Әрекет потенциалы, қозуды өткізу және Н.Е. Введенскийдің парабиоз ілімі

Қазақстан Республикасының Білім және ғылым министрлігі
Семей қаласының Шәкәрім атындағы мемлекеттік университеті
СӨЖ
Тақырыбы: Әрекет потенциалы. Қозуды өткізу. Парабиоз туралы ілім
(14. 7-14. 9)
Орындаған:Құрманта й Ж. Ә.
Тобы:ВС-503
Семей, 2016жыл
Жоспары:
1. Кіріспе
2. Негізгі бөлім
3. Әрекет потенциалы
4. Қозуды өткізу
5. Парабиоз туралы ілім
6. Қорытынды
7. Пайдаланған әдебиеттер.
1. Әрекет потенциалы
Жүйке - ет ұлпалары түрлі тітіркендіргіштер әсерінен қозу жағдайына келеді. Қозудың негізгі белгілерінің бірі торша мембранасының электрлік күйінің өзгеруі болып табылады. Тітіркендіру әсерімен мембрана заряды нөлге дейін күрт төмендейді де, содан кейін кері белгімен қайта пайда болады. Осыдан торша мембранасы зарядының белгісі өзгереді де (ревресия), мембрананың ішкі беті оң, ал сыртқы беті - теріс зарядқа ие болады. Ұлпа тыныштық күйіне оралғаннан соң мембрананың бұрынғы заряды қалпына келеді. Жүйке және ет торшаларының қозуы кезінде пайда болатын мембраналық потенциалдың шапшаң тербелісін (спайк) әрекет потенциалы деп атайды. Потенциалдың осылай тербелісі 1-5 мс. созылады. Жүйке мен бұлшық ет талшықтарында әрекет потенциалдарының деңгейі 115 - 120 мВ шамасында болады.
Әрекет потенциалы 0, 5 - 0, 8 мс созылатын потенциал шыңы және іздік потенциалдан тұрады (1-сурет) . Әрекет потенциалының шыңы қозу процесі кезінде туындайды, ол өрлеу және төмендеу сатыларынан тұрады. Шыңның өрлеу сатысында алдымен мембрананың қалыпты үйектенуі жойылады, сондықтан оны үйексіздену деп атайды. Осыдан кейін потенциал белгісі өзгереді - реверсия байқалады. Шыңның төмендеу сатысында мембрана заряды тыныштық кездегі деңгейге оралады, оны қайта үйектену (реполяризация) деп атайды. Қайта үйектену жағдайында іздік үйексіздену және іздік гиперүйектену құбылыстары байқалады. Іздік потенциалдар қозудан кейін басталатын қалыптастыру процестерінің дамуымен байланысты.
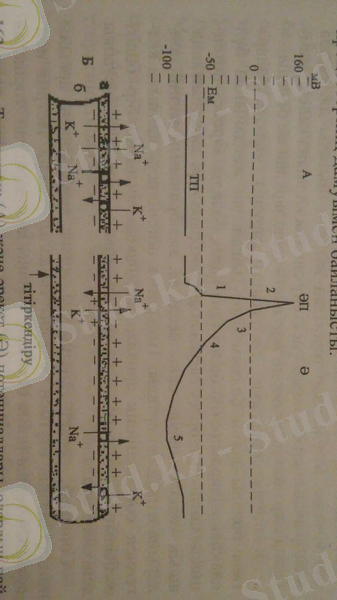
1-сурет. Тыныштық (А) және әрекет (Ә) потенциалдары, олардың пайда болу механизмі (Б) : мВ - милливольттар, Ек - қозу табалдырығы, ТП - тыныштық потенциалы, ӘП - әрекет потенциалы: 1 - үйексіздену; 2 - реверсия; 3 - қайта үйектену; 4 - іздік үйектену; 5 - іздік гиперүйектену; а - мембрана; б - цитоплазма. Тік нұсқама Na және K иондарының ықпалды алмасуын бейнелейді; дөңгелекті нұсқама натрий және калий иондарының ырықсыз тасымалдануы.
Қазіргі мембраналық теория әрекет потенциалының пайда болуын былай түсіндіреді. Табалдырықты және одан күшті тітіркендіргіштер әсерінен торша мембранасында үйексіздену процесі жүреді. Үйексіздену белгілі шекке жеткен кезде бір сәтте көптеген натрий арналары ашылады да, торша мембранасының натрий иондарына өтімділігі тыныштық жағдайымен салыстырғанда, 500 есе артады. Осының нәтижесінде натрий иондарының легі торша аралық сұйықпен торша ішіне өтеді. Оң зарядтың азаюы салдарынан торшааралық сұйықтағы аниондар әсерімен (көбінесе, хлор иондары) мембрана беткейі теріс зарядталады. Бірақ мембрананың натрий иондарына өтімділігі қысқа мерзімге ғана (0, 5 - 1, 5 мс) өседі де, одан әрі натрий арналары жабылып, бұл иондардың өту қарқыны төмендейді.
Бірыңғай салалы ет талшықтарында әрекет потенциалын тудыруда натрий және калий иондарымен бірге кальций иондары да қатысады.
Әлсіз, табалдырықтан төмен тітіркендіргіштер әсерінен мембрананың натрий иондарына өтімділігі аз ғана артады да, үйексіздену қажетті шекке жетпей қалады. Үйексізденудің бұл түрін шектелген, таралмайтын потенциал деп атайды. Ол ұлпаның басқа бөлігіне таралмайды, туындаған жерінің маңында өшіп қалады. Әлсіз тітіркендіргіштер әсері қайталанса, олар жинақтала келіп, үйексізденуді шектік деңгейге жеткізіп, әрекет потенциалын тудыруы мүмкін.
2. Қозуды өткізу
Қозу процесіне жүйке немесе ет талшықтары бойымен өздігінен таралу қасиеті тән, сондықтан қозғыш ұлпалардың бір жерінде пайда болған әрекет потенциалы оның барлық бөліміне таралады. Тітіркеністі таратуға қажет қуат сол қозғыш ұлпалардың өзінде АТФ-тың ыдырауы нәтижесінде бөлінеді. Күші жағынан әрекет потенциалы ұлпаның қатар жатқан учаскесін тітіркендіруге қажетті табалдырықты күштен 5 - 10 есе басым келеді. Осы таралатын жүйке импульсі шеткей жүйке ұштарынан (рецепторлардан) хабардың ОЖЖ-де орналасқан жүйке орталықтарына, ал одан эффекторларға (атқарушы органдарға) өтуін қамтамасыз етеді.
Әрекет потенциалы айналмалы немесе шектелген токтың пайда болуы нәтижесінде таралады. Жүйке немесе ет талшықтарының белгілі бір учаскесі қозып, әрекет потенциалы туындаған кезде, осы қозған және көршілес - қозбаған учаскелер арасында потенциалдар айырмасы пайда болады. Ол қозған және қозбаған учаскелердің ішкі және сыртқы беткейлерімен айналып отыратын шектелген, айналмалы токтың пайда болуына соқтырады. Мембрананың ішкі бетінде ток қозған учаскеден қозбаған учаскеге, ал сыртқы бетінде - қозбаған учаскеден қозған учаскеге бағытталады (2, 3-суреттер) .
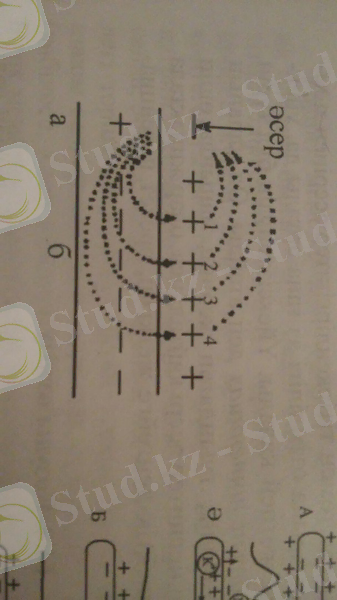
2-сурет. Айналмалы ток түрінде қозудың ет талшықтары мен миелинсіз нервтерде таралуы: а - қозған бөлік, ә - қозбаған бөлік. Нұсқамалар - токтың таралу бағыты.
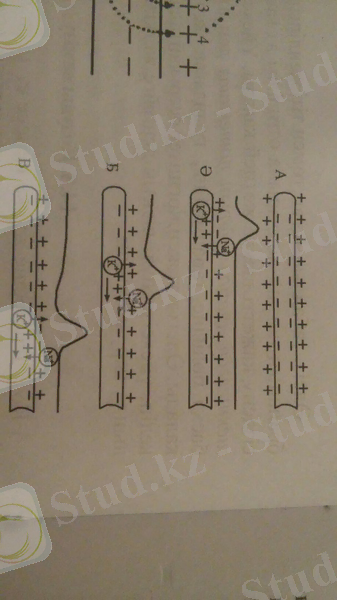
3-сурет. Қозудың нерв бойымен таралу сызбасы: А - физиологиялық тыныштық күйі, Ә, Б, В - әрекет потенциалының таралу сатылары. Натрий иондарының енуі мембрананы үйексіздендіреді, калий иондарының шығуы - гиперүйектендіреді.
Айналмалы токтың әсерінен мембрананың қозбаған учаскесі тітіркеніп, оның натрий иондарына өтімділігі артады да, осы жерде әрекет потенциалы пайда болады. Ал алғашқы қозған учаскеде тыныштық потенциалы қалпына келеді. Осы процесс қайталана отырып, қозу импульсінің мембрана бойымен өшпей, ілгерілей таралуын қамтамасыз етеді. Импульстердің кері бағытта оралуына қозу толқыны өткен учаскеде рефрактерлік аймақтың пайда болуы мүмкіндік бермейді. Сонымен, қозу толқыны ет немесе жүйке талшықтары бойымен әрекет потенциалы түрінде таралады.
Миелинді жүйке талшықтарында айналмалы токтар тек қана іргелес орналасқан қозған және қозбаған Ранвье үзілімдері арасында пайда болады (4-сурет) . Айналмалы токтың осы үзілімде шығуы мембрананы үйексіздеп, әрекет потенциалын тудырады. Дәл осы сипатта келесі үзілім қозып, қозу толқыны жүйке бойымен тарала береді. Осыған байланысты қозу толқыны миелинді жүйке талшықтарымен секірмелі, сальтаторлы (латын тілінде Salto - секіремін, ырғимын деген мағына береді) түрде, ырғый тарайды. Қозу импульсі мұндай сипатпен тарағанда қуат аз шығындалады және оның таралу жылдамдығы шапшаңдайды. Мысалы, қозу импульсінің таралу жылдамдығы миелинсіз жүйке талшықтарында секундына 0, 5 - 3 м, ет талшықтарында - 5 м, ал миелинді жүйке талшықтарында - 30 - 120 м. Айта кетер жәйт, жүйке талшықтары неғұрлым жуан болса, соғұрлым олар қозу импульсін шапшаң өткізеді. Жуан жүйке талшықтарының қозғыштық қасиеті де жоғары болады. Сондықтан аралас жүйкелерді әлсіз тітіркендіргіштермен тітіркендіргенде алдымен жуандау жүйке талшықтары қозады. Тітіркендіргіш күші арта түссе, біртіндеп майда жүйке талшықтары қозады да, әрекет потенциалының амплитудасы өсе бастайды.
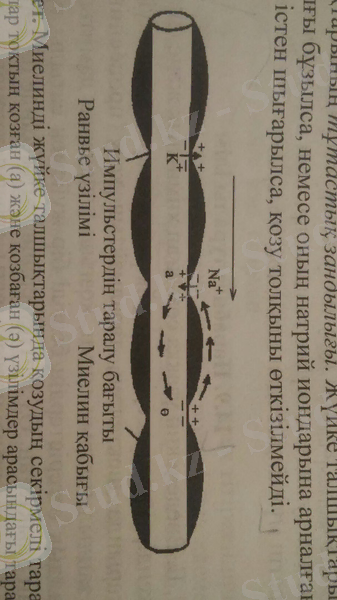
4-сурет. Миелинді жүйке талшықтарында қозудың секірмелі таралуы: нұсқамалар токтың қозған (а) және қозбаған (ә) үзілімдер арасындағы таралуын көрсетеді.
Жүйке талшықтары қозу импульсін белгілі заңдылықтарға сәйкес өткізеді. Қозу толқыны жүйке талшығы тұтас және оның мембранасының иондық арналарының функционалдық белсенділігі қалыпты күйде болғанда ғана өткізіледі. Бұл - жүйке талшықтарының тұтастық заңдылығы. Жүйке талшықтарының тұтастығы бұзылса, немесе оның натрий иондарына арналған арналары істен шығарылса, қозу толқыны өткізілмейді.
Қозу толқыны жүйкенің тітіркенген жерінен екі бағытта таралады. Бұл - жүйке талшықтарында қозуды қос бағытта өткізу заңдылығы. Бұл құбылыс жүйке құрамында орталыққа тепкіш және орталықтан тепкіш талшықтардың болуымен байланысты деген болжам да бар. Ет талшықтары да қозуды тітіркенген жерден қос бағытта өткізеді.
Қозу толқыны бір жүйке талшығынан көршілес талшыққа берілмейді. Бұл қозуды жеке - дара өткізу заңдылығы деп атайды. Жүйке талшықтарының бұл қасиеті оларды қоршап тұрған миелин қабығының болуына байланысты. Миелинсіз талшықтарда аталған құбылыс оларды қоршап тұрған сұйық кедергісінің төмендігімен байланысты. Осының нәтижесінде айналымды токтың негізгі тұзақтары көршілес талшықтарға жетпей торша аралық сұйық арқылы жайылып кетеді.
Әлбетте қозу кезінде жүйке талшықтарында зат алмасу процесі күшейіп, оттегі шығыны өседі, көмір қышқылының бөлінуі артады, қуаты мол фосфорлы қосылыстардың (АТФ пен креатинфосфат) шығыны көбейіп, сүт қышқылы көп түзіледі, аммиак мол бөлінеді. Қозу кезінде жылу да көп өндіріледі. Бірақта қозған жүйке талшықтары қуатты көп шығындамайды. Олар қуатты тек натрий - калий сорабының қызметін атқаруға жұмсайды. Қуатты аз шығындап, ресинтез процестерінің күшті жүруімен байланысты жүйке талшықтары қажымайды. Тіпті, бірнеше сағат бойына үзбей тітіркендіргеннің өзінде жүйке талшықтары өткізгіштік қасиетін сақтайды (Н. Е. Введенский) . Тұтас организмде жүйке талшықтарының қажымауының себебі олардың құбылмалылығының жоғарылығымен байланысты. Жүйке орталықтарында қозу импульстері секундына 50 - 100 рет жиілікпен туындаса, жүйке талшықтарының импульстер өткізіп үлгеретін мүмкіндігі одан әлдеқайда жоғары.
3. Парабиоз туралы ілім
Н. Е. Введенский ұзаққа созылған химиялық және физикалық тітіркендіргіштер әсерінен жүйке-ет препаратының жүйке талшықтарында шектелген, таралмайтын қозу процесінің пайда болу заңдылықтарын ашып, қозғыш ұлпаларда дамитын функционалдық өзгерістердің белгілі бір жүйемен сатылы түрде өрбитінін анықтаған. Өз зерттеулерінің нәтижесінде ол 1901 жылы «Қозу, тежелу және наркоз» атты еңбегін жариялап, парабиоз туралы теорияны ұсынды (грек тілінде para-маңы, bios-өмір, тіршілік, демек парабиоз деген сөз тіршілік шегі, өлара күй деген мағына береді) . Бұл теория қозу және тежелу процестерінің биологиялық мәнін ашып, қозудың тежелуге айналу сырын және соңғы құбылыстың тіршіліктің тоқтауына соқтырар қауіпі бар екенін көрсетеді.
Парабиоз құбылысының физиологиялық мәнін Н. Е. Введенский ашса, оның шәкірттері бұл құбылыстың табиғатына талдау жасады.
Н. Е. Введенский тәжірибесін бақаның жүйке-ет препаратымен жүргізген. Алдымен жүйкенің бір учаскесі эфир, хлороформ, новокаин, калий хлориды, т. б. тітіркендіргіш әсерімен еліттірілген (альтерациялаған) . Жүйке еліткенге дейін еттің жиырылу күшінің тітіркендіру күші мен жиілігіне сәйкес өзгеретіні анықталған. Содан соң осы еліткен жерге, оның алдына және етке жақын бөлігіне индукциялық катушка электродтары қойылған (5-сурет) . Жүйкенің еліткен жерінің электрлік күйін гальванометрмен бақылап отырған.
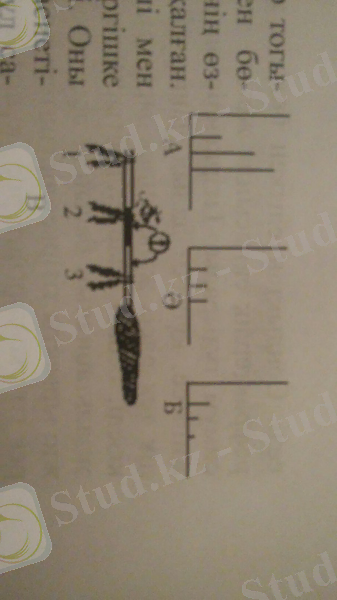
5-сурет. Парабиоз (өлера күй) сатылары: А- тітіркендіру күші мен жауап реакция арасындағы сәйкестік; Ә- теңестіру сатысы; Б- бұрмалау сатысы.
Енді осы жүйкені электр тогымен тітіркендіргенде еліткен бөліктің функционалдық күйінің өзгерістерінде үш саты байқалған. Жүйке еліткеннен соң күші мен жиілігі әртүрлі тітіркендіргішке ет бір деңгейде жауап береді. Оны жүйкенің құбылмалылық қасиетінің өзгерісімен түсіндіріп, бұл сатыны Н. Е. Введенский теңестіру немесе трансформациялық саты деп атаған.
Тәжірибе әрі қарай жалғасқанда препарат күшті және жиі тітіркендіргішке әлсіз жиырылумен, ал әлсіз және сирек тітіркендіргішке - керісінше, күшті жиырылумен жауап берген. Ұлпа қасиетінің бұлай құбылуы бұрмалау, парадоксалдық саты деп аталған. Осыдан кейін препарат ешбір тітіркендіргішке жауап бермеген. Бұл тежелу сатысы. Еліттіруші әсер әрі қарай жалғасса, тежелу сатысы тереңдеп, тіршілік белгілері - қозғыштық пен өткізгіштік, жойылады, ұлпа өлара (парабиоз) күйіне көшеді. Байқалған өзгерістерді парабиоз сатылары дейді.
Еліттіруші факторлардың әсері тоқтаса, парабиоз тиылып, жүйкенің жансызданған бөлігінің бастапқы функционалдық күйі қалпына келеді. Айта кетер жайт, парабиоздан ұлпа кері тәртіппен шығады (алдымен тежелу сатысына көшеді, содан соң бұрмалау, теңестіру сатылары арқылы қалыпқа келеді) . Ал еліттіруші фактор әсері тиылмаған жағдайда ұлпа өліп қалады.
Жүйкенің еліткен жерінде әртүрлі тітіркендіргіштер әсерінен туындаған әрекет потенциалын зерттеу парабиоздың теңестіру сатысында жүйкенің еліткен бөлігінде құбылмалылық, онымен байланысты өткізгіштік төмендемейтінін көрсеткен. Сондықтан жүйкенің бұл бөлігі зақымданбаған бөліктен жеткен жиі тітіркеністерге синхронды жауап бере алмайды, оны өзгертіп, өз күйіне сәйкес сирек ырғақпен жауап қайтарады. Ал, сирек ырғақты тітіркеністер бұл жерден өзгеріссіз өткізіледі. Еліткен бөлікте тітіркеніс күші мен ырғағының өзгеруі нәтижесінде олар белгілі шамада теңгеріледі, сондықтан еттің жиырылу күші де теңеседі.
... жалғасыСіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz