Ақуыз синтезі: молекулалық механизмдері мен табиғаттағы маңызы

Ақуыз синтезі және оның табиғаттағы маңызы
Жоспар:
І. Кіріспе . . . 3
ІІ. Негізгі бөлім . . . 4-29
2. 1 Белоктар тұралы жалпы түсінік . . . 4-8
2. 2 Белок конформациясы . . . 9-17
3. Клеткадағы белок синтезі және оны реттеу . . . 18-22
3. 1 ДНҚ репликациясы . . . 23-25
3. 2 РНҚ құрылысы . . . 26-27
ІІІ. Қорытынды . . . 28-29
IV. Әдеби шолу . . . 30
Кіріспе
Организм клеткасының химиялық құрамы аса бай және алуан түрлі. Онда көптеген реакцияларға қатысатын және метаболизм түзетін әр түрлі заттар бар. Мұндай алмасу нәтижесінде заттар үздіксіз өзгеріп, ыдырайды және осының арқасында жаңа заттар түзіледі. Алмасу реакциялары белгілі бір қатаң тәртіппен өтеді және әр түрлі ферменттердің әсерімен реттеліп отырады. Тірі клеткада болатын ерекше жағдайлардың арқасында реакциялар жоғары жылдамдықпен өтеді. Клетканың немесе протопластың химиялық құрамына талдау жасағанда, біріншіден, ондағы заттардың аса көптігі мен алуан түрлілігіне, екіншіден, талдау барысында тірі клеткаға тән емес заттардың пайда болуына байланысты көптеген қиындықтар туады. [3]
Академик В. И. Вернадский жер қыртысындағы элементтердің барлығы да (тіпті сирек кездесетін элементтер де) клеткада белгілі бір мөлшерде кездеседі деп санайды. Алайда тірі заттың құрамына бар болғаны 22-27 элемент кіреді. Кез келген организмнің тірі затының элементтік құрамы мынадай: С, О, Н, N. Р, S, Na, К, Мg, Са, С1, Мn, Ғе, Со, Сu, Zn, В, Аl, V, Мо, I, Si, Ni, Сг, Sе, Sn, Аs. Көрсетілген элементтердің 16-сы барлық организмдерде кездеседі. [3]
Тірі клетка затының құрамын дәлелдеу әлдеқайда қиын. Ішекте тіршілік ететін бактерия Escherihia coli осы мәселені зерттеуде ыңғайлы материал болып табылады. Оның құрамында мынадай заттар табылған: су - 70%, бейорганикалық иондар - 1%, көмірсулар - 3%, амин қышқылдары - 0, 4%, нуклеотидтср - 0, 4 %, липидтер - 2 %, белоктар - 15%, ДНҚ - 1 %, РНҚ -6%. Барлық басқа организмдердің тірі затының құрамы да осы сияқты деген пікір бар. Тірі заттың құрамына кіретін молекулаларды биомолекула деп атайды, демек олардың ерекше құрылымы және өзара әрекеттесуі тіршілік негізінде жататын процестерді қамтиды. Бұл заттардың молекулаларының құрылымы, физикалық-химиялық қасиеттері және олардың тірі клеткадағы атқаратын физиологиялық ролі жөнінде қысқаша мәліметтерге тоқталайық. Осының ішінде белоктар және оның синтезделуіне тоқтап өтсек. [3]
2. 1 Белоктар тұралы жалпы түсінік.
Тірі организмдердің негізін құрайтын белоктардың маңызды ролін Ф. Энгельс: «Тіршілік-белокты денелердің тіршілік ету әдісі»-деп көрсеткен болатын. Белок - организмдегі заттардың ең күрделісі, ал оның элементтік құрамы айтарлықтай қарапайым болып келеді. Онда 51-53% көміртегі, 16-18% азот, 7% сутегі, 21-23% оттегі, 0, 7-1, 3% күкірт болады. Кейбір белоктарда бұған қосымша фосфор да кездеседі. Үрмебұршақ, соя, күнбағыстың тұқымында белоктың мөлшері едәуір көп болады. Бұл өсімдіктер тұқымының үгілген массасын сумен, тұзды, спиртті және әлсіз сілтілі ерітінділермен тұндыру жолымен олардан белокты бөліп алу қиын емес. Күшті қышқылдармен және сілтілермен бірге қайнатқан кезде, сондай-ақ ферменттердің әсерімен белок амин қышқылдарының қоспасына ыдырайды. [3]
2. 1. 1 Амин қышқылдары өсімдікте кетон қышқылдарын тікелей аминдеу немесе қайта аминдеу деп аталатын екі негізгі реакция арқылы синтезделеді. Олардың құрамында карбоксил тобынан басқа, амин тобы да болады. Амин қышқылдарының тірі организмдер үшін физиологиялық маңызы бар екі қасиетіне назар аударайық. Оның бірі амфолиттік, екіншісі оптикалық активтік. Сулы ерітінділерде амин қышқылдарының СООН және NH 2 тобы ортадағы реакцияға қарай диссоциацияланады. Мысалы, сілтілі ортада амин қышқылының карбоксил тобы диссоциацияланғанда минус заряд, қышқыл ортада NH 2 тобы диссоциацияланғанда плюс заряд пайда болады.
Сөйтіп, амин қышқылы ортадағы реакцияға байланысты бірде қышқыл, бірде сілті ретінде қызмет атқарып, амфолиттік қасиет көрсетеді. Бейтарап ортада олар қос зарядты цвиттерион түрінде болады. [3]
Амин қышқылдарына оптикалық активтік тән. Олардың ерітіндісі арқылы полярланған сәулені өткізгенде сәуленің поляризация бағыты өзгереді. Оптикалық активтік кеңістіктік изомерияға, яғни хиралды көміртегі жағдайында атомдар тобының әркелкі орналасуына байланысты. Мысалы аланин амин қышқылы екі түрлі болып кездесуі мүмкін.
Өсімдік клеткасынан 150-ден астам амин қышқылдары табылған. Олардың көбі фотосинтез кезінде немесе топырақтан азотты қабылдау кезінде, жалпы метаболизм процесінде түзіледі. Адам организміне қажетті амин кышқылдары азықпен бірге қабылданады. Олардың ішінде валин, лейцин, изолейцин, метионии, треонин, фенилаланин, лизин, аргинин, гистидин және триптофан ерекше қажет. Азық-түліктің құрамында осы амин қышқылдарының болуы азық-түліктің қоректік құндылығын арттыра түседі. Олар міндетті түрде қабылданатын амин қышқылдары деп аталады. [3]
Бүйірлік радикалдарының құрылысына байланысты барлық амин қышқылдары 4 класқа бөлінеді. I класка гидрофобты амин қышқылдары аланин, лейцин, изолейцин, валин, пролин, фенилаланин, триптофан, метионин жатады. II класқа поляризацияланған, электр заряды жоқ амин қышқылдары серин, треонин, тирозин, цистеин жатады. Бұл амин қышқылдары сумен Н-байланыс түзуге қабілетті. III класқа плюс зарядты лизин, аргинин, гистидин амин кышқылдары жатады. IV класс минус зарядты аспарагин және глутамин амин қышқылдарын біріктіреді. Олар рһ бейтарап аймақта тұрғанда теріс зарядқа ие болады.
Өсімдіктерде амин қышқылдары кең таралған. Ал белоктардың құрамына 20-22 амин қышқылы кіреді. Белок молекуласының конформациясы амин қышқылдарының бүйірлік радикалдарына байланысты.
2. 1. 2 Белок молекуласының құрылымы. Белок химиясының тарихы, бәлкім 1745 ж. "Белок ғылым және өнер институтының комментарийларында (хабарла-маларында) " итальяндық Я. Беккаридің жарияланған жұмысымен басталған бо-лар. Бидай ұнынан бұрын белгісіз зат алғанын ғалым осы еңбегінде хабарлаған. Бидайдан алынған клейковина ұқсас болған. [3]
Осылайша алғашқы белок препараты дайындалған. Сөйтіп, белоктық заттарды зерттеудің ұзында және өте қиын тарихы басталды. Ғалымдар ең қарапайым белок - инсулиннің молекуласының құрылысын ашқанша 200 жылдан артық уақыт өтеді. Осы мезгіл бойы белок құрамы, оның ұйымдасу жөнінде ондаған болжамдар ұсынылып және олар бекерге шығарылған. Барлық болжамдар ғылыми мұрағаттарда жайлы орын тапқан.
Әйтеуір, 1945 ж. көптеген зерттеушілердің күшімен белок молекулаларының бір ұзын тізбекке жалғанған амин кышқылдарынан тұратыны анықталды. Белок молекуласын түзетін амин қышқылдары көп емес - барлығы 20. Ал қараңыздар, белоктар молекуласы амин қышқылдарының кезектесіп орналасу бір ізділігімен ерекшеленеді. Мысалы, бір белокта валиннен кейін триптофан келеді. Олардың орнын ауыстыру арқылы басқа қасиеті бар жаңа молекула алуға болады. Айталық, егер біріншісі организмде маңызды қызмет атқарса, екіншісі мүлде қажетсіз болуы мүмкін. [3]
Ал енді математикаға оралайық. Егер біз белок молекуласын әрдайым жаңадан 20 амин қышқылынан құрастырсақ, төрт амин қышқылынан тұратын пептид 20-20-20-20= 16 әртүрлі тізбек, ал амин қышқылының п санынан - 20" тізбек береді. Сөйтіп, орташа ұзындығы 300-ге таяу амин қышқылынан тұратын 10 390 әртүрлі белок болуы мүмкін. Ал жиырма амин қышқылынан тұратын молекула "нағыз" белок молекулаларымен салыстырғанда өте кұртымдай құрылым. Осыдан бар жоғы 20 амин қышқылынан ғана құрылған белоктардың шексіз түрлерінің пайда болатынына көз жеткізуге болады.
Белоктар - биологиялық макромолекулалардың негізгі кластарының бірі. Клеткалардың көп мөлшері белоктардан тұрады: құрғақ заттың жартысы белоктардың үлесіне тиеді . Клетканың құрылымын және пішінін белоктар анықтайды; сонымен қатар, молекулалық тану және катализдік құрал қызметін атқарады. Клетка құрылысына қажетті ақпаратты ДНҚ "шартты" түрде, клеткалық процестерге тікелей араласпай, ұстайды. Мысалы, оттегін тасымалдау гемоглобинге тән қасиет, бұл белокқа жауапты геннің оған қатысы жоқ. Компьютерлік терминологияны қолдансақ, нуклеин қышқылдары аналық клеткадан алынған нұсқауды "бағдарламамен қамтамасыз етеді". Белоктар "аппараттың қамсыздандыру" - клетканың жадында сақталған бағдарламаның физикалық механизмін іске асырады. Осындай нуклеин кышқылдары мен белоктар қызметінің айырмашылығы, оларды құрайтын суббірліктердің химиялық табиғатынан байқалады.
ДНҚ және РНҚ молекулалары химиялық жағынан өте ұқсас алып молекулаларды құрайтын нуклеотидтерден тұрады, олардың қасиеті бірізділігіне көп байланысты емес. Ал белоктар әртүрлі 20, бір біріне ұқсамайтын, амин қышқылдарынан тұрады, олардың әрқайсысының химиялық қасиеті ерекше .
Атқаратын қызметінің шеңберінің кеңдігі, олардың химиялық құрылымының және кеңістіктегі пішінінің молшылығымен сипатталады. Әртүрлі белоктардың химиялық қасиеттерінің әмбебаптығы да осыған негізделген.
Белок макромолекуласының күрделі құрылымында бірнеше деңгейлер болады. Оның ішіндегі алғашқы ең қарапайымы полипептид тізбегі, яғни өзара пептидті байланыспен жалғанған амин қышқыл буындарының тізбегі. Бұл - белоктың алғашқы қүрылымы деп аталады; мұндағы байланыстардың бәрі ковалентті, яғни нағыз химиялық берік байланыс. [6]
Полипептидтік тізбектің бөлігінің екінші құрылымы деп бүйірлік тізбектің конформациясын есептемегендегі сол фрагменттегі негізгі тізбектің конформациясын айтады. Мұнда белок жіпшесі шиыршық тәрізді ширатылып тұрады. Шиыршық орамдары тығыз орналасады да, көршілес орамдардағы атомдар мен амин қышқылының радикалдары арасында бір-біріне тартылу пайда болады. Атап көрсеткенде, көршілес орамдарда орналасқан пептидтік байланыстар арасында (NH-пен СО-топтары арасында) сутектік байланыстар түзіледі. Сутектік байланыс ковалентті байланыстан әлдеқайда әлсіз, алайда, бірнеше қайталанғаннан кейін ол да берік ілінісетін болады. Сансыз көп сутектік байланыстармен "жөрмелген" полипептидтік шиыршық едәуір тұрақты құрылым болып саналады. Белоктың екінші құрылымы одан әрі тағы түйінделеді. Ол әлдеқайда қызық иіріле келіп оралады, бірақ соның өзінде де ол бір қалыпты және әр белокта ерекше пішінде болады. [3]
Жекеленген полипептидтік тізбектің барлық атомдарының кеңістікте орналасуын белоктың үшінші құрылымы деп атайды.
Кейбір белоктар бірнеше полипептидтік тізбектерден құралады. Әрбір тізбек - бір суббірлік немесе мономер. Үшінші құрылымды жалғастырушы байланыстар сутекті байланыстардан да әлсіз болады. Оларды гидрофобты деп атайды.
Бұл - полюссіз молекулалардың немесе полюссыз радикалдардың арасындағы ілініс күші. Ондай радикалдар амин кышқылдарының бірсыпырасында кездеседі. Полипептидтік байланыстағы гидрофобты радикалдардың бір-біріне тартылып, жабысу себебі суға шашыраған майдың немесе басқа бір гидрофобты заттың тамшыға жиналу есебімен бірдей. Гидрофобты ілінісу күші өте әлсіз байланыс болғанымен де, бірнеше рет қайталанатындығынан олардың барлығы қосылғанда едәуір әрекеттесу қуатын береді. Белок молекуласының аса күрделі құрылымын ұстап тұруға "әлсіз" байланыстардың қатысуының салдарынан, ол анағұрлым тұрақты және өте қозғалғыш болады. Кейбір белоктардың макромолекуласының үшінші құрылымын ұстастыруға амин қышқылы цистеиннің арасында пайда болатын, S-S байланыс деп аталатын берік коваленттік байланыс әжептәуір роль атқарады, бірақ тұрақты үшінші құрылым ұйымдасу үшін олардың болуы шарт емес. Мысалы, миоглобин және гемоглобин молекулаларында S-S байланыстар жоқ. Көпшілік жағдайларда белоктың бірнеше макромолекуласы бір-бірімен қосылып. өте үлкен агрегаттар түзеді. Мысалы, гемоглобин осы белоктың төрт макромолекуласының жиынтығы болып саналады. Тек осылай жиялу арқылы ғана гемоглобин қалыпты қызмет атқарады, яғни оттегі молекулаларын қосып алып тасымалдай алады екен.
Белоктың төртінші құрылымы деген ұғым - мономердің кеңістіктегі өзара орналасуы, олар бірнеше суббірліктерден құралып, белок молекуласын құрады. Гемоглобин молекуласы әдеттегі тетрамер болғандықтан, оның құрамына екі бірдей α-тізбек және екі ұқсас β-тізбек кіреді. Төртінші құрылым байланыстар (сутекті, гидрофобты) әлсіз жалғасқан, ал кейде S-S-байланыстармен де жүзеге асырылады.
Дисульфидтік көпірше - бұл екі цистеин қалдықтарының арасындағы ковалентік байланыс. Мұндай көпіршелер кейбір секреторлық белоктарда кездеседі. Көпірше глобуланың ішінде де, сондай-ақ оның үстінде де орналаса береді. Көптеген белоктарда, қалыпты цистеиндер болғанмен, дисульфидтік көпіршелер болмайды. Атқаратын қызметіне сай белоктар: глобулалық белоктар, шамалап салыстырсақ сфера пішіндес, катализ, транспорт немесе реттеу сияқты арнаулы процестерге қатысады. Кейбір белоктық заттардың құрамына фосфор кіреді, аз мөлшерде кейде темір, мыс, йод, хлор, бром және т. б. элементтер де кездеседі. [3]
Белоктардың құрылымдык элементіне амин қышкылдары енеді. Белок молекулалары бір-бірімен бірнеше байланыстар арқылы біріге алады. Әсіресе жануарлар мен өсімдіктер организміндегі кездесетін белоктардың көпшілігінің пептидті байланыс арқылы байланысатыны анықталды. Пайда болған амин қышқылы үшінші бір амин қышқылымен байланыса алады. Осы пептидті байланысты анықтаған және дамытқан немістің атақты ғалымы Эмиль Фишер болды. [3]
Амин қышқылының белок молекуласында алатын орны тек өзіне ғана тән. Егер амин қышқылы басқа амин қышқылымен орын алмастырса, қасиеті өзгереді. Сенджер деген ғалым белок молекуласында кездесетін қышқылдарды, олардың табиғатын анықтады. Инсулин белогы ұйқы бездерінің гормондарынан алынады. Қарапайым белокка жатқанымен тіршілікте маңызы зор. Осы инсулин белогінің полипептидті қалдықтардан тұратыны анықталды.
Белок молекуласына кіретін амин қышқылдарының саны өте көп. Гликокол немесе глицин H
2
N-СН
2
-СООН бұдан басқа аланин амин қышқылы немесе аминопропион қышқылы
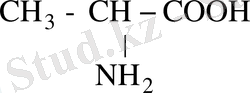 сол сияқты цистеин амин қышқылы, метионин және т. б. кіреді.
сол сияқты цистеин амин қышқылы, метионин және т. б. кіреді.
Белок молекуласына моноамино-, диамино-, тиоқышқылдар және басқа қышқылдар енеді. Белоктардың біріншілей құрылымы кейбір белоктар үшін ғана анықталады. Мысалы: инсулин, рибонуклеаза, т. б. І950 ж американ ғалымы Полинг белок молекуласының біріншілей құрылымын көрсетті. Белоктар 2 үлкен топқа бөлінеді:
1. Протеиндер - қарапайым /жай/ белоктар.
2. Протеидтер - күрделі белоктар.
Белоктар өте күрделі жоғары полимерлі заттар. Оларды құрайтын мономерлер - амин қышқылдары бір-бірімен жалғасып полипептид береді, осы полипептидтердің бірігуінен белок молекуласы пайда болады. Әр организмнің өзіне тән белогы болады. Неміс ғалымы Абдер Гальден былай деген "егер 32 әріптен канша сөз жасауға болса, 22 амин қышқылдарынан сонша белок молекуласын жасауға болады". [6]
2. 1. 3 Белоктың физикалық және химиялық қасиеттері организмнің тіршілік әрекетінің негізін құрайды. Белоктардың коллоидтық қасиеті, коацерват түзушілігі, денатурация құбылысына ұшырауы, сумен байланыс жасай отыра гидрадтануы, электр заряд түзушілік қасиеттері маңызды роль атқарады.
2. 1. 4 Белоктың атқаратын қызметі. Организмде белок алуан түрлі қызмет атқарады. Белоктың қызметін көбіне жекелеген молекулалар да жүзеге асыруы мүмкін. Белоктың ең басты қызметі - катализаторлық қызмет. Барлық тірі организмдерде зат алмасу реакциялары ферменттердің әсер етуімен жүзеге асады. Белгілі ферменттердің барлығы белоктардан құралған.
Заттарды тасымалдау да белоктың маңызды бір қызметі. Заттардың клетка мен органоидтар ішінде қозғалуын белок реттеп отырады, яғни оларды активті түрде тасымалдайды. Соңғы кезде клетка мембранасының құрамында түрлі тасымалдаушы белоктардың болатыны анықталған.
Белок сондай-ақ организмнің иммундық қасиеттерін жүзеге асырады. Сонымен бірге белоктың тағы маңызды қызметінің бірі - оның құрылыс материалы ретінде пайдаланылуы. Белок барлық протоплазмалық органоидтардың негізін кұрайды. Ол құрылым компонентінің бірі ретінде барлық клетка мембраналарының құрамына кіреді, тіпті сұйық гомогенді цитоплазмалық матриксте де белок кездеседі. [3]
2. 2 Белок конформациясы.
Белок молекуласының конформациясы амин қышқылының бірізділігімен анықталады. Полипептидтік тізбекте көптеген байланыстардың төңірегінде айналым болуы мүмкін, сондықтан белоктың кез келген молекуласы өте көп, әртүрлі пішін немесе конформация түзетін қабілеті бар. Бірақ биологиялық жағдайда полипептидтік тізбектердің көпшілігі осы конформациялардың бір түрінде ғана кездеседі. [1]
Бұл амин қышқылдарының бүйірлік топтарының өзара және сумен өте осал ковалентті емес арақатынасына байланысты. Белгілі бір конформация әдеттен тыс тұрақты бола алады, ал оның қандай болатыны - амин қышқылдарының полипептидтік тізбекте орналасуына байланысты. Көптеген белоктардың полипептидтік тізбектері өз бетімен түзілу конформацияға түйінделеді. Мысалы белокты жазу, немесе денатурациялау (лат. «denaturare» - табиғи қасиетін жою) арқылы алғашқы конформациясын жоғалтқан икемді полипептидтік тізбекке айналдыруға болады. Бірақ, мейлінше жұмсақ денатурациялау әсері әдетте қайтымды, жазылған полипептидтік тізбек табиғи конформациясына өзі түйінделді. Оның мұндай қылығы белок молекуласының конформациясын анықтайтын ақпараттың бәрі амин қышқыл бірізділігінде екендігінің айғағы. Полипептидтік тізбектің түйінделуін басқаратын маңызды факторлардың бірі - полярлық және полярлық емес бүйірлік топтардың орналасу тәртібі болып табылады. [1]
Белок синтезінің барысында олар дың көпсанды гидрофобты (гр. "һуdог" -су, "рһоЬоs" - қорқу) бүйірлік топтары белок глобуласының ішіне жиылуға тырысады, себебі судан құтылуына мүмкіндік туады. Сол кезде барлық полярлық топтар белок молекуласының үстіне жиылады, онда бұл түзілімдер сумен және басқа полярлық топтармен арақатынас жасай алады. Пептидтік топтар өздеріде жеткілікті полярлы, сондықтан сутектік байланысты бір-бірімен және полярлық бүйірлік топтармен құруға тырысады. Осындай жолмен белок глобуласының ішіндегі, түгелге жуық полярлық топтардың жұптасуы жүреді. Сөйтіп, сутектік байланыстар белок молекуласының түйінінде бір полипептидтік тізбектегі әртүрлі бөлшектерінің арақатынасында басты роль атқарады. Сонымен қатар, олар белок молекуласының үстіндегі арақатынастарда да маңызды орын алады. Цитоплазмадан тыс белоктар (секреторлык белоктар немесе клетка үстіндегі белоктар), бір полипептидтік тізбектің әртүрлі бөлшектерінің арасында косымша коваленттік байланыстар түзеді. Мысалы, цистеиннің екі SH-топтарының арасындағы дисульфидтік байланыстар ( SS-көпіршелер деп те аталады), түйінделген полипептидтік тізбектің көршілес келетін бөлшектерінің арасында жүреді, олар клеткадан тыс белоктардың кеңістіктегі құрылымын тұрақтандырады; дұрыс түйінделу үшін дисульфидтік байланыстың қажеті жоқ.
Амин қышқылдарының барлық жеке арақатынасының нәтижесі, көптеген белок молекулалары өздеріне тән конформацияға спонтанды түрде келе береді: әдетте қомақты глобулалық, бірақ ішінара фибриллярлық созылған пішінде де болады. Глобуланың ортасы бүйірлік гидрофобтық топтармен тығыз қапталған, кристалдың ішіндегідей, ал полярлық бүйірлік топтар күрделі және тұрақсыз сыртқы қабатты құрайды. Белоктың кіші молекулалармен және басқада молекулалардың үстіңгі қабаттарымен байланысу ерекшелігі әртүрлі адамдардың осы күрделі беткі қабатта орналасуына және химиялық қасиеттеріне байланысты. Химиялық тұрғыдан белоктар - белгілі молекулалардың ішіндегі ең күрделілері.
Тізбектің бір-біріне ұқсас жиылу тәсілі әртүрлі белоктарда үнемі қайталанып отырады. Полипептидтік тізбектердің амин қышқылдарының бірізділігінде олардың түйінделуіне қажетті ақпарат болса да, ол ақпараттың қалай оқылатыны әлі белгісіз, сондықтан белоктың кеңістіктегі болашақ құрылымын бірізділік бойынша дәл болжайтын мүмкіндік жоқ. Сондықтан, белоктың табиғи конформациясын белок кристалдарын анықтайтын өте күрделі рентгенқүрылымдық талдау әдісінің көмегімен ғана табады. Бұл әдісті қолдану арқылы осы күнге дейін 200-ден артық белок талданған. Лизоцим белогының мысалында көлемді немесе қаңқалық үлгісі арқылы белоктың толық құрылымын көруге болады.
Әртүрлі белоктардың кеңістіктегі құрылымын салыстыра отырып, әрбір белоктың конформациясы бірегей болғанмен де макромомолекуланың жеке бөлшектерінде тізбектің орам түрлері үнемі қайталанып отыратындығы анықталды. Әсіресе, жиылудың (орамның) екі жолы жиі кездеседі, себебі олар пептидтік топтардың өздерінің арасындағы сутектік байланыстардың дұрыс ұйымдасуынан болады, ал оған бүйірлік тізбектердің арақатынасының бірегейлігінің қатынасы жоқ. Осы екі тәсіл де 1951 ж. үлгінің көмегімен жібек және шашқа жасалған рентгенқүрылымдық талдау нәтижесіне негізделіп, дұрыс болжанған болатын. [1]
Қазір бұл кезендік кұрылымдарды β-қатпарлы қабат және а- шиыршық деп атайды. В-катпарлы конформацияда жібек фибрионының а- кератинде тері және оның туындыларының (шаш, тырнақ және қауырсындар) белогында кездеседі. β-қатпарлы қабаты құрылымы көптеген глобулалық белоктардың өзегінің басым бөлігін құрайды. β-қабат полипептидтік тізбектің β-құрылымдық β-бөлшектері деп аталатын екі, не одан да көбірігінен құралады.
Белоктардың кәдімгі полимерден өзгешілігі - құрылымы жағынан бір-біріне ұқсас болғанмен де, мүлдем бірдей емес мономерден құралған. Белоктың мономерлері, жоғары айтылғандай, амин қышқылдары. Әрбір амин қышкылының ерекше құрылысы, қасиеті және аты болады.
... жалғасыСіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz