Сұлының тәжтәріздес тат ауруына төзімділігінің генетикасы

ҚАЗАҚСТАН РЕСПУБЛИКАСЫНЫҢ БІЛІМ ЖӘНЕ
ҒЫЛЫМ МИНИСТРЛІГІ
Жаратылыстану факультеті
«Жалпы биология және экология» кафедрасы
Дипломдық жұмыс
Тақырыбы: «Сұлының тәжтәріздес тат ауруына қарсы төзімділік генетикасы»
Орындаған:
Тобы :
Жетекешісі:
Кафедра меңгерушісі:
Жұмыстың мақсаты мен міндеті
- Сұлының тәжтәріздес ауруына қарсы тиімді көздерін анықтау.
- ЖЖЖергілікті немесе аудандастырылған сұлы сорттарының төзімділігін арттыру және жаңа өнімді сорттар шығару.
Ғылыми жаңалығы:
- Жергілікті сорттардың төзімділігі популяциядағы вирулентті нәсілдердің үлесіне байланысты екені анықталды.
- Жергілікті немесе аудандастырылған сұлы сорттарының төзімділігін арттыру үшін жаңа өнімді сорттар шығару үшін коллекциялық төзімділік көздерін пайдалану тиімді екені анықталды.
Практикалық маңызы: Өсімдік қорғаудың ең тиімді және экологиялық залалсыз тәсілі төзімділік селекциясы. Зиянды ағзаларға төзімді сортты пайдалану қарқынды дамып келе жатқан ауыл шаруашылығындағы өндірісіндегі өнімділік деңгейін көтеруге ықпалын тигізетіні күмәнсіз.
АННОТАЦИЯ
Дипломдық жұмыс «Сұлының тәжтәріздес тат ауруына қарсы төзімділік генетикасы» тақырыбында орындалған. Жұмыстың жалпы көлемі 46 бет.
Жұмыстың мазмұны кіріспе, негізгі бөлім, қорытынды, ұсыныстар, жұмыстың экономикалық тиімділігі, еңбек қорғау, қоршаған ортаны қорғау бөлімдерінен тұрады.
Соңында 22 тұратын пайдаланылған әдебиеттер тізімі келтірілген. Дипломдық жұмыс 6 суреттермен көркемделген. Эксперименттік деректер 5 кестелерде көрсетілген.
АНЫҚТАМАЛАР
- Тат ауруы- дәнді дақылдарды залалдайтын, базидиомицеттер класына жататын микроскопиялық саңырауқұлақтар тудыратын ауру түрі.
- Патоген- ауру тудыратын ағза.
- Вируленттік- патогеннің популяциясының өсімдікті залалдау қабілеттігі.
- Авируленттік-патогеніің өсімдікті залалдау қабілетінің болмауы.
- Төзімділік гені- өсімдіктің патогеннің әсеріне қарсы тұратын гені.
- Дифференциация- популяцияны жеке физиологиялық нәсілдерге бөлу.
- Жіктеуіш сорттар- патогеннің популяциясын жеке нәсілдерге бөле алатын, әртүрлі төзімділік гендері бар сорттардың жиынтығы.
- Ювенилдік- өсімдіктің вегетациясының алғашқы фазалары.
ҚЫСҚАРТУЛАР
- АҚШ- Америка Құрама Штаттары.
- КСРО-Кеңес Социалисттік Республикалар Одағы.
- ТМД-Тәуелсіз Мемлекеттер Достастығы.
- ФАР-Фотосинтетикалық белсенді радиация.
- Рс-Puccinia coronata.
- Мг-миллиграмм.
- Кг-келі.
МАЗМҰНЫ
Аннотация . . .
Анықтамалар . . .
Қысқартулар . . .
Кіріспе . . .
1. Әдебиетке шолу . . .
1. 1 Сұлының шығу тегі мен селекциясының тарихы . . .
1. 2 Сұлының биохимиялық және дақылдық ерекшеліктері . . .
1. 3 Басты түрлері мен түрішілік топтары . . .
1. 4 Морфологиялық белгілері мен биологиялық ерекшеліктері . . .
1. 5 Физиологиялық ерекшеліктері . . .
1. 6 Тәжтәріздес тат ауруының биологиялық ерекшеліктері . . .
1. 7 Сұлының тәжтәріздес тат аурунына қарсы тиімді күрес жолдары . . .
2. Негізгі бөлім . . .
2. 1 Сұлының тат ауруына төзімділігін бағалау әдістемесі . . .
2. 2 Экспериментке пайдаланылған материалдар . . .
2. 3 Эксперименттің нәтижелері . . .
3. Қорытынды . . .
4. Ұсыныстар . . .
5. Дәнді дақылдардың төзімділік селекциясының экономикалық тиімді . . .
6. Еңбек қорғау қағидалары . . .
7. Қоршаған ортаны қорғау . . .
8. Пайдаланылған әдебиеттер тізімі . . .
КІРІСПЕ
Сұлы дәнді дақылдардың бірінші тобына жатады. Ол бидай, арпа және қара бидай сияқты дақылдармен қатар аса маңызды болып табылады. Әлемдегі ауылшаруашылық мәдени өсімдіктердің егістік көлемінің (980 млн. га) 75% осы өсімдіктердің үлесінде. Өткен ғасырдың аяғындағы статистикалық деректерге жүгінсек, бұл дақылдар әлемде 725 млн. га, ал кеңес одағы елдерінде 138, 8 млн. га жерге себілген. Сұлы, егістік көлемі жағынан, бұл өсімдіктердің ішінде төртінші орын алады. Құндылығы жағынан басқа дәнді дақылдардан бағалы. Сұлының құрамында валин, лизин және триптофан сияқты алмастырылмайтын амин қышқылдары көп мөлшерде болады. Бұл қасиеттері сұлыны құнды диеталық тағам ретінде пайдалануға негіз болды. Қабықсыз дәнді сұлы диеталық күлше және, өзімізге белгілі, «Геркулес» ботқасын жасауға пайдаланылады. Сұлыдан ас қазанды емдейтін кілегейлі сусын даярлайды.
Мал шаруашылығында сұлының дәні өте пайдалы азық. . Сұлы дәні мал азығының құндылығының халықаралық өлшем бірлігі болып саналады. Басқа дақылдардың дәндерімен араластырылған сұлы жас төлдердің ағзасының тез және мықты болып қалыптасуына жағдай жасайды. Сұлы қосылған концентрлі азық құс шаруашылығында кеңінен қолданылады. Сұлының сабаны басқа дақылдардың сабанына қарағанда қоректік заттарға бай және малға жағымды болады. Сонымен қатар, оның ылғал сіңіргіштік қасиетіде жоғары болғандықтан ірі қара және шошқа шаруашылығында төсеніш ретінде пайдаланылады.
Сұлы басқа дәнді дақылдарға қарағанда мәдени өсімдіктер қатарына кейін ендірілген. Себістік көлемінің кемдігінің де бір себебі осыдан. Дегенмен, сұлы өнімін оның себістік көлемін ұлғайту арқылы емес, гектардан алынатын өнімнің түсімін жетік технологияларды пайдалану арқылы арттыру барлық елдерде көзделіп отыр. Бұл мақсатқа сұлы өсімдігіне қажетті физиологиялық қалыпты жағдайларды жасау тәсілдерін меңгеру арқылы ғана қол жеткізуге болады. Өсімдікті жерге еккеннен бастап оны жинауға дейінгі созылатын вегетациялық кезеңде барлық агротехникалық талаптар толығымен орындалуы тиіс. Өнімнің түсім мөлшері сұлының ауыспалы егіс жүйесіндегі орны, топырақ өңдеудің сапасы, минералды тыңайтқыштардың түрлері мен мөлшері, оларды топыраққа ендіру тәсілдері мен себу мерзімі, өскіндерді зиянкестер мен аурулардан қорғау сияқты маңызды технологиялық элементтерді деркезілік және сапалы орындаудан тәуелді. Барлық аты аталған агротехникалық тәсілдер деркезілік атқарылуы керек. Ауылшаруашылығының қарқындылығын арттыру жолында көптеген мәселелер туып отыр. Соңғы жылдары тек өнімнің түсімін жоғарылату мақсатында жүргізілген технологиялық енгізулер көптеген экологиялық мәселелерді тудыратынын көрсетті. Бұған өсімдік шауашылығындағы арам шөптерге, зиянкестерге және ауруларға қарсы қолданылған химиялық қорғаныс тәсілдеріндегі кемшіліктерді және топырақ құнарын сақтау мақсатындағы іс-шаралардың жетіспеушіліктерін мысал етуге болады.
Аталған проблемаларға орай, сұлының өнімділігін арттыру жолында перспективті тәсіл болып төзімділік селекциясы саналады. Селекциялық жұмыстың қарқындылығы патогеннің популяциялық құрамын дифференциациялау арқылы тиімді гендерді анықтауға тікелей байланысты.
1. ӘДЕБИЕТКЕ ШОЛУ
1. 1 Сұлының шығу тегі мен селекциясының тарихы
Сұлыны дақылдандырудың көпғасырлық тарихы бар. Азия елдерінде көне заманнан бастап сұлының құнды қасиетін білген. Сұлыны егістікте дақыл ретінде егіп өнім жинаған. Осындай көпғасырлық тұқым шығару жұмыстарының нәтижесінде сұлының жергілікті сорттары қалыптасқан. Дегенмен, сұлыны зерттеудің қарқынды кезеңі ретінде өткен ғасырдың екінші жартысын бөліп айтуға болады. Онтоғызыншы ғасырдың екінші және жиырмасыншы ғасырдың бірінші жартысында Ресейде өсімдікшаруашылығының ғылыми негізі қалана бастады. Орыс ғалымдары И. А. Стебут (1833-1923), К. А. Тимирязев (1843-1920), Д. Н. Прянишников (1865-1948) өсімдік шаруашылығының физиологиялық, агрохимиялық, биохимиялық негіздерін қалады. К. А. Тимирязев өсімдіктердің өнімділігін едәуір көтерудің ғылыми негізін ашты. Ол өсімдік өнімінің басты өмірлік факторлар арқылы реттелетінін көрсетті. Олар, ғылыми еңбектерден басқа, тәжірибелік іс бастамалар арқылы өсімдік шаруашылығын дамытты. Бұл жылдары Мәскеу ауылшаруашылық академиясы және көптеген өсімдік шаруашылығы тәжірибелік бекеттері ашылды. Осы кезеңде (1910), Қазақстандағы ең бірінші, Қызыл сарқырама (Красноводопадская опытная станция) ғылыми-зерттеу, селекциялы - тәжірибелік бекеті ашылды. В. Г. Хржановский, с. ф. Пономаренко. 1982. (1)
Бұрынғы ССРО да дәнді дақылдардың селекциясы өте қарқынды жүргізілді десек артық болмайды. Бұл жылдары ел көлемінде сорттарды сынау жұмыстары жүргізіліп олардың ішіндегі ең бейімділері белгілі аудандарға аудандастырылып отырды. Селекциялық жұмыстар Бүкілодақтық өсімдіктану ғылыми-зерттеу институтының әлемдік коллекциясын пайдалану арқылы жүргізілді. Аталған коллекция орыстың ұлы ғалымы - генетик, селекционер, Н. И. Вавиловтың орасан зор еңбегімен жиналған. Қазіргі замандағы аудандастырылған сорттардың ішінде бұл коллекцияның қатысуынсыз шығарылған сорттар кемде-кем.
Қазақстанда дән шаруашылығының қарқынды дамуы жиырмасыншы ғасырдың екінші жартысынан басталды. Қазақстанның ССРО бойынша дән алқабы болып саналуына байланысты, Бүкілодақтық дән шаруашылығы ғылыми-зерттеу институты тың астанасы - Целиноград облысында орналасты. Алматыда егістіктану, өсімдік қорғау және өсімдік физиологиясы ғылыми-зерттеу институттары өз жұмысын бастады. Қазақстандық ғалымдардың өсімдікшаруашылығының дамуына қосқан үлесі орасан зор. Тың жерді меңгерудің технологиялық негізін қалаған ғалым академиктер -А. С. Сүлейменов және А. И. Бараевтың еңбектері өсімдікшаруашылығы ғылымының классикалық тұрғысына айналды. Қазіргі кезде Қазақстанда аудандастырылған сорттардың басым көпшілігі жергілікті ғылыми-зерттеу мекемелерінде шығарылған.
Сұлы ылғал сүйгіш және салқын климатты жақсы көретін өсімдік. Қазақстанның сұлы себетін аймақтары болып солтүстік облыстар мен оңтүстік облыстарының таулы аудандары саналады. Қазақстанда бұл дақылдың егістік көлемі қазірге айтарлықтай көп емес. Дегенмен, жоғарыда келтірілген құнды қасиеттеріне байланысты бұл өсімдік өз сұранысын табады деген сенімділік бар.
Сұлының өнімін арттырудағы атап айтар резервтер болып өнімділікке бағытталған селекциялық жұмыстар, жоғары өнімді сорттарды қолдану және өнім өндірудің алдыңғы жетік технологиясын пайдалану саналады. Жукова А. Э 1985 (2)
1. 2 Сұлының биохимиялық құрамы мен дақылдық ерекшеліктері
Сұлының таралуы . Сұлы өнімінің басым бөлігі мал азығы ретінде қолданылады. Сұлы дәнінің құрамында 11, 4-13% дейін белок, 40-45% крахмал және 4-5% май болады. Сұлы дәні жылқыға және басқа түліктердің төлдері үшін өте құнды концентрлі азық болып саналады (1-кесте) .
Сұлыны танаптық егістікте ас бұршағымен араластырып сүрлем, пішен және көк балауса өндіруде қолданады. Орта белдеулерде және солтүстік ендікке жақын аудандарда аталған қоспаны бұршақ тұқымдас көпжылдық шөптерді еккенде, бірінші жылы жабындық өсімдік ретінде қолданады. Жабындық өсімдікті ерте, жаз ортасында, көк балаусалық азық ретінде немесе сүрлем даярлауға орып алады. Н. Мухитдинов, Ә. Бегенов 1993 (3)
1-кесте
Сұлы пішені мен көк балаусасының азықтық құндылығы
азық. бірлік
қорытлкгы. прот. (%)
протеин
белок
май
клетчатка
АЭЗ
Көк балауса
Сұлы пішені
16, 8
47, 7
2, 5
6, 2
149
130
2, 5
6, 2
1, 8
3, 2
0, 7
1, 5
3, 7
14, 5
5, 8
23, 8
Сұлы жабындық өсімдік ретінде кеңінен қолданылады. Көпжылдық бұршақ тұқымдас шөптер вегетациясының бастапқы кезеңдерінде арам шөптерге қарсылық көрсете алмайды. Сондықтан, танапты арамшөп қаптап кетпес үшін оларды сұлымен аралстырып себеді. Бұл тәсілдің екінші пайдасы - ол танапты тиімді пайдалану. Бұл кезде жабындық өсімдіктің себіс мөлшері 20кемітіледі. Өйткені, өте қалың өсімдік тұрғысы күн сәулесінің мөлшерін өте азайтып басты өсімдіктердің жақсы дамып өсуіне кедергі туғызуы мүмкін. ауыспалы егістік жүйесінде сұлыны демалуға арналған танаптарда (жасыл парда) өсіреді.
Осы себептерге орай сұлы өсімдігі, көбінесе, мал шаруашылығы дамыған аймақтарда кеңінен таралған. Жер бетінде жаздық сұлы көп таралған. Өткен ғасырдың екінші жартысында әлемдегі сұлының егістік көлемі 35 млн жеткен. Сұлыны көп егетін елдер қатарына АҚШ (9, 2 млн га), Канада (4, 1 млн га), Франция (1, 31 млн. га), Польша (1, 8 млн. га) және Жаңа Зеландия (0, 9 млн. га) жатады. Бартоне Г, Шебеста И 1984 (4)
ССРО да жаздық сұлының егістік көлемі 9 млн жуық болған. Ресейде сұлы егістігінің басым бөлігі сұртопырақты зона үлесіне тиеді. Солтүстік - Батыс экономикалық ауданының егістік көлемінің 40% сұлы өсімдігі қамтиды. Сонымен қатар, қиыр шығыста. волга бойында, батыс облыстарында және орал таулы аудандарында сұлы айтарлықтай көлемде егіледі. Украинада орманды және орманды- дала аймақтарында сұлы кеңінен таралған. Кеңес өкіметі кезеінде сұлының егістік көлемі, барлық дәнді дақылдардың егістік көлеміне шаққанда, 43% болатын. Бұл жиналатын өнімнің 31% құраған. Ал Ресей федерациясның егістігінде сұлы әлі күнге дейін үлкен үлес алады. Солтүстік - батыс облыстарында Еділ жағалауындағы аудандарда оның егістік көлемі 40% асады.
Сұлының күздік пішіндері Қазақстанда және орта Азия мен Закавказье мемлекеттерінің таулы аймақтарында егіледі.
Сұлы дән түсімі жағынан басқа дәнді дақылдардан айтарлықтай төмен емес. Ресей мемлекетінің солтүстік батыс және солтүстік шығыс ендіктік аймақтарында өнім түсімі 1985-1990 жылдары, орта есеппен алғанда, бір гектардан 22, 3 -25, 8 центнер көлемінде болған. Қазақстаннның оңтүстік аудандарында бұл көрсеткіш сәл төмендеу, 16, 5 - 17, 8 центнер. Жоғарыда айтылғандай, оңтүстік аймақтарда көбіне күздік сұлы қолданылады. Чупцова О. Е 1981 (5)
1. 3 Басты түрлері мен түрішілік топтары
Сұлы туысына 70 астам түрлер кіреді. Олар мәдени және жабайы пішіндер. Хромосомалар саны жағынан барлық сұлы түрлері үш топқа бөлінеді: диплоиды түрлерінің соматикалық жасушаларында 14, тетраплоидты түрлерінде 28 және гексаплоидты түрлерінде 42 хромосомалар болады.
Сұлының төрт түрі, басқа түрлеріне қарағанда, кеңінен таралған: егістік сұлы- барлық ауылшауашылықтық аймақтарда кездеседі; византиялық сұлы жерортатеңізінің жағауындағы елдерде, АҚШ та, Аргентинада, Уругвайда, Австралияда, Жаңа Зеландияда және Африка континентінің оңтүстігінде кеңінен таралған; құмыдық сұлы Испанияда, Португалияда, Францияда және Англияда дақылданған; абиссиналық сұлы Эфиопияда және Йеменде қолданылады. Жемчужина А. И (6)
ТМД елдерінде және Қазақстанда сұлының егістік және византиялық түрлері ғана дақыл ретінде пайдаланылады.
Ауылшаруашылығы өндірісінде пайдаланылатын сұлы сорттарының басым көпшілігі осы түрлерге жатады. Бұл түрлердің хромосомалар жиынтығы бірдей және өнім түсімі де жоғары. Сондықтан, бұл түрлерді селекционерлер өз жұмыстарында жиі пайдаланады. Ал, жабайы түрлері басқа дақылдардың егістігінде, өте қиын ажыратылатын арам шөп ретінде кездеседі.
Түрлерді өзара ажыратуда келесі белгілерді есепке алады:
- сыртқы гүл қабықшасының төбесінде қылтық тәріздес өскіннің болу болмауы;
- дән астында таға тәріздес құрылымның болу болмауы;
- сұлы піскендегі масақшаның үгітілу ерекшеліктері.
Сұлының кейбір түрлерінде дән астында ерекше таға тәріздес бөлшек болады. Бұл бөлшек дән пісіп жетілгеннен кейін оның масақшадан бөлініп үгітіліп түсуіне жағдай жасайды. Сондықтан, мұндай бөлшегі бар түрлердің дәндері піскен кезде жеке-дара дәндерге бөлініп үгітіліп кетеді. Жабайы сүле осылай таралады. Ал, оңтүстіктік овсюг түрінде бұл тағатәріздес бөлшек тек масақшаның төменгі дәнінде ғана болғандықтан, пісіп жетілген дәндер бүтін масақшалар түрінде үгітіліп түседі. Сұлының жабайы түрлерінің гүл қабықшалары түктенген және ұштары қылтық тәріздес өскінді болады, әсіресе оңтүстік овсюг түрінде (1-2 -суреттер) .
- дәннің қабықшалы немесе қабықшасыз болуы (дән қабықты немесе жалаңаш болады) ;
- гүлшоғырының шашыраңқы немесе жинақы болуы;
- гүл қабықшаларының түсінің сары, сұр, ақ немесе қоңырлау болуы;
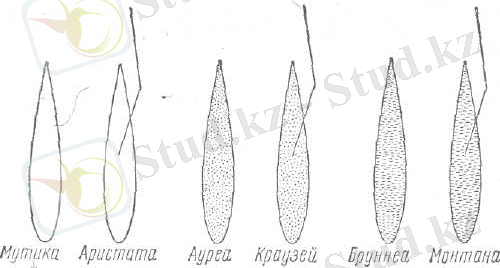
- гүл қабықшаларының қылтықты немесе қылтықсыз болуы. Егер масақшалардың 25% көбі қылтықты болған жағдайда сұлының түрішілік пішінін қылтықты деп санайды (3-сурет) .
Осы белгілер арқылы сұлының барлық түрішілік пішіндерін анықтауға болады .
Қазақстанда және ТМД елдерінде кеңінен таралған сұлы сорттарының басым көпшілігі мутика, аристата және ауреа түрішілік топтарына жатады. Солдатов В. Н 1969 (7)
1. 4 Морфологиялық белгілері және биологиялық ерекшеліктері.
Сұлы қоңырбас (Poaceae) тұқымдастарының сұлы (Avena) туысына жатады. Тамыр жүйесі - шашақ тамырлы. Сұлы дәні себістен кейінгі 9-11 күн аралығында өніп шығады. Алғашқы өскіннің 3-6 жақсы дамыған тамырлары болады. Қосалқы тармырлары сұлының жер асты сабағының бірінші буынынан таралады. Күздік сұлы үшін қосалқы тамырлардың аяз басталғанша дамып жетілуі өте маңызды. Маңызды агротехникалық талаптардың бірі болып сұлының себіс мерзімі саналады. Деркезілік дамыған
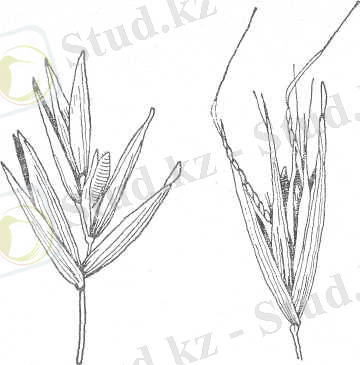
1-сурет. Жалаңаш дәнді сұлының 2-сурет. Құмдық сұлының масақшасының
масақшасының құрылысы. құрылысы.
егер жалаңаш
дәнді болса -
инермис.
3-сурет. Шашыраңқы сыпырғылы және қабықшалы дәнді сұлының түрішілік топтарын анықтау сызба-нұсқасы (нүктелі дәндер сары түсті, сызықшалы дәндерді қоңырлау деп түсіну керек) .
қосалқы тамырлар өсімдікті топыраққа тұрақты орнықтырып қысқы аяздың қысуынан сақтайды. Тамырлар көптеген тамыр түктерімен қоршалған (тамырдың сыртының 90 пайызы) . Сұлы дәнінің өсуі алғашқы тұқым бүршікті тамырдың өсуінен басталады. Өскіннің өсіп даму барысында жер асты буынынан тараған тамырлар қарқынды өсіп шашақ тамыр жүйесін құрайды. Өсу барысында тамыр жүйесіде тереңдікке (0, 7-2 метрге дейін) және жан-жақты бағытта өсе бастайды. Дегенмен, тамыр жүйесінің басты бөлігі топырақтың жоғарғы жер жырту горизонтында орналасады. Топырақтың бұл қабатында көптеген аэробты микробиологиялық үрдістер қарқынды жүреді. Тамыр жүйесі өсімдікті қоректік заттармен және сумен қамтамасыз етеді. Қоректік заттар өсімдікке тамыр түктерінің - ризодермис қабаты арқылы енеді, минералдық элементтердің ассимиляциясы тамырдың қабығында іске асады. Соңғы кездегі зертеулердің нәтижелері тамырда белоктың және кейбір амин қышқылдарының синтезделетінін дәлелдеді. Сұлының сабағы жұмыр және іші бос сабан. Сабақтың ұзындығында 5-6 буын орналасады. Буындары қатты және ісіңкі пішінді келеді (4-сурет) . Сабақ қыстырмалы немесе интеркалярлы меристеманың қызметімен өседі. Бұл кезде буынаралықтар негізі
жағынан созылып өсе бастайды. Өсе келе жоғарғы буынаралықтары төменгі буынаралықтардан әлде қайда ұзарып кетеді. Сондықтан, жетілген өсімдіктің тамырға жақын, төменгі, буынаралықтары қысқа да жоғарғылары ұзын болады. Сұлының сабағы жерасты буындарынан, қосалқы тамырлардың қасынан, жанама сабақтар арқылы түптенеді.
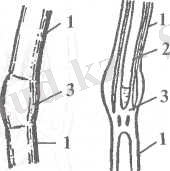
4-сурет. Сұлының сабағының құрылысы: 1-буын аралық; 2- сабақтың ішіндегі қуысы; 3- буын.
Сұлының түптенуі өте маңызды физиологиялық қасиет. Түптену арқылы өнім беретін сабақтар саны артады және сорттар көптеген зиянкестердің әсеріне компенсаторлық жауап бере алады. Түптену арқылы пайда болған сабақтардың барлығы масақ түзе бермейді, оған көптеген физиологиялық факторлар әсер етеді. Өндірістік жағдайда сұлының өнімділік түптенуі 1, 2 - 1, 3 аспайды.
Жапырақтары ұзын, қынаптан және паралелді торланған тақташадан тұрады. Қынап сабақтың буынынан басталып, оның біршама бөлігін қаптап жатады. Буынға бекінген жері сақина тәрізді жуан белдеуге айналады. Жапырақтың буындық сақинасы сабаққа жапырылуға қарсы қосымша мекемдік береді. Жапырақ қынабы мен тақташасының шекарасында ерекше, түссіз тілше болады. Тілше сабақты қымтап қоршап тұрады. Ол сабақ пен жапырақтың арасына су мен зиянкестердің енуіне қарсы тосқауылдық қызмет атқарады. Тілшенің пішіні мен ұзындығы әртүрлі дәнді дақылдарда ерекше болады. Мысалы, арпаның тілшесі сабақты айнала қоршап құлақша тәрізді пішінді болса, сұлының тілшесі өте ұзын келеді. Дәнді дақылдардың осы морфологиялық ерекшеліктері бойынша танаптағы өсіп тұрған, бірақ әлі масақтанбаған өсімдіктерді ажыратуға болады. Страхов Т. Д 1976 (8)
Сұлының гүлшоғыры күрделі сыпырғы. Сыпырғының орталық сабағынан жан-жақты екінші қатарлы сабақшалары таралады, олардан үшінші деңгейдегі, соңғысынан төртінші ары қарай т. с. с. Соңғы сабақшалардың ұшында масақшалар орналасады. Масақшалар екі масақшалық қабықшалармен қоршалған. Қабықшалар дөңес және ірі-ірі жүйкеленген. Қабықты дәнді сұлының масақшалық қабықшалары гүлді толығымен жауып тұрады (жалаңаш тұқымды сұлының масақшасындағы қабықшалар гүлді толығымен жаппайды, гүлден пайда болған дән гүлқабықшаларынан жеңіл ажыратылып тұрады, 1-сурет) . Масақша қабықшаларының ішінде гүл
қабықшалары бар. Сыртқы гүл қабықшасының арқасында (қылтықты сұлыда) қылтық орналасады. Гүл қабықшаларының ішінде 2 ден 4 гүлге дейін болуы мүмкін. Гүл қабықшалары өте жұқа түссіз келеді, олар пісіп жетілген дәнге ажырамайтындай болып жабыспайды. Гүлдің генеративтік мүшелері гүл қабықшаларының ішінде орналасады. Сұлының гүлі үш аталықтан және бір аналықтан тұрады. Аталық екі ұялы тозаңқаптан және жіпшеден құралған. Аналықтың ауызы екі тармақты қауырсын тәрізді. Гүл жатыны жұмыр келген қысқа мойынды. Жатын жанында екі етженді нәзік перделер болады (лодикулалар), пісіп жетілген гүлдерде олар ісініп гүлдің ашылуына жағдай жасайды. Әр ұрықтанған гүлден бір дән пайда болады. Скурыгина В. Н 1974 (10)
Сұлы өздігінен ұрықтанатын дақыл. Оның гүлдеу үрдісі гүл қабықшаларының ішінде өтеді. Кейбір жағдайларда сұлының ашық гүлдеуіде байқалады. Бұған қоршаған ортаның ерекше шарттары әсер етеді. Дегенмен, сұлының 10% айқас тозаңданады деген тұжырым бар. Сыпырғының әр деңгейіндегі гүлдер әр мерзімде гүлдейді. Алдымен сыпырғының негізіне жақын орналасқан масақшалардағы гүлдер, содан кейін ортасындағы, ең соңында сыпырғының ұшында орналасқан масақшалардың гүлдері гүлдейді. Ұрықтану үрдісі барлық дәнді дақылдарға тән жолмен өтеді.
... жалғасыСіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz