Іле өзеніндегі тыран (Abramis brama) балығының биологиялық сипаттамасы және жыныс бездерінің гистологиялық құрылымы

Қазақстан Республикасының білім және ғылым министірлігі
әл- Фараби атындағы Қазақ ұлттық университеті
«Қорғауға жіберілді»
зоология және гистология
кафедрасының меңгерушісі
б. ғ. д., профессор Нуртазин С. Т.
ДИПЛОМ ЖҰМЫСЫ
Тақырыбы: « ІЛЕ ӨЗЕНІНДЕГІ ТЫРАН ( ABRAMIS BRAMA ) БАЛЫҒЫНЫҢ БИОЛОГИЯЛЫҚ СИПАТТАМАСЫ ЖӘНЕ ЖЫНЫС БЕЗДЕРІНІҢ ГИСТОЛОГИЯЛЫҚ ЕРЕКШЕЛІКТЕРІ »
050804- «Балық шаруашылығы» мамандығы
Орындаған Алимгазиев. Н
Ғылыми жетекшісі
зоология және гистология
кафедрасының доценті, б. ғ. к., Көбегенова С. С.
Нормабақылаушы Қожабаева Э. Б.
Алматы 2011
РЕФЕРАТ
Дипломдық жұмыстың тақырыбы: Іле өзеніндегі тыран ( Abramis brama ) балығының биологиялық сипаттамасы жене жыныс бездерінің гистологиялық ерекшеліктері.
Дипломдық жұмыс 32 беттен, 2 кестеден, 6 суреттен, 35 әдебиеттер тізімінен тұрады.
Кілттік сөздер: тыран балығы, тұқымдылық, аналық без, аталық без, сперматогониялар, сперматоциттер, оогониялар, ооциттер, көбею жүйесі.
.
КІРІСПЕ
Қазақстандағы өзендердің көбі гидроэнергетикалық жане ирригацалық құрлыстар турғызу натижесінде өзендердің балық шаруашылықтық маңызын түгелге жуық жоғалтты. Казіргі кезде балық аулау өзендердің жоғарғы ағысында немесе ағысы өзгертілмеген өзендерде ғана жүргізіледі. Өзендердің балық щаруашылығындағы маңызы олардың ластануы натижесіндегі жыл сайын төмендеп келеді [1] . Тыран балығы ( Abramis brama Linne ) кәсіптік балық аулау түріне жатады. Оңы Балқаш көліне 1949 жылы жерсіндірілді [2] . Қазіргі кезде тыран Іле-Балқаш бассейінінде кең таралып кәсіпті балықтың қатарына жатады.
Тыран балықтың және басқа кәсіптік маңызы бар немесе жоқ балықтардың сақтау үшін олардың ареалда таралуың, морфо-биологиялық сипаттамаларың, популяционды құрылымың және популяциялардың биологиялық ерекшеліктерін зерттеу керек. Сондықтан біздін жұмыстың мақсаты: Іле өзеніндегі табан балықтың морфо-биологиялық сипаттамасың және көбею жүйесінің гистологиялық құрылысын зерттеу.
Жұмыстың мақсаты: табан балықтың морфо-биологиялық және көбею жүйесінің дамуын және гистологиялық құрылысын зерттеу
Міндеттері:
- Тыран балығының систематикасы, биология, экология, қазіргі сандық көрсеткіштерімен танысу;
- Биологиялық және морфометрикалық анализ жүргізу;
- гистологиялық әдістемелерді пайдалана отырып көбею жүйесінен препарат жасау;
- препараттарға анализ жүргізу және зерттеу нәтижелерін эко-токсикологиялық жағдайымен салыстыру.
- ӘДЕБИЕТТЕРГЕ ШОЛУІле өзенінің жүйесінің физика-географиялық және зерттелетін аудандардың биотоптық сипаттамасы
Іле өзенінің негізгі сипаттамасы және гидрологиялық режимі . Балқаш көлінің су деңгейі Іле өзенінің жоғары ағынының көпшілік бөлігімен бір қалыпты болып келеді: одан барлық көл бассейнің шамамен 82%-ы су ағынымен өтеді. Өзен мұзды тау қорегі және деңгейінің ауытқу маусымымен сипатталады. Су тасқыны көктем-жаз мезгілінде басталса ал межень күз-қыс маусымында белгіленеді: өзеннің орта ағынында 1970 жылы тұрғызылған Қапшағай суқоймасы реттелді [3, 4] . Берілген гидротехникалық құрылым жыл ішіндегі аспекті ағын су бетіндегі буланған сулардың орның толтыруға және өзеннің төменгі ағысы деңгейінің ауытқуына әсерін тигізеді: Іле өзенінің ағын талдауы гидростап бойынша Қапшағай су қоймасынан 164 км жоғары екені көрсетіліп, 2007 жылғы су кірісі орташа шамамен 17, 835 км 3 құраған, сонымен қатар төменгі бъеф бөгетінде орташа шамамен 17, 389 км 3 су жіберілген.
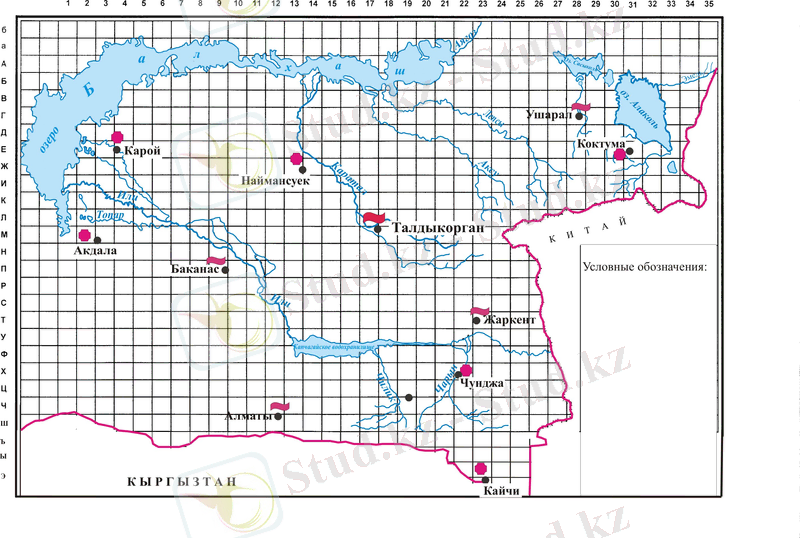
Сурет 1. Іле өзенінің төменгі ағысы
Жоғарыда айтылғандай, 2007 жылғы Қапшағай су қоймасындағы су көлемінің шығысы мен кірісі 1998-2005 жыл аралығындағыға қарағанда аз.
Берілген мәлімет Іле өзен су шығының суаны Қапшағай су қоймасына түскен және шығарылған су көлемі, км 3 [5] .
Іле өзенің ағыны реттегеннен кейін, гидрологиялық режимі төменгі ағысты толығымен көлемі және Қапшағай ГЭС-індегі жылдық су шығынының таралуына байланысты. РГП «Казгидромет» мәліметі бойынша бүгінге дейінгі су шығынының саны (Қаңтар-тамыз, 2008) 9 айдың ішінде шамамен 10, 699км 3 құрады, ал өткен жылғы (былтырғы) жылдың попуси көлемі шамамен 10, 531км 3 болды. Балқаш көлінің 1998-2005 жж. Су көрсеткіші көтерілуі кезеңінде төмен бьер бөгетінің орташа көпжылудың тасталған су көлемі 17, 317 км 3 шамасында болды, ал жылдық попусктар 2006 және 2007 жж. 15, 884 және 15, 164км 3 құрады, осыған солі көтерілу куеңіне қарағанда аз. Бұл фант 2006-2007 жж өзен көрсеткішінің төмендеуіне әкелді. Қапшағай су қоймасындағы попусктік амплитуда ауытқуы Балқаш көлінің көтерілу деңгейінің кезенінде шамамен 6, 023 км 3 құрады, яғни 2002 ж. (21, 003км 3 ) максимум және 2005 ж. минималдық көрсеткішімен (14, 98 км 3 ) . көрсетілгендей, попусктар санының көбеюі көктем айларында (сәуір-мамыр) басталып, жаздың екінші жартысында максималды көрсеткішке жетеді. Берілген жағдай Іле өзенінің ағын реттелуіне дейінгі табиғи жағдай тәртібіндегі кезеңдік ауытқуы сәйкес келіп, кәсіптік балық түрлерінің уылдырық шашуына әсерін тигізеді [5] .
Іле өзенінің гидрохимиялық режимі. Іле өзенімен трансшекаралас сулардың сандық және сапалық құрамы және судың техногенді ластануы, ағын судың мөлшері, Балқаш көлінің экожүйесінің басымдық көрінісі болады. Батыс Балқаш суқоймаларының атырауының сапасының қалыптасуына доминанты әсер ететін Іле өзені, себебі ол Балқаш көлінің бассейінінің негізгі су артериясы. Іле өзені бойынша 2008 жылғы гидрохимиялық сипаттамасы створлармен беріледі: Іле өзені Қапшағай су қоймасынан жоғары, ММ Ұшжарма, Арал - Төбе ауылы және төменгі атырау. Іле өзенінің бастамасынан сағасына дейін баратын жолында судың сапалық құрамында айтарлықтай өзгерістер байқалады. Қапшағай суқоймасының жоғары су минерализациясы жыл мезгіліне байланысты болады, ауытқуы 205-570 мг/дм 3 деңгейінде, ал ортажылдық көрсеткіші 334 мг/дм 3 болады [6, 7] . Бұл нашар минералданған категория суларға жатады, ол иондар қарым - қатынасы бойынша гидрокарбонаттар класы, кальций тобы, ІІ типке. Іле өзенінің суы Қапшағай су қоймасынан шығар жерден төмен ұқсастық су тыздылығын көрсетеді (ауытқуы 275-362 мг/дм 3 ) . Ұшжармадағы МК су минерализациясы 359 мг/дм 3 ( ауытқуы 249 -521 мг/дм 3 ), ал Арал - Төбе ауылының маңайында ол өзгермелі (2006 - 2007 жылдар көрсеткіші бойынша) ауытқуы 392-446 мг/дм З , ал 2008 жылы 442 мг/дм З болды. Құйған ауылының маңайындағы судың тұздылығы 406-446 мг/дм З болды. Іле өзенінің төменгі ағысында және иірім ағысында (протока) ағынды жылы минерализациялануының келесі көрсеткіштер болды 800-883мг/дм З , ол өзеннің төменгі ағысының створларында тұздылықтың жоғарлауын көрсетеді. Доминанты иондардың болуы О. А. Алекиннің [7] класиффикациясы бойынша су ІІ типтің гидрокарбонатты кластың калцийлі тобында жатады. Су минерализациясы жоғарласа ол ІІ типтің гидрокарбонатты - натрийлі класқа көшеді. Тұздар жиыны өзендегі судың барлық ағысы бойынша тұшы екенін көрсетеді
Судың минералдануы жылдар бойынша айтарлықтай емес екенін көп жылдар аспектінде білінеді. Барлық зерттелген жерлерде су ортасының реакциясы - нашар сілтілікті, pH мөлшері 7, 56-8, 10, өзен сағасына қарай ол 8, 25 дейін жоғарлайды.
Газды режимі қанағатанарлық. Қапшағай су қоймасынан төмен суда еріген оттегі мөлшері абсолютті көрсеткіш бойынша ауытқуы 4, 93-11, 1 мг О 2 /дм аралығында, бұл 62-115% қаныққандыққа сәйкес келеді. Судың оттегімен максимальды қанығуы Арал - Төбе ауыл аймағында өктемелі, ол өзен сағасына қарай төмендейді. Атырау ауданында оттегінің төмендеуі саға жағалауындағы аймақтарда өсімдіктердің қопасының кепілдеуімен түсіндіруге болады.
Фотосинтез процессіне және су ортасының реакцисына байланысты көмірқышқылы болуы бірқалыпты емес және оның концентрациясы тек З, 20-7, 04мг/дм З . Іле өзенінің төменгі ағысында органикалық заттардың болуы перманганатты қышқылдану көлеміне қарай жоғары емес, ортаңғы ағысында мөлшері 2, 7-4, 5мгО/дм З және төменгі ағысында 4, 5-7, 5 мг О/дм З аралығында. Көп жылдық бақылаудағы динамика кәзіргі уақытта сақталған.
Өзен створларындағы биогенды элементтердің концентрацисы келесі түрмен реттелген: аммонинийлі азот және темір ауытқуы 0, 01-0, 13 мг/дм З , нитритті азот және фосфор 0, 003 тен 0, 064мг/дм З дейін, нитраттар 0, 16 дан 4, 10 мг/дм З дейін. Кремний саны 2, 8-3, 9 мг/дм З аралығында. Алынған материалдар бойынша уақыт және кеңістік салыстырмасы бойынша ең вариабельді нитраттар, нитриттер, фосфор болуып табылады, ол өзеннің су мөлшерінен сонымен қатар суғарылған егістік жерлерді суғарғаннан ластанып қайтарылған сулардың өзенге келіп құйылуына байланысты болады [7] .
Іле өзенінің сапалық көрсеткіштері судың тасу мезгілінде және оның деңгейінің төмендеуінің басталуы балықшаруашылығына маңызы бар су қоймаларға ұсынған Мемлекеттік стандарт бойынша нормативті талаптарға сәйкес келеді. Створлардың төменгі ағысы бойынша биогенді элементтердің уақытша көбеюі антропогенді әрекеттерінің болуы, ол реттелмеген ластанған ағынсулардың ауылшаруашылық
егістіктерінен келуіне байланысты [6, 7, 8] .
1. 2 Іле өзенінің жүйесінде тыран балығының пайда болуы
Қазақстанда тыран балық Каспий және Арал бассейінің аборигендері болып табылады. 1949 жылы оны Бақаш-Іле және басқа бассейндеріне жерсіңдірілген [2] . Тыран балығы - ерекше ірі балық, салмағы 4-5 килограмм, ұзындығы 30-35 сантиметр, інішара салмағы 10 кг, ұзындығы 75 см болатын балықтар кездеседі. Тыран балықтар 20 жыл өмір сүретін балық. Днепр жағалауында табан балықтар тез өсіп өнеді, ал солтүстік су қоймаларында баяу өседі. Аналықтары аталықтарынавн ірілеу болып келеді. Табанның дене бітімі биік, бүйірі қысыңқы, басы кішірек ауыз қуысы трубка тәрізді, төмен орналасқан. Балық түсі жасына байланысты, топырақ және су қоймасындағы суға байланысты өзгеріп отырады. Әдеттегі түрелері жасына байланысты алтын, сұр түстен басталып, кейіннен қою алтын сары түсті болып анық көріне бастайды. Табан балығы өмірінің көп уақытын көлдің тұздалған қабатында өткізеді [9] .
Еуразияда табан балығының Abramis cuvier туысының 4 түрі кездеседі, оның ішінде Қазақстанда 3 түрі: Олар Abramis brama (linne), Abramis sapa (Ballas) және Abramis ballerus (linne ) . Олардың сипатталуы ұзын анальді жүзгіші мен бір қатарлы 5-5, 6-5, 5-6 және қиғаш орналасқан тұтқыншақ тістерімен айқындалады. (Никольский, 1980) . Бірқатар ғалымдардың зерттеулерінше олардың басқа табандардан өзгешілігі басының және арқа жүзгішінің биіктігі. Тыран балықтарының көптеген морфалогиялық зерттеуіне қарағанда олар белгілі географиялық өзгеріске ұшыраған. Қапшағай су қоймасындағы табандар зерттеулерінің нәтижесінде қарағанда ұқсастықтары Арал табандарына келеді [9, 10, 11, 12] .
Қазігі уақытта дейін Abramis blicca және bimba туыстық ара қатынасы толығырақ дәлелденген. Ихтиологиялық әдебиеттерге онда уақыт бойы blicca bjoerkna балықтары екі қатарлы тіс жұтқыншағы және аузы монотипті деп жазылып келген. Кейінгі деректерге сүйенетін болсақ осы балықтардың ұқсастығы қайта қаралған олар: B bjoerkna және Abramis табан балығының үш түрі Abramis brama, Abramis ballerus және ақ көз Abramis sapa. Осы табан балықтарын жақындастыратын ерекше белгі: арасында бір қатарлы және екі қатарлы тісті табандар кездеседі. Ал Abramis ballerus пен ақ көз балықтарының тістерінің екінші қатарлы пайда болу жайында әдебиеттерге дерек жоқ. Автордың қысқаша дерегіне қарасақ, Abramis brama, blicca, bimba балықтарының бір-біріне туыстығына қарама-қайшы жайттар туындайды, әлі де бұл материалдар терең зерттелуі тиіс [11, 13] .
Abramis sapa - ақ көз балыққа тоқталатын болсақ, басқа тыран балықтарына қарағанда ауыз қабаты қалың, ауыз төмендеу орналасқан және басқа табан балықарына қарағанда тығыз су қабатына үйренген. Табандарға қарағанда дене бітімі кішіректеу, көздері үлкен. Бұл табан балықты қара, Азов, Арал, Каспий теңіздерінен және Дунай, Дон, Днестр, Днепр, Волга, Урал, Терек өзендерінен кездестіруге болады. Ал Қазақстанда жүйірек кездесетін екі түрі бар: кәдімгі Abramis sapa aralensis Tiapkin. Abramis sapa sapa pallasтаралуы Урал өзенінің барлық жақтарында және каспий теңізінде кездеседі, ал Ембі, Сағыз өзендерінде кездеспейді. Арқа жағынан түсі қоюлау болып келеді, ал бүйірі күміс түстес болып келеді. Жүзгіштерінің жұпсызының ұштары қою түспен көнкерілген, ал жұпты жүзгіштерінің түсі ақ сары болып келеді. Ұзындығы 30 см-ге дейін жетеді, салмағы 800 г. Солтүстік Каспийде 33 см-ге дейін, толығырақ айтқанда басым көпшілік табанның ұзындығы 17-18 см, ал Орал өзенінде 20-24 см, салмағы 170-280 г. Оралдың басқа өзендерінде Меркулованың деректеріне қарағанда, ұзындығы 29, 5см-ге дейін болатын тәрізді [11, 14] .
- Тыран балықтың систематикасы және жалпы сипаттамасы
Тип: Chordata - Хордалылар
Тип тармағы: Vertebrata (Craniota) - Омыртқалылар (Бас қаңқалылар)
Бөлім: Gnathostomata - Жақтылар
Класс үсті: Pisces - Балықтар
Класс: Ostichthyes - Сүйекті балықтар
Класс тармағы: Actinopterygii - Сәуле қанаттылар
Отряды Cypriniformes - Тұқытәрізділер
Тұқымдасы Cyprinidae -тұқылар.
Туысы Abramis Cuvier 1817- Тыран
Түрі 1. Abramis brama (Linne),
Түрше Abramis brama orientalis Berg - шығыс табан балығы [9, 11] .
Туысы Abramis Cuvier 1817- Табан Л. С. Берг [10] бойынша, Евразияда бұл балықтың 4 туысы мекен етсе олардың 3 түрі Қазақстанда кездеседі: Abramis brama (Linne), Abramis sapa (Pallas) және Abramis ballerus (Linne) . Олар аналь жүзбеқанатының ұзындығымен (біздің түрде сәулелерінің саны 22-ден кем емес) және бірқатарлы қиғаш кесілген 5-5, 6-5, 5-6 жұтқыншақ тістерімен сипатталады. Понто- каспийлік тұщысу комплексіне жатады (Никольский) .
Abramis brama (Linne) -табан. Қабылданған түрлердің бөлінуін әдеттегі түрше Abramis brama, brama және барлық орталық Солтүстік Еуропада мекен ететін (Солтүстіктегі, Балтикадағы, Қара және Ақ теңіз бассейндерінде) түрше Abramis brama orientalis (Каспий және Арал теңіздері бассейндерінде) деп бөледі.
Л. С. Берг [10] бойынша бұл екі түршенің айырмашылығы негізінен бірінші желбезек доғасындағы желбезек қылтандарынан, омыртқа және бүйір сызығындағы қабыршақтарынан көптеген зерттеушілер аналь қанатының сәулелері санынан, құйрық қанатының қалақшасынан, арқа қанатының ұзындығынан, антедорсальды қашықтығынан және басының биіктігінен айырмашылықтарын белгілеп берді.
Тыран балықтың сандық белгілерінің ауытқуын толығымен типтік форманың ауытқуына жатқызады.
Тыран балығында диплоидты хромосома жиынтығының 2n=50, олардың ішінен 12 мета - және субметацентрикалық сонымен қатар 38 субтелоцентрикалық және акроцентрикалық бар [15] .
Таралуы. Понто- каспийлік фаунистикалық комплекске жатқызады.
Қазақстанда тыран балығы Каспий және Арал теңіз бассейндері суқоймаларында аборигенді болып табылады [2, 9, 10] .
Табанды орналастырудың нәтижесі. Балхаш -Іле бассейнінде 1949 ж, Зайсан көліне, Бұхтырма және Өскемен суқойма бассейніне, Ертіс өзеніне 1959-1964 жж., Талас өзені көл бассейніне 1958 ж., Көкшетау көліне 1959-1965 жж. Солтүстік Қазақстанға 1962-1966 жж және Ақмола облыстарына 1965-1966 жж отырғызылған. Қазіргі кезде бұл балық республикамыздың негізгі балықөндірістік суқоймаларында таралған [2, 9, 16] .
Сипаттамасы. D III 9-12, көбінде 9-10; А ІІІ 22-30, көбінде 24-26; бүйір сызығындағы қабыршақтар саны 47-58; сыртқы жағындағы бірінші желбезек доғаларындағы желбезек талшықтарының саны 19-32, көбінде 22-28; омыртқалар саны 38-46, көбінде 41-44; жұтқыншақ тістерінің формуласы 5-5 кейбір жағдайда 6-5 немесе 5-6. Антедорсальды қашықтығы дене ұзындығынан пайызбен есептегенде (С-сыз) 50, 2-64, 5 құрайды, постдорсальды- 28, 6-40, 7, пектовентральды-17, 5-27, 0, вентральды -13, 6-26, 5, денесінің ең биік жері 30, 0-44, 6, денесінің аласа жері 7, 4-7-13, 5, құйрық қанатының ұзындығы 7, 3-18, 1, кеуде жүзгіш қанатының ұзындығы 16, 3-28, 9, құрсақ қанатының ұзындығы-13, 6-26, 5; арқа қанатының негізгі ұзындығы 9, 9-19, 8, оныңбиіктігі 19, 8-34, 9; аналь қанатының ұзындығы 19, 3-33, 1; оның биіктігі 13, 8-24, 7; басының ұзындығы 16, 0-27, 6, оның биіктігі-14, 5-24, 6; тұмсық ұзындығы 4, 2-9, 0, көз диаметрі 3, 6-7, 4, көз арты бөлігі 8, 6-14, 2, қабағының ені 4, 2-10, 0 [9] .
Денесінің түсі күміс түсті, үлкен особьтарында қола секілді арқасы неғұрлым қою, ал құрсағы ашықтау боялған. Денесі биік, бүйірінен өте қатты сығылған.
Аналық және аталық табан балықтарының сыртқы айырмашылығы уылдырық шашар алдында және уылдырық шашу кезінде ғана ажырытылады. Аналықтарының құрсағы үлкен, жыныс тесігі ісінген және уылдырық шашқаннан кейін қызыл түсті болып келеді. Аталықтары көбейер алдында- кеудесі мен басының дорсальды және латеральды жақтауларында маржан бүршіктері пайда болады, жүзгіш қанаттары кедір бұдырлы болып келеді. Тыран балықтарының жыныстық деморфизмі әлсіз ажыратылған, әртүрлі белгі әртүрлі суқоймаларда. Сондай-ақ Балхаш көліндегі жыныстық ауытқуы бүйір сызығындағы қабыршақтарының, Р-ұзындығының (көбінесе аналық балықтарда) және көзінің диаметрі [17, 18] .
V-ұзындығы көбінесе аталық балықтарда бойынша белгіленген. Х. К. Исмұхановтың берілгендері бойынша Бұқтырма суқоймасындағы табан балығының аталықтары аналығынан үлкендеу, вентроанальды арақашықтығынан және бастың көз арты бөлігінен сонымен қатар басының аз биіктігімен ерекшеленеді. 70-ші жылдардың басында Қапшағай суқоймасында табан балығының жыныстық деморфизмі постдорсалды арақашықтығынан және Р-ұзындығынан болатыны көбірек аналық балықтарда кездескені белгіленген [16, 17] .
1978 ж таңдамалар бойынша жыныстық деморфизмнің 4 нақты белгілері байқалған: аналықтарында үлкен көзінің диаметрі, бірақ көзарты бөлігі кіші, V ұзындығы және С үстіңгі сабақшасының ұзындығы. Арал теңізі және Орал өзенінде мекендейтін тыраннын аналығында желбезек талшықтарының саны көп болған аталықтарына қарағанда. Осындай кейіпте, тыран балығының әр түрлі суқоймаларда, әртүрлі уақытта морфологиялық белгілері жыныстық деморфизмінің нақты ауытқуларын анықтамайды [17] .
- Тыран балықтың биологиялық ерекшеліктері
Көбеюі. Тыран балығы жылына бір-ақ рет ұрық шашады және олар порциялап ұрық шашатын популяцияға жатады. Жалпы заңдылық бойынша оның порциясының көбеюі солтүстіктен оңтүстікке қарай шашуы бақыланады. Қазақстанның суқоймаларында табан балығы порциялап ұрық шашпайды. Көптеген суқоймаларда тыран балығының уылдырық шашу кезеңі сәуір айының соңымен мамыр айының басына дейін болады, судың температурасы 11-14 С-тан төмен болмай тұрған кезде. Бақылау бойынша Қапшағай суқоймасында 1979 жылы бірінші «өтпелі» аналықтары сәуірдің 15-16, судың температурасы 11-12 С, ал сәуірдің аяғында судың жылуы 17-19 С балық аулығанда шұңқыр жерлерде табан балықтың аналықтары көптеп кездескен. Г. М. Дукровецтің бақылауы бойынша Раим көлімен Сырдария өзенінде 1973 жылы бірінші порциялап ұрық шашу 10 тәулік бойы жүрген 7-17 мамыр аралығында. Барлық уылдырық шашу кезеңі әртүрлі суқоймаларда 1 айдан 2, 3 айға дейін ауытқиды. Қапшағай суқоймасында табан 1971-1972 жылдарда уылдырық шашудың екі типі, 1973-1974 жылдары үш порциялық типтері болған. 1979 жылғы мәліметтерге қарағанда және одан кейінгі жылдарда да жетіліп шыққан аналықтар бір уақытта уылдырық шашады, ал қайталанып уылдырық шашатын аналықтары екі порциялап және аздаған бөлігі үш порциялап шашатын типке жатады. Уылдырық мөлшері өзгерген жағдайда, сонымен қатар кезекті уылдырық шашу кезеңінде ерте жетілетін особьтары кейбір жылдары, стадиялардың көбейіп кетуі олардың қорекпен қамтамасыз ету базасына байланысты болған. Осыған ұқсас Арал теңізінің табан балықтарының заңдылығын өмір сүру жағдайының тез нашарлағанын байқаған. Кейбір зерттеушілер уралдық балық аулау жағдайына үйірленіп, порциялап уылдырық шашуын жоғалтқанын ұйғарды. Табанның жоғары экологиялық икемділігі уылдырық шашу бір суқоймада жартылай өткінші кездеседі. Табан балығы суқойманың жағдайына тез бейімделіп кетеді. Табан балықтың уылдырық шашу қабылеті әртүрлі, кез-келген суқойманың типіне бейімделген (өзен, көл, суқойма, теңіз және т. б. ), 1, 5-3 м. Уылдырықтың дамуы, яғни эмбрионнан жарып шығуы 3-тен 7 тәулікке дейін жүреді. Эмбрионнан личинкалардың жарып шыққан ұзындығы 5, 6-6, 4 мм. Личинкалық кезең 30-35 күн өтеді. F этапында ұзындығы 24 мм. Табан балығының абсолюттік тұқымдылығы әртүрлі әрқалай, ол оның қорекпен қамтамасыз етілуі жетілуі және өмір сүретін жағдайына байланысты. Арал тыран балығының тұқымдылығы 60 жылдары біртіндеп төмендей бастады. Жоғарғы орташа 1960 жылы 235, 8 мың уылдырық болған. 1968 жылы тұқымдылығы 21, 4-96, 6 мыңға төмендеп кетті. Арал табан балығының ең төменгі массасы (0, 35-0, 45 кг) басқа жылдармен салыстырғанда. Сырдария өзенінде 70 жылдары ол кездері жоғары болған жоқ орта есеппен 1 уылдырық болған. Бұқтырма табан балығының абсолюттік тұқымдылығы 1975 жылы (198, 8 мың уылдырық болған) 1974 жылмен салыстырғанда 154, 0 мыңға жеткен. Балхаш көліндегі тыран балығының тұқымдылығы заңдылық бойынша массасы, денесінің ұзындығы және жасы жоғарылайды. Қапшағай суқоймасының табан балығында да дәл осындай жағдай кездеседі [17, 19] .
... жалғасыСіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz