цитокинин -6-(γ γ -диметилалил) ﹣аминопурин немесе изопентениладенин деп аталады. ол тасымалдаушы РНК құрамына минорлы негіз түрінде кіреді. 6-фурфуриламинопурин ИСҚ-ның қатысында темекі сабағының орталық ұлпасын оқшаулағанда клеткалық бөліну жүрген. Ол кинетин деп аталды. Түрлі синтетикалық цитокининнің активтілігі салыстырылып, тексерілді. Солардың ішіндегі ең активтісі- 6-(γ γ - диметилалил) аминопурин ал 6-бутил 6-амил-, және 6-гексиламинопурин активтілігі бойынша 6-БАП-қа жақындаған [9] . 1913-1923 жылы Г. Габерланд өсімдіктен бөліп алынған жапырақ тақташасының бетіне белгісіз заттың бөлініп шыққанын байқаған. Ол осы заттың өсімдіктің жарақат алған жерін қалпына келтіретін қасиеті бар екенін байқады. Осыдан кейін Габерланд бұл зат клетка бөлінуіне әсер ететінін, өсімдік жарақат алған кезде қалпына келтіру гормонына айналады деп тұжырымдады [10, 11] . Кокос “сүтінің ” құрамынан бөлініп алынған цитокининдер тобына жататын активті қосылыстың бірі - дифенилмочевина, өсімдіктер клеткаларының бөлінуіне күшті әсер етеді. Цитокинин тамырдың ұшында түзіліп, су ағынымен қоса жоғары қарай жылжиды [12] . Қазіргі кезде цитокинин микроорганизмдерден, балдырлардан, папортниктерден, мүктерден және көптеген жоғары сатылы өсімдіктерден табылған. Негізінен цитокинин клетканың индукциялық бөлінуін активтендіреді. бұл қасиетін жекеленген темекі ұлпасын арнаулы ортада өсіру кезінде байқаған. Сонымен қатар цитокининнің тағы бір қасиеті бұл мүше түзуге қатысуы. Цитокинин ауксинмен қатарласа тамыр, гүл, тағы басқа мүшелерді түзетіндігі ашылған болатын. Цитокинин споралы өсімдіктер мүк, папортниктердің мүше түзуіне қатысып, оны реттеп отырады. Генеративті мүшелердің түзіліп оның қалыптасып, тез жетілуіне де әсері бар. Бір сөзбен айтқанда цитокинин басқа гормондармен бірігіп морфогенез процестерін активтендіреді [13, 14] . Алғашқы табиғи цитокининді 1964 жылы Лезамом Letham бөліп алды, бұған дейін жүгерінің сүттену кезеңіндегі дәнін алып, спирттік экстрактысын бірнеше саты тазартудан өткізіп қолданған [15] . Цитокининді жүгеріден бөліп алынғандықтан оны зеатин деп атаған. Табиғи цитокининнің формуласы 2 - суретте көрсетілген. Сонымен қатар зеатин өріктің пісіп жетілгенінен, өсіп тұрған алма өнімінен микоризді саңырауқұлақтың культуралық сұйықтығынан табылған. Люпин ұрығынан - дегидрозеатинді тапқан. Сонымен бірге цитокиинин папортниктен, қыналардан, және балдырлардан табылған [16] .
Зеатинге басқа туыс заттар өсімдіктер бактериясынан Corynebacterium fascians патогендік культурасынан бөлінген. Өсімдікке оның әсері цитокинин препаратының әсеріне ұқсайды және апикальды басымдылықтың бұзылуымен байланысты. Осымен бірге зеатин және оған туыс цитокинин активтілігімен байланыстар өсімдіктер әлемінде кеңінен таралған.
Цитокининнің физиологиялық әсерінің бірі жапырақтың қартаюын кештетуінде. Бұл алғаш рет Richmond және Lang-тың жұмыстарын әйгілі етіп көрсетті. Цитокинин және апикалдық басымдылықты тежейді.
1955 жылы алғаш рет американ зерттеушісі Ф. Скугтың лабораториясында цитокинин ашылған. Олар in vitro жағдайында тәжірибе жасау кезінде өсімдікте белгісіз бір заттың клетка бөлінуін жылдамдататынын байқаған. Ол зат 6 - фурфураминопурин екенін анықтады. Бұл зат кейіннен кинетин деп аталды [17] .
Кинетеин - ақ түсті кристалды зат, құрылымдық формуласы C 10 H 9 N 5 O молекулалық салмағы 215, 5 Кинетин суда нашар ерісе, этанолда, сілтілі ерітінділерде, қышқылда жақсы ериді. Ол қыздырғанда қышқыл мен сілтілердің әсеріне өте төзімді келеді.
Қазіргі кезде цитокининдердің төмендегідей түрлері белгілі: кинетин, зеатин, 6- (γ γ - диметилалил) - аминопурин, дифенилмочевина, фитогормондардың бұл топтары аденин туындылары болып табылады.
Жетілмеген яғни пісе қоймаған жүгері дәндерінен анықталған цитокининді зеатин деп атайды.
Өте активті цитокинин - 6 - (γ γ - диметилалил) -аминопурин деп аталады. Ол тасымалдаушы РНК құрамына минорлы негіз түрінде кіреді. Кокос сүтінің ” құрамынан бөлініп алынған цитокининдер тобына жататын активті қосылыстың бірі - дифенилмочевина, өсімдіктер клеткаларының бөлінуіне күшті әсер етеді: Цитокинин тамырдың ұшында түзіліп, су ағынымен қоса жоғары қарай жылжиды [18] .
Қазіргі кезде цитокинин микроорганизмдерден, балдырлардан, папортниктерден, мүктерден және көптеген жоғары сатылы өсімдіктерден табылған. Негізінен цитокинин клетканың индукциялық бөлінуін активтендіреді. Бұл қасиетін жекеленген темекі тканін арнаулы ортада өсіру кезінде байқаған. Сонымен қатар цитокининнің тағы бір қасиеті бұл мүше түзуге қатысуы. Цитокинин ауксинмен қатарласа тамыр, гүл, тағы басқа мүшелерді түзетіндігі ашылған болатын. Цитокинин споралы өсімдіктер мүк, папортниктердің мүше түзуіне қатысып, оны реттеп отырады. Генеративті мүшелердің түзіліп оның қалыптасып, тез жетілуіне де әсері бар. Бір сөзбен айтқанда цитокинин басқа гормондармен бірігіп мүше түзу процестерін активтендіреді [19] .
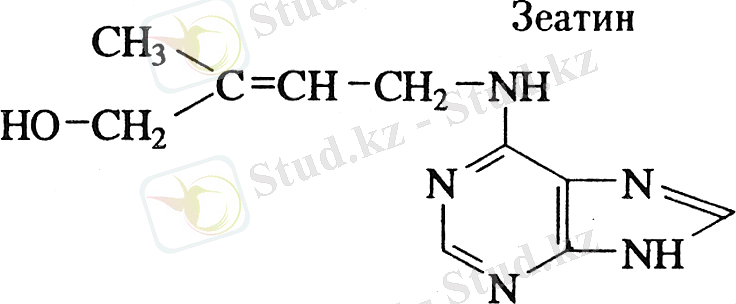
1-сурет Цитокининнің табиғи түрі - зеатиннің формуласы
1. 2 Өсімдік гормондарының атқаратын ролі
Кейінгі зерттеулердің барлығы дерлік өсімдік фитогормондарының өсу процесіне, клетка бөлінуіне әсерін зерттеу бағытында жүргізіледі. Небәрі 60 жыл ішінде бірінен соң бірі этилен, цитокинин ашылған болатын. Ал енді фитогормондарға толығырақ анықтама беретін болсақ фитогормон - бұл аз молекулалы органикалық салмағы төмен эндогендік зат. Клеткада, тканде өте аз мөлшерде болады. Фитогормондар клеткамен клетка, тканмен ткан, мүше мен мүше байланысқанда көптеген морфогенетикалық және физиологиялық программаларды атқарады. Фитогормондар өсімдік организміндегі заттардың мөлшеріне, маңыздылығына, ерекшелігіне көбеюге, клетканың бөлінуіне, тыныс алуға, қартаюға, зат алмасуға қатысады. Сонда әр топтың өзіне тән атқаратын қызметі бар. Мысалы:
- Өсімдік гормоны тек органикалық қосылыстардан тұрады. Оның молекулалық салмағы 28 ден (этилен) 346 (ГА) дейінгі аралықта болады. Өсімдік тканінен осы уақытқа дейінгі фитогормондар құрамынан белокты және полипептидті қосылыстар табылған жоқ.
- Фитогормондар клеткада, тканде бос физиологиялық активті күйде болады. Өте аз концентрациядада 10-5-10-10концентрацияда өте активті бола алады.
- Гормондар бір клеткада түзіліп, басқа клеткаларға әсер етеді. Бұл өсімдіктің барлық мүшелері бірдей бір қалыпта жұмыс істеуіне мүмкіндік жасайды.
- Фитогормондар физиологиялық және морфогенетикалық процестерді реттейді. Сонымен қатар генді аппараттың белоктарымен байланысады. Осы жағынан фитогормондар витаминдерден ерекшеленеді [20, 21] .
Өсімдіктердің гормондық жүйесінде фитогормондарды 5 топқа бөледі: ауксин, гибберлин, цитокинин, абсциз қышқылы, этилен. Бұл физиологиялық заттардың арасында көптеген байланыстар бар. Цитокинин-бүршік қалыптасуын реттейді. Фитогормонның физиологиялық әсері рецептормен байланысты. Рецепторлар - бұл берілген гормонның физиологиялық әрекетін іске асыру үшін қажетті, нәтижесінде биохимиялық реакциялардың рецепторларымен инициация жүретін гормонды қайтымды байланыстыру үшін қажетті жоғары спецификалық сайттарға ие, сәйкес нысана клеткаларында болатын химиялық құрылымдар. Сол себепті рецепторлар клетканың гормонға жауап беретіндігін ескертуде ең маңызды қызмет атқарады [22] .
Негізгі биохимиялық процестердің негізгі нәтижелері арқылы және жануарлар мен өсімдіктердің клеткаларында ерекше белгілерге ие рецепторлардың критерийлерін таратып қарауға болатындығы белгілі болды. Жануарлардың рецепторлары жәй және модификацияланған болып бөлінетіндігі белгілі, оларды негізінен екі негізгі типке жатқызады:
1. Клетка ішіне енген гормон молекулаларын байланыстырушы - клетка ішілік рецепторлар.
2. Клетка сыртындағы сұйықтықтағы мембрана үстіндегі гормондарды байланыстырушы - мембраналық рецепторлар.
Сонымен қатар бірінші типтік рецепторлар гормонмен байланысқан уақытта өзінің конформациясын өзгертіп, гормон рецепторлық комплекс түрінде клеткадағы әртүрлі метоболиттік процестерге әсер ету қабілеттілігіне ие бола алады. Екінші типтің рецепторлары берілген гормонның физиологиялық әрекетін іске асыратын заттар: с АМР, с GMP, Са 2+ , фосфоинозитид, диацилглицерин, екіншілік медиаторлардың клетка ішілік концентрациясының күрт үлкеюіне шақырады. Сонымен қатар, гормондарды рецепторларымен басқа рецепторлық емес гормон байланыстырушы белоктары болады. Олар: метаболизм ферменттері. Сондықтан оларды рецепторлардан ажырату үшін келесі критерилер қолданылады:
- Гормон мен рецепторлардың бір - бірімен қатынасы жоғары маманданған болуы қажет. Рецептор гормон молекуласының құрылымын танып, оны жоғары аффинділігімен байланыстыруы қажет.
- Гормон мен рецептордың байланысы қайтымды болуы қажет, бірақ гормон химиялық өзгермеген күйінде қалуы керек.
- Гормон мен рецептордың байланысуы үшін процесс қанықты болу қажет.
- Физиологиялық эффекті шақыратын, гормонның рецепторы және концентрациясы мен байланысуы қандырылған болса, гормондардардың концентрациялары арасында сәйкестілік болуы керек.
- Осы рецепторлармен ұлпа және мүше реттеуші клетканың үйлісімділігі болуы керек
Бұл жұмыста өсімдіктер гормондарының рецепторларын зерттеудегі соңғы жылдардағы жетістіктер келтірілген [23, 24] .
Этилен рецепторларындағыдай, N - ұшында гидрофобтық сегменттер бар: екі сегмент АНК 4-те және үш сегменттен АНК2 мен АНК3 -те. Негізгі фитогормондардың нысанасы ретінде өсімдіктер клеткалары қолданылады. Өсімдіктердің өсуі мен даму процестеріне әртүрлі гормондар әсер етеді. Сонымен қатар, бір гормон әсер ету орны және өсімдіктердің даму фазасына байланысты әртүрлі физиологиялық процестердің жүзеге асырылуына қатыса алады. Бұл жағдайда гормондардың әсерінің көп функциялдылығы өсімдіктердің гормональді жүйесін зерттеуде қосымша қиыншылықтар тудырады. Мысалы, бір өсімдіктердегі жан-жақты жалғыз рецептор арқылы әртүрлі бағдарламаларды индукциялайтындығы әлі де табылмаған. Өсімдіктер мен жануарлардың гормондары химиялық жағынан ұқсас емес, ал белгілі гормондардың жалпы саны яғни өсімдіктердің гормондары жануарлар гормондарына қарағанда саны жағынан аз. Бұл жағдайда гормондар биохимиялық реттеуші - өсімдіктерде ең маңызды және жалғыз реттеуші болып табылады, ал бұл жануарлардың нерв реттеушісіменен қатар әсер етеді. Дегенмен өсімдіктер клеткаларының фитогормондарға спецификалық реакциялардың мүмкіншілігін ескере отырып, жануарлардың гормональды информациясын іске асырудағы молекулалық механизмдерін зерттеу нәтижесінде өсімдіктер клеткаларындағы гормондардың реттелуін және трансдукциясын зерттеуде үлкен табыстарға жетті. Мысалға алсақ өсімдіктерде де жануарлар секілді гормон рецепторларының екі типі бар екені белгілі болды. Рецепторлардың бірінші типіне цитоплазмалық және ядролық локализацияның ерігіш белоктары жатады, олар гормондармен байланысқа түсіп, гормон рецепторлық комплекс түзеді. Екінші типтің рецепторлары мембранада жинақталған, сонымен бірге, гормонмен мембрананың сыртқы бетінде орналасқан рецептор домені әсерлеседі. Гормон - рецепторлық әсерлесуінің нәтижесінде, белок - рецепторындағы конформациялық өзгерістер, екіншілік сарапшылардың түзілуін шақыратын, G - белок арқылы ферменттердің біреуіне (мысалы: фосфолипазалар, аденилатциклазалар) берілетін спецификалық сигналды индукциялайды. Нәтижесінде клеткада амплификацияға және әртүрлі клеткалық құрылымдарға берілуіне әкелетін екіншілік сигналды молекулалар пайда болады. Екіншілік сарапшыларға сАМФ молекулалары, инозитол үш фосфат, және басқа да мембрана липидтерінің жатады. Олардың кейбіреулері клетка метаболизмін өзгертумен қатар ондағы ферментерді, құрылымдық белоктарды модификациялайтын протеинкиназаларды немесе протеинфосфотазаларды белсендіреді. Көп жағдайда сигналды тапсыруда Са 2+ қатысады, ол клеткаға әсер еткен кезінде құрамын өзгертіп отырады. Бұл жағдайда кальций мембрананың сыртқы бетінде гормон молекуласымен рецепторды байланыстыру арқылы индукцияланған соң өзгерістердің өсуіне қатысады. Бірінші және екінші типтің рецепторлары цитокининдердің сигнальды трансдукциясына қатысады [25, 26] .
Фитогормондардың рецепторларын іздеуде екі маңызды бағыт болды:
1. In vitro жүйесінде біріншілік гормональды жауаптардың ұдайы өндірілуі.
2. Шашыраңқы гормондарды спецификалық түрде байланыстыратын белоктарды іздеу. Дегенмен бірінші бағыттың дамуы күрделі болып шықты, ал тәжірибелердің нәтижелері аз өндірілетіндікке және қайшылыққа әкеліп соқтырды. Екінші бағыттың нәтижелері үлкен табыстарға, жетістіктерге жетті - онда фитогормондардың рецепторлары болатындығын, соның ішінде цитокининдер бар өсімдіктерде көптеген белоктар табылды [27] .
1. 3 Цитокинин медиаторының өсімдік морфогенезіне әсері
Фитогормондар - өсiмдiктердің өсуiн реттегiштер. Өте аз мөлшерде болса да жоғары сатыдағы өсiмдiктердiң зат алмасу процессiне әсер ететiн табиғи және синтетикалық органикалық қосылыстар тобы. Осы реттегiштердiң әсерiнен өсiмдiктердiң өсуi және дамуы бiршама өзгерiстерге ұшырайды.
Табиғи реттегiштер бiзге фитогормондар ретiнде белгiлi. Фитогормондардың көмегiмен клеткаларда, ұлпалар мен органдарда физиологиялық және морфологиялық бағдарламалар реттелiп iске қосылады. өсiмдiктер гормоны - әртүрлi органдар мен ұлпаларда түзiлетiн органикалық заттар. Олардың химиялық құрамы және физиологиялық әсерi әр алуан. Фитогормондар өсiмдiктердiң өздерiнiң өнiмi бола тұра мүше түзу процестерiн анықтауға және метаболизм процессiн реттеуге қатысады. Морфогенетикалық және физиологиялық бағдарламалардың iске қосылуы немесе iстен шығарылуы фитогормондар мөлшерiнiң ара қатынасының өзгеруiмен жүргiзiледi [28] .
Өсiмдiк гормондарын негiзгi бес топқа жiктейдi: ауксиндер, гибберелиндер, цитокининдер, абсциз қышқылы және этилен. Қазiргi кезде
фитогормондардың өзiндiк әсерiн мәлiмдейтiн көптеген деректер бар. Әрбiр фитогормонның физиологиялық активтiлiгi жоғары және арнайы әсер етедi. Жалпылама айтатын болсақ: цитокининдер клетканың бөлiнуiне, ауксиндер мен гиббереллиндер клетка мөлшерiнiң ұлғаюы мен жiктелуiне, абсциз қышқылы тыныштық күйге, этилен жемiстiң пiсiп жетiлуiне әсер етедi.
Фитогормондардың басты белгiсi - өсiмдiк бойында жылжи алу қабiлеттiлiгi. Гормондық жүйе барлық мүшелерiн iске қосатын заңды бiрiздiлiк жеке даму сатысымен сәйкестендiрiлген. Өсiмдiк организмiнiң функционалды бiртұтастығын қамтамассыз етедi.
In vitro жағдайында морфогенез процесi мен регенерацияға фитогормондар мен химиялық өсу реттегiштерiнiң әсерi жөнiнде әдебиеттерде көптеген мәлiметтер бар. Қоректiк ортаның құрамына қосымша гормондар енгiзу арқылы морфогенез процесiнiң жолдарын реттеуге болады. Скуг пен Миллердiң тәжiрибесiндегi көрсетiлгендей ауксин мен цитокининнiң мөлшерлiк ара қатынасын өзгерте отырып, морфогенез типiн анықтауға болады. Осы гормондар концентрациясын өзгерту арқылы регенерациялық қабiлетi жоғары немесе төмен каллустар алуға болады. Цитокинин концентрациясы ауксиннен басым болғанда бүршiктер мен сабақтар түзiлген, ал керiсiнше жағдайда тамыр түзiлуіне әкелiп соққан, екi гормонның концентрациясы шамалас болғанда ұйымдаспаған каллустар өскен. Гормондар мөлшерiн өзгерту арқылы жүргiзiлген зерттеулер көрсеткендей жасанды ортада өсiрiлген ұлпалардан сабақ бүршiктердiң түзiлуi каллус ұлпаларының гормондарының концентрациясының физиологиялық және диффузиялық градиентiнiң өзгеруiмен байланысты екен [29] .
... жалғасыСіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz