Цитология және гистология: университеттік лекциялар жинағы

Қ. Ә. Сапаров
ЦИТОЛОГИЯ
ЖӘНЕ ГИСТОЛОГИЯ
Лекциялар жинағы
Алматы 2004
Әл-ФАРАБИ атындағы ҚАЗАҚ ҰЛТТЫҚ УНИВЕРСИТЕТІ
Қ. Ә. Сапаров
ЦИТОЛОГИЯ
және
ГИСТОЛОГИЯ
Лекциялар жинағы
Алматы
«Қазақ университеті»
2004
ББК 28. 070я73
С21
Баспаға әл-Фараби атындағы Қазақ ұлттық
университеті биология факультетінің Ғылыми кеңесі және Редакциялық-баспа кеңесі ұсынған
П і к і р ж а з ғ а н д а р
биология ғылымдарының докторы, профессор С. Т. Төлеуханов;
биология ғылымдарының докторы, профессор Қ. С. Рымжанов;
биология ғылымдарының кандидаты, доцент Р. Әбилдинов;
биология ғылымдарының кандидаты, доцент Ж. М. Базарбаева;
Сапаров Қ. Ә.
С 21 Цитология және гистология. Лекциялар жинағы: Оқу құралы. - Алматы: Қазақ университеті, 2004. - 89 бет.
ISBN 9965- 12-857-X
Бұл оқу құралының 1-бөлімінде клеткалардың құрылысы, дамуы, өсіп-өнуі, сондай-ақ зат алмасу кезіндегі қызметі туралы баяндалады. Клетканың жалпы құрылысы, оның құрамындағы негізгі құрылымдар (ядро, мембраналар, органойдтар) туралы жеке-жеке айтылады.
2-бөлімінде ұлпалардың құрылысы, қызметтері, олардың ерекшеліктері туралы мағлұматтар берілген. Эпителий, ішкі орта, бұлшық ет және нерв ұлпаларына түсініктемелер берілген. Оқу құралы жоғары оқу орындары биология факультеттерінің студенттеріне арналған.
С 1 90303-168 ББК 28. 070я73
ISBN 9965-12-857-Х Сапаров Қ. Ә., 2004
Әл-Фараби атындағы ҚазҰУ, 2004
Алғы сөз
Бұл оқу құралында цитологияның өзекті тақырыптары: клетка теориясы, клетканы зерттеу тәсілдері, цитоплазма, органоидтар, ядро, клетканың өмір сүру циклі, клетканың бөліну жолдары туралы сөз болады.
Оқу құралының екінші бөлімінде гистологияның тақырыптары қамтылады. Эпителий, ішкі орта, бұлшық ет, нерв ұлпаларының құрылыстары, қызметтері, ерекшеліктері баяндалады.
Цитология мен гистология университтердің биология факультеттерінде, медицина және аграрлы университтерінде негізгі пән ретінде оқытылып жүр. Бұл салада қазақ тілінде оқулықтар шыға бастады, әлі де бұл пәндерден оқулықтар жетіспейді.
Бұл оқулықты жазуда автор өзінің әл-Фараби ұлттық университетінде ондаған жылдар бойы дәріс беру тәжірибелеріне сүйенеді.
Кітаптың мазмұнын толықтыруға бағытталған ниеттеріңізді автор ілтипатпен қарсы алады.
ЦИТОЛОГИЯ ІЛІМІ ЖӘНЕ КЛЕТКА ТЕОРИЯСЫ ТУРАЛЫ ҰҒЫМ
Цитология клеткалардың құрылысын, атқаратын қызметін, дамуын зерттейтіи ілі. м. Грекше kytos - клетка, logos- ілім деген мағынаны білдіреді.
Цитологияның биология ілімінен өзінің іргесін бөлгеніне небары жүз жылдай ғана уақыт өтті. Осы аралықта ол жедел қарқынмен дамып, жазбаша морфологиялық ілімнен экспериментальды ілімге айналды. Сөйтіп цитология бүгінгі таңда клетканының құрылысын ғана емес, ондағы күрделі физиологиялық процестерді де зерттейтін ауқымды ғылым саласының біріне айналып отыр.
Морфология ілімінен өрбіген цитология анатомия, гистология, физиология, эмбриология, генетика, биохимия т. б. ілімдерімен тығыз байланыса келіп, клетка физиологиясы, цитохимия, цитогенетика, цитоэкология, салыстырмалы цитология сияқты өзінің төл тармақтарын туындатты.
Цитология да биохимия, биофизика, генетика және молекулалық биология салаларындай ғылыми әдістемелік тәсілдерге жүгінеді. Осы тәсілдер арқылы ол соңғы жылдары клетканы жан-жақты зерттеуде нәтижелі жетістіктерге жетті.
XIX ғасырдың басында жүргізілген микроскопиялық зерттеулер жануарлар мен өсімдіктер организмдерінің клеткадан құрылатынын дәлелдеп қана қоймады, органикалық дүниенің даму заңдылықтарын ашып берді. Я. Э. Пуркине және И. П. Мюллер ұйымдастырған ғылыми мектептер өмірге клетка теориясы жөнінде көп жаңалықтарды әкелді. Жалаң физиологиямен және фармакологиямен айналысқан Пуркине енді өзінің ғылыми бағыт-бағдарын өсімдіктер мен жануарлар клеткаларын зерттеуге қарай. бұрды.
Клетка теориясы ашылғанға дейін биология саласында оптикалық құралдармен жабдықтау, оны жетілдіру сияқты күрделі жұмыстар жүргізілді. Сөйтіп өсімдіктер мен жануарларды зерттеуде алғашқы мағлұматтар алына бастады. 1665 жылы Роберт Гук тұңғыш рет үлкейтіп көрсететін шынының көмегімен тозағашының құрылысын зерттеп, оның «клеткадан» тұратынын анықтады. Кейін өсімдіктердің өсіп дамуын бақылай келе М. Мальпиги (1671), II. Грю (1671) бұл жаңалықтарды толық. дәлелдеді.
А. Левенгук (1680) бірінші рет қан құрамында эритроциттердің барын анықтаса, Фантана (1781) жануарлар клеткаларындағы небір құпияларды ашты. Осыдан кейін өсімдіктер мен жануарлардың клеткаларының құрылыстары белгілі бола бастады. Қлетканың құрамындағы негізгі элемент - протоплазма (Пуркине 1830) мен ядро (Браун 1833) табылды. Осы мағлұматтарды негізге алып әрі әр түрлі ұлпалардың құрылысын, дамуын жанжақты зерттеп, соңынан нәтижелі қорытындыларын саралай отырып, 1838-1839 жылдары Т. Шванн өзінің атақты клетка теориясын жазды. Бұл жаңалық табиғаттану ғылымдарында бұрын-соңды болмаған ұлы жетістіктердің бірі еді. Т. ІІІванның тұжырымы бойынша клетканың пайда болуы өсімдіктерге де жануарларға да қатысы бірдей заңдылыққа бағынады. Ғалымның ой елегінен өткізілген осы қағида органикалық дүниенің даму заңдылығын тағы да бір қырынан көрсетті.
Ф. Энгельстің клетка теориясын XIX ғасырдағы ұлы жаңалықтардьң бірі деп атауына да осы негіз болса керек.
Клетка зерттеу әдістері
Цитололгияда негізгі колданылатын әдістердің бірі-жарық микроскопы. Соңғы жылдары клетканы зерттеуде жарық микроскоптарының бірнеше түрлері қолданылып жүр (люминесценттік, фазасы карама-карсы, электронды микроскопты) . Жарық микроскоптарының көмегімен ұлпадан алынған және әр түрлі бояулармен боялған жұқа кесінділерді (препарат) зерттеуге болады. Ол үшін кесіндінің қалыңдығы 5-7 микроннан (мк) аспау керек, сонда ғана жарық кесінділер арқылы өте алады. Жарық микроскоптары арқылы тексеретін ұлпалардан кесінділер дайындау (препарат) өте күрделі жұмыс. Цитологиялық препараттар жасау бірнеше кезеңдерге бәлінеді: материал алу және оны бекіту, ұлпаларды тығыздау, парафинге құю, кесінділер жасау, бояу, бальзамға бекіту.
Микроскоптың көру қабілеттілігі қолданылып отырған жарық ағымына байланысты және жарық ағынының 1/3 бөлігіне тең болады. Жарық толқынының ұзындыры неғұрлым қысқа болса, микроскоптың көру қабілеттілігі соғұрлым артады. Егер жарық толқынының ұзындығы 0, 6 милли микрон (мкм) болса, микроскоптың кәру қабілеттілігі- 0, 2 мкм- 1/3 ХО, 6 мкм - 0, 2 мкм. Люмиесцент микроскопы ультракүлгін жарық толқынымен жұмыс істейді, толқын ұзындығы- 0, 27-0, 4 мкм. Осындай толқын препаратқа түскенде ол сәулені сіңіре отырып, өзінен жарық шығарады, бүл құбылыс флюоресценция деп аталады. Шыққан жарық толқыны сінген жарық. Толқынына қарағанда әрдайым ұзын болады. Кейбір заттар түскен жарық толқынының жартысын сіңіріп, өзімен жасыл, сары, қызыл спектрді шығарады. Флюоресценция деп заттарды ультракүлгін жарығымен шағырылыстырылғанда өзінен жарық бөлуін айтады. Оларға пигменттер, витаминдер, майлар жатады. Кейбір заттарды флюрохром бояуларымен бояу арқылы флюоресценцияны көруге болады. Мысалы, ДНК-ны акридин қызыл сары бояуымен боялғанда клеткадағы дизоксирибонуклеин қышқылы (ДНҚ) ашық жасыл сәуле береді, ал рибонуклеин қышқылы (РНҚ) ашық қызғылт сәуле береді.
Фазасы қарама-қарсы микроскоп.
Бүл микроскоптың ерекшелігі тірі клеткаларды, боялмаған объектілерді зерттеуге мүмкіндік береді.
Электронды микроскоп.
Электронды микроскоптарда жарықтың орнына электрон сәулелері қолданылады, осыған байланысты қолданылатын қуаттың күші 50-100 кВ-қа дейін барады, ал толқын үзындығы 0, 056-0, 035 А°-ге жетеді. Толқын ұзындығы неғұрлым қысқа болса, микроскоптың көрсеткіштік қабілеттілігі сорғұрлым артатынын физика курсынан жақсы білеміз. Осыған байланысты электроиды микроскоптардың көрсеткіштік қабілеттілігі -1-7А°-ға, ал үлкейткіштік қабілеттілігі 600 000-ға дейін жетеді. Электронды микроскопты пайдаланып қалыңдығы 400-600А° препаратты көруге болады, өйткені қалың препараттан электрондар өте алмайды, олардың өткізгіштік қасиеті нашар. Электронды микроскопқа препарат дайындайтын приборды ультрамикротом деп атайды. Осы прибордың көмегімен жұқа кесінді жасап, оны объекті торына бекітіп, арнайы бояулармен бояп, электронды микроскоппен қарайды. Электрон сәулелері препарат арқылы өткенде объектінің үлкейтілген «көлеңкесі» экранға түседі.
Цитохимиялық және гистохимиялық әдістер клетканың, ұлпаның құрамындағы химиялық заттардың мөлшерін, саласын тексеруге мүмкіндік береді. Бұл әдіс химиялық реакциялар арқылы клеткалар мен ұлпалардың құрамындағы қосылыстарды тексереді. Қазіргі кездерде қолданылып жүрген тәсілдер арқылы клетканың құрамындары белоктарды, нуклеин қышкылдарын (ДНҚ, РМҚ), ферменттерді анықтауға болады. Мұндай заттарды анықтау үшін химиялық қосындылар мен клеткалардың немесе ұлпаның арасындағы реакцияның нәтижесінде заттардың боялып көрінуін қамтамасыз ету керек. Мысалы, клетканың құрамындары РНҚ-ны анықтау үшін - галлоцианин бояуы қолданылады, ал оның клеткада барын білу үшін рибонуклеазамен РНҚ-ны ажыратады (немесе жояды) . Сәуле спектрін өзіне сіңіру арқылы клетканың құрамындағы химиялық заттардың мөлшерін есептеуге негізделген приборды цитоспектро-фотометр деп атайды. Бұл әдіс абсорбция заңына бағынады.
Авторадиография .
Бұл әдіс клеткадағы зат алмасу процесін зерттеуде қолданылады. Ол үшін фосфор- (Р 32 ), көміртегі (С14), сутегі II'3) радиоактивті элементтер немесе олардың қосындылары пайдаланады.
Клеткаларды толық және жан-жақты зерттеуде басқа да әдістер қолданылады: ұлпалар мен клеткаларды өсіру, микрохирургия т. б.
ЦИТОПЛАЗМА
Тірі организмдердің құрамына кіретін клеткаларды екі топқа бөлуге болады: ядро қабаты болмайтыны прокариоттар оларға көк жасыл балдырлар және бактериялар жатады (1-сурет), ядро кабаты жақсы жетілгені эукариоттар, бұған өсімдіктер мен жануарлар клеткалары жатады (2-сурет) .
Эукариотты клеткалар үш бөлімнен тұрады: сыртын қоршап жатқан плазматикалық мембранадан, цитоплазмадан және ядродан (3-сурет) .
1-сурет. Прокариот клеткаларының қиыстырылған үлгісі: 1 - клетка қабығы, 2 - плазматикалық мембрана, 3- нуклеоид аймағындағы ДНК, 4 - цитоплазманың полирибосомалары, 5 - мезосома, 6 - ламеллалар құрылымдар, 7- плазматикалық мембрананын, клетканың ішкі жағына ығысуы, 8 - хроматофоры, 9 - клетка қосындыларының вакуольдері, 10 - талшықтар, 11- қатпарлы тилакоидтар.
Прокариоттар мен өсімдік клеткаларының плазматикалық мембраналарының сыртында клетка қабаты болады, ал жануарлар клеткаларында мұндай қабат болмайды. Клетканың ядросын қоршап жатқан қоймалжың затты цитоплазма деп атайды Эукариотты клеткалар цитоплазмасының әртектілігі оның құрамында гиалоплазманың болуынан. Гиалоплазма мембраналы және мембранасыз компоненттерден тұрады. Мембраналы ком-поненттерге митохондриялар, пластидтер, эндоплазмалық тор, Гольджи аппараты, лизосомалар, ал мембранасыз компоненттерге центрольдар, рибосомдар, микротүтікшелер, микрофиламенттер жатады. Цитоплазманың кұрамындағы жоғарыда аталған компоненттер бір-бірімен өте тығыз байланыста болады.
Клетканың зат алмасу процесінде гиалоплазма үлкен рөл аткарады. Цитоплазманың негізгі матриксы (негізгі плазмасы) болып саналатын гиалоплазма - клетканын өте маңызды бөлімі. Электронды микроскоптан қарағанда цитоплазманың кұрамында гомогенді жұқа түйіршікті денелердің, бары байқалады. Цитоплазманың физикалық және химиялық ерекшеліктеріне келетін болсак, ол
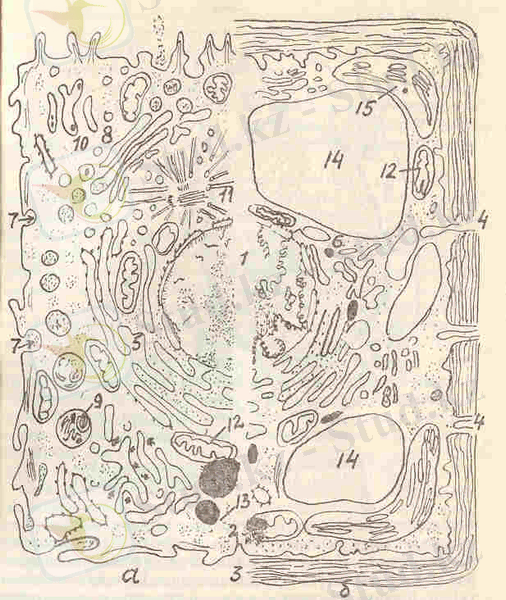
2-сурет. Эукариот клеткаларының қиыстырылған үлгісі:
а - жануар клеткасы, б- өсімдік клеткасы; 1- хроматыны және ядрошығы бар ядро, 2 - плазматика мембранасы, 3 - клетка қабығы, 4 - ІІлазмодеома, 5 - кедір-бұдырлы эндоплазмалық тор, 6 - тегіс эндоплазмалық тор, 7 - пиноцитоз вакуолі, 8 - Гольджи аппараты, 9 - лизосом, 10 - тегіс эндоплазмалық тордағы май тамшылары, 11- центрольдер, микротүтікшелер жоие центросфералар, 12 - митохондрия, 13 - гиалоплазманың полирибосомдары, 14 -- орталық вакуоль, 15 - хлоропласт.
Негізінде кұрамында белоктар, нуклеин қышқылдары, көмірсулар болатын күрделі қоймалжың заттардан тұрады. Бұл жүйе сұйық күйден қатты күйге немесе керісінше ауысып отырады. Бұл жағдай көбіне сыртқы әсерлермен байланысты. Егерде клеткаға жоғарғы гидростатикалық қысыммен әсер ететін болсақ, цитоплазма қатаймай сұйыла бастайды. Бұл құбылыс молекулалардың арасындағы байланыстардың нашарлауынан туады. Физика-химиялық және электронды-микроскопиялық әдістерді қолдана отырып зерттеулердің арқасында негізгі плазма деген түсінік пайда болды. Бұл көп компонентті цитоплазманың жұмысын реттеп, үйлестіріп тұратын жүйе болып саналады.
Атқаратын қызметтеріне карай цитоплазманың әрбір бөлімдері өздерінің агрегатты күйін өзгертіп отырады. Мысалы, кейбір жағдайларда тубулин-белоктарының молекулалары цитоплазмада шашырап, ал белгілі бір мезгілде жинақталып, микротүтікшелер түзеді. Микротүтікшелердің өздігінен жиналуы- қайтымды процесс. Егер де клетканың өмір сүру жағдайын өзгертсек (қысымды көбейтсек немесе клетка мембранасының өзгергіштігін өзгертсек), онда микротүтікшелер тубулин молекулаларына дейін ыдырап кетеді. . Сонымен гиалоплазмада белок молекулаларының кешені пайда болып және ыдырап отырады.
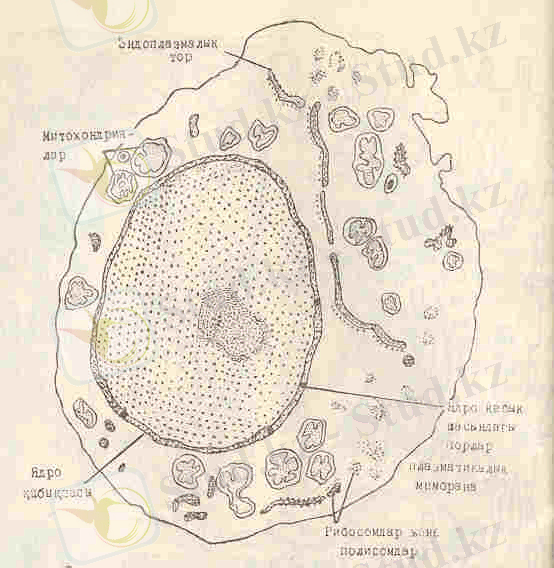
3-сурет. Клетканың жалпы құрылысының үлгісі. (Ж. -Х. Ролан, А. Селоши, Д. Селоши бойынша) .
Егер клетка гомогенатынан оның ядроларын, мембранадан тұратын органоидтарын, рибосомдарын тұндырып алып, қалған сұйықтықты (цитозоль) тексерсек, онда оның күрамында гиалоплазманың негізгі химиялық құрамын табуға болады.
Гиалоплазманың құрамында белоктар, цитоплазмалық ферменттер көрінеді. Эукариотты клеткалардағы белоктың 20-25%-і осы гиалоплазманың кұрамында бар. Бактерия клеткасының гиалоплазмасы барлық белоктардың 50%-тін қамтиды. Гиалоплазма матриксында гликолиз ферменттері, қант метаболизміне қатысатын ферменттер, азот негіздері, аминкышқылдары, майлар және т б. күрделі қосылыстар бар. Матрикс кұрамында белок синтездеу барысында аминқышқылдарын күшейтетін ферменттер де баршылық. Гиалоплазма - клетканың барлық құрамдарын біріктіріп тұратын және бір-бірімен химиялық қатынасын камтамасыз ететін орта. Гиалоплазма арқылы клетка ішіндегі аминқышқылдары, нуклеотидтер: (қанттар, майларды тасымалдау) процесі жүреді. Гиалоплазма АТФ молекулалары жинақталатын орын. Мұнда гликоген. май тамшылары сияқты қоректік заттар жиналады.
Соңғы жылдары меговольтты электронды микроскоптың көмегімен К. Р. Портер клетканың гиалоплазмасынан микротрабекулярлық тор тапты. Осы тордың негізінде цитоплазманың ішкі (2-нм) талшықтардан түратынын байқаған. Трабекуля жүйесі - гиалоплазмадағы бүкіл жүқа түтікшелерді, жіпшелерді, мембраналы органеллаларды және плазматикалық мембрананы біріктіріп тұратын тор. Трабекуля- тордың түйісетін не косылатын нүктелеріне рибосомдар I (полисомдар) орналасады. Сонымен трабекуля жүйесі гиалоплазманы белоктарға бай полимерлі фазаға және трабекулялар арасындағы сүйықтық фазаға бөледі. Трабекуля жүйесі клетканың ішкі тірегі болумен катар, цитоплазмадағы ферменттердің орналасуын реттеп отырады. Трабекуля жүйесі - козғалмалы жүйе. Ол сырттан келген әсерлердің салдарынан ыдырап та кете алады.
Цитоплазма мембраналары.
Клеткада кездесетін барлық мембраналар (плазматикалық органоидтардың мембраналары)

4-сурет. Мембрананың молекулалы қүрылысы. (Ж. -К. Ролан, А. Селоши, Д. Селоши бойынша) .
Мембраналардың кұрылыстарын анықтайтын көптеген модельдер ұсынылды. Олар негізінде белоктар мен фосфолипидтердің байланысын көрсетеді. Кейбір модельде мембрана үзілмейтін қатпарлы құрылым (а) кейбіреуінде мицелла (б) түрінде көрсетіледі. Мембрананың ішкі глобулярлы түйіршіктері ашылғаннан кейін мембраналарды белоктар кіретін сұйық қүрылым ретінде қарайды және ішкі клетка мембраналары кұрылысы және химиялық құрамы жағынан бір-біріне ұқсас келеді. Электронды микроскоптан мембрананың үш кабаттан тұратынын анық көруге болады. Ортаңғы ашық кабат бимолекулалы май қабатынан, ал шеткі күңгірт қабаттары белок молекулаларынан түзіледі (4-сурет) . Клетканың сыртқы қабаты (плазматикалық) оны сыртқы ортадан бөліп тұрады, ал ішкі мембраналар көпіршік түрінде келіп, өзінің құрамындағы заттарды цитоплазмадан ажыратады.
К. летка цитоплазмасындағы мембраналар қуыстар, түтікшелер немесе көпіршіктер жасап, өздерінің ішіндегі заттарды гиалоплазмадан бөліп, күрделі тор құрайды. Мұндай мембраналарға митохондрия, пластидтер, эндоплазмалық торлардың мембраналары жатады (5-сурет) .
Плазмалемма. Плазмалемма. клетканы қоршап тұрғандықтан сыртқы ортамен тікелей байланысты, сондықтан бұл мембрананың клетка тіршілігіндегі рөлі өте зор. Плазмалемманың қалыңдығы 10 нм шамасындай, басқа мембраналарға қарағанда қалыңдау, өйткені оның сыртқы беті көмірсулардан, ішкі жағы қалың белок молекулаларынан тұрады. Мембрананың негізгі химиялық компоненттері - белоктар (60%), майлар (40%) және көмірсулар (2-10%) . Клетканың ішкі мембраналарына қарағанда плазмалемма холестеринге бай.

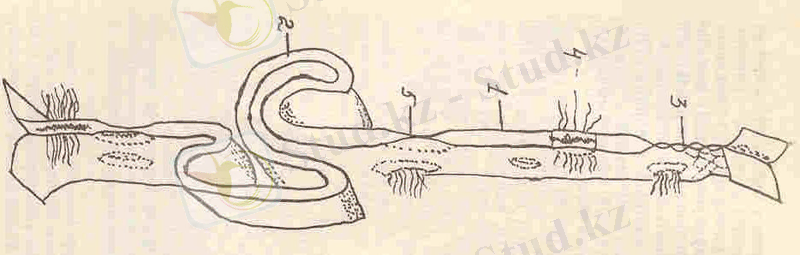
5-сурет. Клетканың ішкі мембраналарының үлгісі:
1 - ядро қабықшасы, 2 - ядро қабықшасындағы торлар, 3 - полисомалар, 4 - кедір-бұдырлы эндоплазмалық тор, 5 - жазық эндоплазмалық тор, 6- ауыспалы көпіршіктер, 7 - вакуоль, 8 - диктиосома, 9 -плазматикалық мембрана, 10- цитоплазмалық көпіршік.
Плазмалемманың бір ерекшелігі оның сыртында көмірсулардан тұратын гликокаликс қабаты орналасады. Бүл қабаттың қалындығы 3-4 нм-дей болады. Плазмалемма негізгі атқаратын қызметі: қорғаныштық, өткізгіштік және тасымалдаушы. Тасымалдаушы плазмалемма сулардың, иондардың және молекулалардың сыртқы ортадан клеткаға өтуін және кері өтуін реттеп отырады. Зат алмасу процесінде клеткада пайда болған қорытылған заттар да осы плазмалемма арқылы сыртқа шығарылып отырады. Плазмалемманың сыртқы бетіне рецепторлық ферменттер орналасады, олар клетканың күйін басқа көрші клеткаларға жеткізіп тұрады. Плазмалемма клетканың бөліну процесінде маңызды рөл атқарады. Оның сыртында микротүтікшелер, талшықтар сияқты әр түрлі өсінділер болады.
Плазмалемманың тасымалдаушы қызметі.
Сыртқы ортадан клеткаға әр түрлі заттар енетінін атап кеттік. Үлкен молекулалар жай өтеді де, кіші молекулалар жылдам өтеді. Ең жылдам өтетін су және оның құрамындағы ерітінділер екен. Егер эритроцитті өзінің цитоплазмасынан гөрі шоғырлануы аз тұзды ерітіндіге салсақ, онда сыртқы ортадан су клеткаға көп енеді де клетканың көлемі өсіп, оның сыртқы мембранасы жарылып кетеді. Керісінше, егер эритроцитті шоғырлануы көп тұзды суға салсақ, онда клеткадағы су бөлініп шығады да, ол қабысып, жиырылып қалады. Клетканың сыртқы мембранасында линопротеин қабатында «порлар» болады, олар арқылы иондар және сулар өтеді. Осы мембрана арқылы К+, N+ катиондары аса жылдамдықпен жүреді. Мембрананың тасымалдау қызметі: иондар мен судың шоғырлануы көп ортадан шоғырлануы аз ортаға (пассивті тасымалдау), және керісінше, шоғырлануы аз заттың Шоғырлануы көп ортаға енуін (активті тасымалдау) реттеп отырады. Активті тасымалдау энергия жұмсау арқылы өтеді (АТФ), Қ+, Nа+ -иондарының шоғырлануын реттеп тұратын К+, Nа+-«насосы» плазмалеммаға орналасады. Осы «насостардың» жұмыстарын зерттеу үшін эритроцитті алуға болады. Бұл клеткада К+, Nа+ иондарының шоғырлануы қанның плазмасына карағанда өзгеше болады (бірақ концентрациясының жиынтығы шамалас, клетканың сыртында және ішінде изотониялы) . Белоктар, нуклеин қышқылдары, майлар, ыдырап барып мономерлер түрінде ғана плазмалеммадан өте алады.
Кейбір жағдайларда макромолекулалар немесе ірі түйіршіктер клеткаға эндоцитоз процесі арқылы өтуі мүмкін. Эндоцитозды фагоцитоз жәме пиноцитоз деп екі түрге бөлуге болады: фагоцитоз процесі дегеніміз клеткалардың ірі түйіршіктерді кабылдап цитоплазмасына өткізуі. Бұл процесті бірінші рет зерттеген орыс ғалымы - И. И. Мечников. Пиноцитоз процесінде клетка цитоплазмасына ерітінділерді қабылдайды. Плазмалемма арқылы ішінде сұйық заттары бар көпіршіктердің клеткаға өтуін пиноцинтоз деп атаймыз. Эндоцитоздың бірінші кезеңінде (адсобция) энергия жұмсалмайды. Сырттан келген көпіршіктер плазмалемманы итеріп, ойыс жасап барып өтеді немесе плазмалемма өсінділері біртіндеп клеткаға енеді, клеткаға енген көпіршіктер плазмалеммадан жеке бөлініп барып орналасады. Клеткаға түскен түйіршікті заттар лизосомадағы гидролаза ферменттер арқылы қорытылады. Пиноцинтоз процесін көптеген клеткалардан, яғни өзіне қоректік заттарды сіңіретін, мысалы ішек клеткаларынан(энтороциттер) байқауға болады. Энтороциттердің жоғарғы (апикальді) бөлімінде пиноцитоз көпіршіктері орналасады, олар клеткаға ыдырап майларды, кемірсуларды т. б. өткізіп отырады.
Пиноцитоз процесі арқылы клеткара заттардың өтуін жануарлар клеткаларынан, жоғарғы сатыдағы өсімдіктерден, төменгі саңырауқүлақтардан, қарапайымдардан, тіпті бактериялардан да байқауғра болады.
Плазмалемма заттарды клеткадан шығаруға эндоцитозға қарама-қарсы процесте - экзоцитозда маңызды рөл атқарады. Мысалға әр түрлі белок молекулалары, ферменттер, мукополисахаридтер, май тамшылары экзоцитоз арқылы клеткадан шығарылады. Клеткадан көп мөлшерде шыққан ферменттер клетканың жоғарғы бетіндегі гликокаликс қабатында жиналады. Ферменттер биополимерлерді және органикалык қосылыстарды мембранадан тыс ыдыратуда үлкен қызмет атқарады.
Әр түрлі иммунологиялық әдістердің көрсетуі бойынша клетканың үстіңгі бетінде (илазмалеммада) антигенді компонеттер шоғырланады, олар антиденелерге керісінше әсер етеді. Антигендер көмірсулардан, гликолипидтер мен гликопротеидтерден тұрады. Осы глипопротеид мембранасында құрамында активті заттар болатын (гормондар, медиаторлар) әр түрлі рецепторлар орналасады. Жануар организміндегі әр түрлі клеткалар әр түрлі рецепторлардың жиынтығына ие болады, басқаша айтқанда, бір рецепторлардың әр түрлі ссзгіштік қасиеттері болады. Осы рецепторлар арқылы клетканың «күй-жағдайы» туралы хабарлар беріліп отырады.
Қорытып айтқанда, рецепторлар физикалық әсерлерге төтеп беруімен, әрі ерекше заттарды байланыстыруымен қатар клетканың бетіндегі хабарды клетканың ішіне өткізу қызметін де атқарады.
Клетка аралық байланыстар.
Көп клеткалы организмдерді клетка аралық қатынастар байланыстырып тұрады (6-сурет) . Мұны тіпті ұрықтық ұлпалардың клеткаларынан да байқауға болады. Осы ұлпалар сияқты клеткалардың бір-бірімен бірігуін, қосылысуын адгезия деп атайды. Ал осындай қасиет клетканың плазмалеммасының бетінде жатқан химиялық қосылыстардың ерекшеліктерімен де сипатталады.
Сіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz