Метаболизм және жасушалық энергетика: анаболизм, катаболизм, АТФ синтезі және фотосинтез

Метаболизм- тірі организмдер тіршілігінің негізі
Кіріспе
Клеткаларда үздіксіз биологиялық синтез процестері жүреді. Ферменттердің көмегімен
төменгі молекулалық заттардан жоғары молекулалы қосылыстар түзіледі: амин
қышқылдарынан - белоктар; моносахаридтерден - күрделі көмірсулар, азотты
негіздер мен қанттардан - нуклеотидтер, ал нуклеотидтерден - нуклейн қышқылдары
синтезделеді. Биосинтез реакцияларының жиынтығы - пластикалық алмасу немесе
ассимиляция деп аталады, ал синтезге қарама - қарсы процесс диссимиляция - ыдырау
реакцияларының жиынтығы болып табылады. Жоғары молекулалы қосылыстардың
ыдырауы кезінде, биологиялық синтез реакцияларына қажетті энергия бөлінеді.
Биосинтездік реакциялар өзінің жүру реті және даралығымен ерекшеленеді.
Синтезделетін үлкен органикалық молекулалардың құрылысы ДНҚ - дағы нуклеотидтердіњ
орналасу ретімен, яғни генотиппен анықталады. Зат алмасу процестері тіршілік ету
жағдайының үнемі өзгеруіне қарамастан организмнің ішкі ортасының тұрақтылығын -
гомеостазды - қамтамассыз етеді.
Гомеостаз, синтез процестері - пластикалық алмасу мен ыдырау процестерін
қамтамассыз етеді, мұндай реакциялардың нәтижесінде АТФ - тiң макроэнергиялық
байланыстарында жиналатын энергия босап шығады. Синтезделген заттар өсу
процесінде клеткалар мен олардың органоидтарын құруға және пайдаланылған немесе
бұзылған молекулаларды ауыстыруға жұмсалады. Синтездің барлық реакциялары
энергияның сіңіру арқылы жүреді.
1-тақырып: Анабализм
Биологиялық синтез реакцияларының жиынтығын пластикалық алмасу немесе анаболизм (грек тілінен anabole - көтерілу) деп аталады. Мұндай алмасу түрінің осылай аталуы оның мәнінін көрсетеді: клеткаға сырттан түсетін қарапайым заттардан, клетка заттарына ұқсас заттар түзіледі, яғни ассимиляция процесі жүреді.
Клетка мен біртұтас организмдегі барлық метаболизм процестері тұқымқуалаушылық аппараттарының бақылауымен жүргізіледі. Осы процестердің барлығы клеткадағы генетикалық ақпараттың жүзеге асырылуының нәтижесі.
Пластикалық алмасу кезіндегі тұқымқуалаушылық ақпараттың жүзеге асу процестерінің ішіндегі маңыздыларының бірі - белок биосинтезі.
Тұқымқуалаушылық ақпараттың жүзеге
асырылуы - белоктар биосинтезі.
Жоғарыда айтылғандай белок молекулалары қасиеттерінің алуан түрлілігі бірінші реттік құрылым, яғни амин қышқылдарының орналасу ретімен анықталады.
Белок синтезделу үшін оның бірінші реттік құрылымындағы барлық амин қышқылдарының орналасу реті туралы ақпарат рибосомаларға жеткілізілуі тиіс. Бұл процесс екі кезеңнен тұрады: транскрипция жєне трансляция.
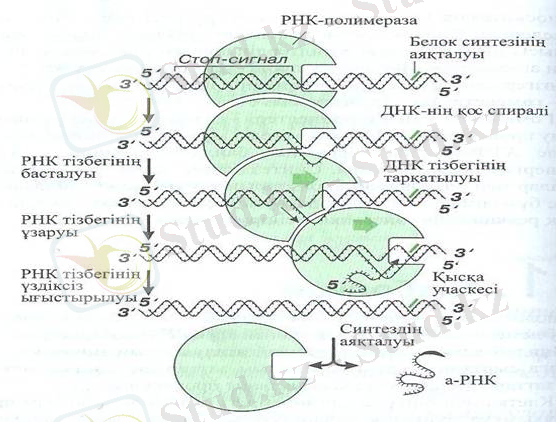
1-сурет. Транскрипция
Транскрипция. (1-сурет) . Ақпараттың транскрипциясы (латын сөзінен transcriptio - көшіру), яғни көшіру - РНК нуклеотидтерінің орналасу реті, матрицалық ДНК тізбегі нуклеотидтерінің орналасу ретіне (комплементарлы заң) сәйкес, бір тізбегі РНК молекуласы, ДНК молекуласының полинуклеидтік бір тізбегінде синтезделуі арқылы жүзеге асады. Синтездің бастапқы нүктесін «танудың», ақпарат оқылатын ДНК тізбегін таңдауды, процестің аяқталуын анықтайтын арнайы механизмдер болады. Ақпараттық РНК осылайша түзіледі.
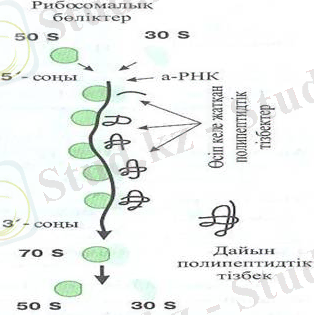
2-сурет. Трансляция
Трансляция (2-сурет) . ( латын сөзінен translation - аудару, беру) . Биосинтездің келесі кезеңі а-РНК молекуласындағы нуклеотидтердің орналасу реті туралы ақпаратты - полипептид тiзбегіндегі амин қышқылдарының орналасу ретіне аудару процесі - трансляция болып табылады.
Қалыптасқан ядросы жоқ организмдер прокариоттарда (бактериялар мен көк жасыл балдырларда) - рибосомалар, жаңадан синтезделіп, ДНЌ молекуласынан бөлінген ақпараттық РНЌ - мен бірден байланыстырады немесе синтез толық аяқталғанға дейін қосылып үлгереді. Ал эукариоттарда а-РНЌ алдымен ядро қабықшасы арқылы цитоплазмаға жеткізілуі тиіс. а-РНЌ цитоплазмаға арнайы белоктармен жеткізіледі, бұл белоктар РНК молекуласымен комплекс түзіеді. а-РНҚ-ны рибосомаларға жеткізу қызметінен басқа, бұл белоктар а-РНҚ-ны цитоплазмалық ферменттердің бұзу әсерінен де сақтайды. Цитоплазмада а-РНҚ бір ұшына (ядрода молекула синтезі басталған ұшына) рибосома орналасады, осылайша полипептидтің синтезі басталады.
Рибосома а-РНҚ молекуласы бойымен жайлап жылжып жүрмейді, керісінше, алдымен, триплеттен триплетке секіріп қозғалады. Рибосоманың а-РНҚ молекуласы бойымен қозғалуы кезінде полипепдтік тізбекке амин қышқылдары бірінен кейін - бірі а-РНҚ триплеттеріне сєйкес қатарласып, орналаса бастайды. Амин қышқылыныњ а-РНҚ триплетінің кодына тура сәйкес келуін транспорттық РНҚ (т-РНҚ), қамтамасыз етеді. Әр амин қышқылының өзінің жеке т-РНҚ-сы болады, бұл т-РНҚ-ның үш триплеттерінің біреуі - антикодон - а-РНҚ-ның нақты бір триплетіне комплементарлы болып келеді. Дәл осылай әрбір амин қышқылына оны т-РНҚ-ға байланыстыратын өзінің ферменті сәйкес келеді.

3-сурет. Тұқым қуалау ақпаратының ДНҚ-дан а-РНҚ-ға және белокқа берілу сызба нұсқасы.
Полипептидтік тізбек биосинтезінің процесі кезінде белок молекулаларының құрылысы турады тұқымқуалаушылық ақпараттың берілуінің жалпы принципі
(3-сурет) бейнеленген.
Синтез аяқталғаннан кейін полипептидтік тізбек матрица - а-РНК молекуласынан - бөлінеді, спиральға оралады, одан кейін белгіленген белокқа сай үшінші реттік құрылымға ие болады.
а-РНК молекуласы рибосома сияқты полипептидтерінің синтезіне бірнеше рет қолданыла алады. Бұл параграфта трансляция мен транскрипцияға өте қарапайым түрде сипаттама берілген. Есте сақтайтын жәйт, белоктардың биосинтезі көптеген ферметтердің қатысуымен және пептидтік байланыстарды түзетін энергиядан асып түсетін, орасан көп энергия мөлшерінің жұмсалуымен жүретін өте күрделі процесс. Биосинтез жүйесінің керемет күрделілігі мен оның көп энергияны қажет етуі полипептидтер синтезінің өте жоғары дәлдікпен және нақты тәртіппен өтуін қамтамасыз етеді. Клеткадағы белокты емес молекулалардың синтезі үш кезеңнен тұрады. Алғашында ерекше белок - ферменттің құрылысы туралы ақпарат жүзеге асырылады, одан кейін бұл ферменттердің көмегімен белгілі бір көмірсудың немесе майдың молекуласы түзіледі. Осыған ұқсас жолмен басқа молекулалар: витаминдер, гормондар жєне т. б. түзіледі.


2-тақырып. Энергетикалық алмасу - катаболизм
Синтезге қарсы процесс диссимиляция - ыдырау реакциясының жиынтығы болып табылады. Жоғарғы молекулалы қосылыстардың ыдырау кезінде биосинтез реакциясына қажетті энергия бөлінеді. Сондықтан диссимиляцияны клетканың энергетикалық алмасу немесе катаболизм (грек тілінен katabole - қирау, бүлдіру) деп атайды.
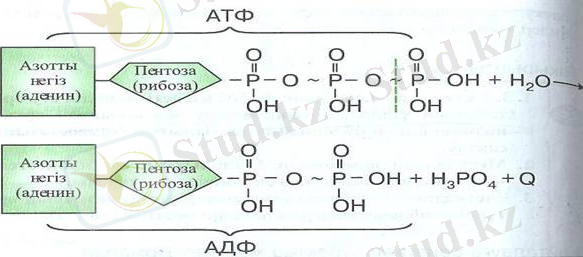
4-сурет. АТФ құрылысы мен оның АДФ-ға айналу сызба нұсқасы.
Қоректік заттардың химиялық энергиясы органикалық қосылыстар молекулаларындағы атомдар арасындағы әр түрлі ковалентті байланыстарда жинақталған. Мысалы, пептидтік тәрізді химиялық байланыстың үзілуі кезінде 1 мольге шаққанда 12кДж энергия босап шығады. Глюкозаның С, Н және О атомдарының байланыстары арасындағы жинақталған потенциалды энергияныњ мөлшері 1 мольге (яғни 180г глюкозаға) шаққанда 2800 кДж - тең. Бірқатар ферменттер қатысуымен глюкозаның ыдырауы кезінде энергия келесі теңдеуге сәйкес біртіндеп бөлініп шығады:
С6Н12О6+ 6О2
 6Н2О + 6СО2 + 2800 кДж
6Н2О + 6СО2 + 2800 кДж
Қоректік заттардан бөлініп шығатын энергияның бір бөлігі жылу ретінде таралады, ал басқа бөлігі энергияға бай АТФ-тың фосфаттық байланыстарында жиналады.
Сондықтан, АТФ клеткалық функциялардың барлық түрлерін энергиямен қамтамасыз етеді. Бұл функцияларға биосинтез, механикалық жұмыс (клетканың бөлінуі, бұлшық еттердің жиырлуы), заттардың мембраналардан өтіп белсенді түрде тасымалдануы, жүйке импульсын өткізу процесі кезіндегі мембраналық патенциалды ұстап тұру, әртүрлі түзінділердің бөлінуі жатады. АТФ молекуласы адениннің азоттық негізінен, рибоза қантынан және фосфор қышқылының үш қалдығынан тұрады, (4-сурет) . Аденин, рибоза және бірінші фосфат аденозинмонофосфатты (АМФ) түзеді. Егер бірінші фосфатқа екіншісі қосылатын болса, аденозиндифосфат (АДФ) түзеледі. Фосфор қышқылының үш қалдығы бар молекула (аденозинтрифосфат-АТФ) энергияға милінше бай. АТФ-тың соңғы фосфатының ажырап, түсіп қалуы кезінде 40 кДж энергия бөлінеді, салыстыру үшін: жай химиялық байланыстардың үзілуі кезінде тек 12 кДж энергия бөлінеді. АТФ молекулаларындағы энергияға бай байланыстардың көмегімен клетка өте кішкене кеңістікте энергияның көп мөлшерін жинай алады және оны қажет болған жағдайда мөлшерлеп қана пайдалана алады. АТФ синтезі митохондрияларда жүзеге асырылады. Бұл жерден АТФ молекулалары клетканың әр түрлі бөліктеріне беріліп отырады. Осылайша клетканың тіршілік әрекеті процестеріне қажетті энергия жеткізіліп отырады.


3-тақырып: Энергетикалық алмасу кезеңдері.
Әдетте энергетикалық алмасу процесін үш кезеңге бөледі. Бірінші кезең - дайындық кезеңі. Бұл кезеңде ди- және полисахаридтердің, майлардың, белоктардың молекулалары кіші молекулаларға - глюкозаға, глицеринге, май қышқылдарына, амин қышқылдарына ыдырайды; ал нуклеин қышқылдарының ірі молекулалары нуклеотидтерге ыдырайды. Бұл кезеңде энергияның аз мөлшері бөлінеді және осы энергия жылу түрінде таралады.
Екінші кезең - оттексіз немесе толық емес кезең. Ол анаэробты тыныс алу(гликолиз) немесе ашу процесі деп аталады. «Ашу» термині әдетте микроорганизмдердің немесе өсімдіктердің клеткаларында жүретін процестерге қатысты пайдаланылып айтылады. Бұл кезеңде ферменттердің қатысуымен түзілетін заттар одан әрі ыдырайды. Мысалы, бұлшық еттерде анаэробты тыныс алу нәтижесінде глюкоза молекуласы пирожүзім қышқылының (С 3 Н 4 О 3 ) екі молекуласына ыдырайды, олар одан әрі сүт қышқылына (С 3 Н 6 О 3 ) дейін тотықсызданады. Глюкозаның ыдырау реакцияларына фосфор қышқылы мен АДФ қатысады. Бұл реакцияныњ жалпы түрі мынадай:
С
6
Н
12
О
6
+ 2Н
3
РО
4
+ 2АДФ
 2С
3
Н
6
О
3
+ 2АТФ + 2Н
2
О
2С
3
Н
6
О
3
+ 2АТФ + 2Н
2
О
Ашытқы саңырауқұлақтарында глюкоза молекуласы оттегінің қатысынсыз этил спирті мен көміртегінің қос тотығына айналады (спирттік ашу) :
С
6
Н
12
О
6
+ 2Н
3
РО
4
+ АДФ
 2С
2
Н
5
ОН + 2СО
2
+ 2АТФ + 2Н
2
О
2С
2
Н
5
ОН + 2СО
2
+ 2АТФ + 2Н
2
О
Басқа микроорганизмдер гликолиз ацетонның, сірке қышқылының және т. б. түзілуімен аяқталуы мүмкін.
Барлық жағдайларда глюкозаның бір молекуласының ыдырауы АТФ-тың екі молекуласының түзілуімен жүреді. Глюкозаның оттексіз ыдырауы барысында АТФ молекуласында химиялық байланыс түрінде энергияның 40% - ы сақталады, ал қалғаны жылу түрінде тарап кетеді.
Энергетикалық алмасудың үшінші кезеңі - аэробты тыныс алу немесе оттекті ыдырау кезеңі. Энергетикалық алмасудың бұл кезеңіндегі реакцияларда ферменттермен катализденеді. Клеткадағы алдыңғы кезеңде түзілген заттар оттегі қатысында соңғы заттарға - Н 2 О, СО 2 - дейін тотығады. Оттекті тыныс алу энергияның көп мөлшерінің бөлінуімен және оның АТФ молекулаларында жиналуымен жүреді. Аэробты тыныс алудың жинақтық теңдеуі мынадай:
2С
3
Н
6
О
3
+ 6О
2
+ 36Н
3
РО
4
+ 36АДФ
 6СО
2
+ 38Н
2
О + 36АТФ
6СО
2
+ 38Н
2
О + 36АТФ
Сонымен, сүт қышқылының екі молекуласының тотығуы кезінде АТФ - тыњ 36 молекуласы түзіледі. Демек, клетканы энергиямен қамтамасыз етудегі негізгі рөлді аэробты тыныс алу атқарады.
Энергияны қабылдау тәсіліне байланысты барлық организмдер екі топқа - автотрофты және гетеротрофты болып бөлінеді.


4-тақырып : Лимон қышқылы айналымының сатылары. Пируваттың өзгеруі.
Лимон қышықылы айналымына даярлық.
Лимон қышықылының айналымына пируват
молекуласының өзгеруінен басталады. Атап айтқанда, пируват көміртектің (
3
С) бір атомын СО
 түрінде
түрінде
жоғалтатыны сызбанұсқада бейнеленген.
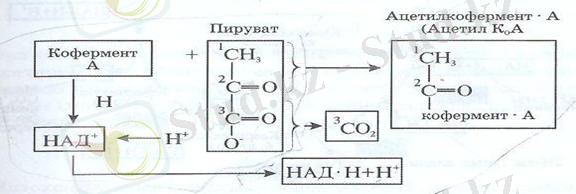
Осы кезде кофермент «А» пируватының қалған екі көміртек ацетил тобын қосып алады да, лимон
қышқылы айналымының алғашқы сатысына өтеді.
НАД
+
пируват пен конферемент А - дан 2Н - ты қосып алып, НАД Н + Н
+
- ке айналады. Реакция нәтижесінде түзілген ацетил К
 А лимон қышқылының сатысына өтеді де, лимон қышқылы айналымына
А лимон қышқылының сатысына өтеді де, лимон қышқылы айналымына
даярлық аяқталады. (Сызбанұсқада) Даярлық аяқталғаннан соң (5-суреттен) көріп отырғанымыздай лимон
қышқылының айналым реакциясы басталады. Ол реакция сағат тілінің бағытымен жүреді. Реакция
бастапқы зат ретінде ацетил К
 А қатысады. Ол өзіндегі екі көміртекті ацетил тобын төрт көміртекті
А қатысады. Ол өзіндегі екі көміртекті ацетил тобын төрт көміртекті
қосылыс - қымыздық сірке қышқылына беріп, алты көміртекті қосылыс - лимон қышқылына айналады.
(5-сурет) . Бұл реакция - ферменттердің жәрдемімен үздіксіз жүріп отыратын күрделі процесс. Лимон
қышқылы(5-суреттен) көріп отырғанымыздай екі көміртек атомын СО
 түрінде бөліп шығарады да, қалған
түрінде бөліп шығарады да, қалған
төрт көміртек атомынан жаңа қымыздық сірке молекуласы түзіліп отырады. (5-суретте. ) Лимон қышқылы
айналымының қысқартылған түрі берілген.
Лимон қышқылының айналымын 1930 жылдары ағылшын биохимигі Г. Кребс. ашты. Сондықтан бұл процесс Г. Кребстің атымен аталады. Осы еңбегі үшін Г. Кребс 1953 жылы Нобель сыйлығының лауреаты атағын алды.
Қорытып айтқанда, лимон қышқылы айналымының нәтижелері мынадай:
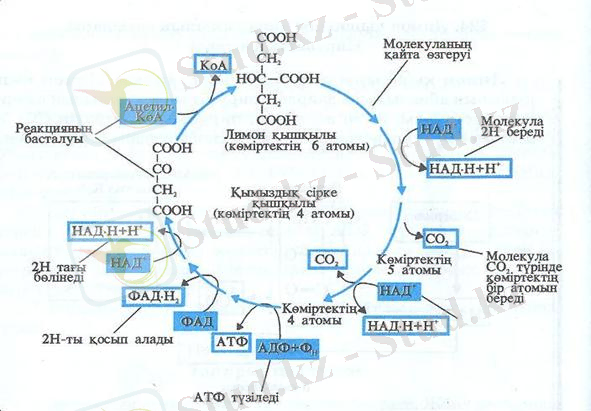
5-сурет. Лимон қышқылыныњ айналымы.
1. Біз тыныс алу мүшелеріміз арқылы СО
 өнімдерін бөліп шығарамыз. Ол өнімдер пируват молекуласы реакцияға түскен кезде пайда болады.
өнімдерін бөліп шығарамыз. Ол өнімдер пируват молекуласы реакцияға түскен кезде пайда болады.
2. Сутек атомдары лимон қышқылдарының айналымының әр-бір сатысында ыдырайды да олар электрондардың айналымының акцепторларына беріледі. Осы уақытта НАД + жєне ФАД түзіледі . Бұлар қайтадан сутек атомдарын қосып алып, НАД·Н+Н + ЖӘНЕ ФАД·Н 2 - ге айналады.
3. Лимон қышқылының әр айналымында АТФ - тың бір молекуласы синтезделеді.
4. Лимон қышқылының айналымы мынадай жалпы теңдеумен қорытылады:
А) қымыздық сірке қышқылы → Ацетил К
 А түрінде жинақталған → АДФ + Ф
А түрінде жинақталған → АДФ + Ф
 → 3 НАД ∙Н
+
→ ФАД;
→ 3 НАД ∙Н
+
→ ФАД;
Є) реакция нәтижесінде мынадай заттар түзіледі: қымыздық сірке қышқылы +2СО
 + К
+ К
 А + АТФ + 3НАД∙Н + 3Н
+
+ ФАД·Н
2
.
А + АТФ + 3НАД∙Н + 3Н
+
+ ФАД·Н
2
.
Н
+
қоймасын толтыру. Электрондар тасымалдау тізбегі.
Реакцияның бастапқы заты ретінде алынған глюкоза ферменттердің жәрдемімен толық ыдырайды (глюкозаның оттекті ыдырауын еске түсіріңдер) . Осы кезде бөлінген энергия АТФ синтезіне қатысқанымен, оның негізгі бөлігі электрондармен толықтырылған тасымалдаушы молекулалар НАД · Н + Н
+
жєне ФАД · Н
 түрінде жинақталады.
түрінде жинақталады.
Бұл молекулалар гликолиз бен лимон қышқылы айналымының өнімдері ескерген жөн. Тасымалдаушы молекулалар (6-суреттен) көріп отырғанымыздай, сутектің атомдарын электрондар тасымалдау тізбегіне береді де, қайтадан сутектің жаңа атомдарын тасымалдауға дайын тұрады. Бұл процесс үздіксіз жүріп жатады.
Сутек атомдары митохондрияның ішкі мембранасы өтіп, ферменттердің әсерінен тотығып, электрондарын жоғалтады:
--
Н 0 ― е →Н +
Бұл тотығудың нәтижесінде пайда болған сутектің электрондары мен катиондарын Н + - ті тасымалдайтын молекулалар қосып алып, мембрананың ішкі жағына өткізеді. (6-сурет) . Олар сол жерде оттекпен қосылады (митохондрияға молекула күйіндегі оттек сыртқы ортадан үздіксіз келіп тұрады) :
О
 ― 4 е → 2О
2 -
― 4 е → 2О
2 -
Митохондрияның сыртқы мембранасына Н + , ал ішкі жағына теріс зарядты бөлшектер, яғни аниондар жинақталады. Демек, мембрананың сыртқы және ішкі жағында қарама - қарсы зарядталған бөлшектердің саны арта түседі. Олардың санының артуына байланысты арасындағы потенциал айырмашылығы да артады.
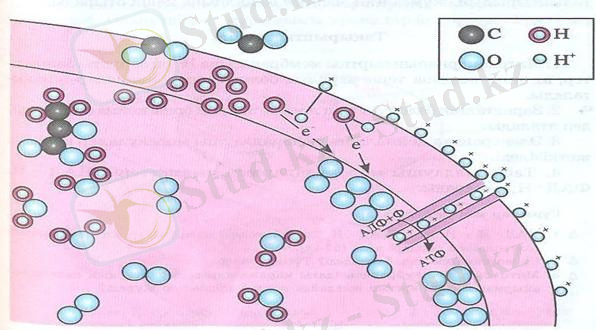
6-сурет. Митохондриядағы АТФ синтезінің сызбанұсқасы.
Қарама - қарсы зарядтардың потенциал айырмашылығы артқанда, энергия толы бөлшектер бір - бірімен қосылуға әрекет жасайтынын физика курсынан білесіңдер.
Мембрананың кейбір жерлерінде АТФ - ті синтездейтін фермент молекулалары орналасқан. Ол ферменттердің молекуласында Н + катиондары өте алатын каналдар болады. Мембрананың сыртындағы катиондары Н + потенциал айырмашылығы шамадан тыс көбейіп кетеді. Осы уақытта электр өрісінің күшімен оң зарядталған бөлшектер. Н + АДФ ферменттерінің каналдары арқылы мембрананың ішкі жағына өтеді де, оттекпен әрекеттеседі. Нәтижесінде су малекуласы түзіледі. :
4Н
+
+ О
 → 2Н
→ 2Н
 О
О
Электрон тасымалдаушы тізбек мембрананың ішкі қабатында, ал Н
+
қойма (акцептор) ішкі және сыртқы мембраналар арасында болады. Н
+
иондарының қоймаға толуына байланысты энергия жинақталып, АТФ синтезіне жұмсалады. Мысалы 6-суретте бейнеленгендей, мембрананың түтікшелері арқылы сырттан іш жағына Н
+
иондары өтеді. Олардың энергиясы АДФ пен Ф
 - тың қосылуына жұмсалады. Бұл реакцияның нәтижесінде АТФ молекуласы синтезделеді. Осы жолмен АТФ синтезделген сайын Н
+
қоймасында жиналған энергия азая береді. . НАД ∙ Н және ФАД ∙ Н
- тың қосылуына жұмсалады. Бұл реакцияның нәтижесінде АТФ молекуласы синтезделеді. Осы жолмен АТФ синтезделген сайын Н
+
қоймасында жиналған энергия азая береді. . НАД ∙ Н және ФАД ∙ Н
 - ден электрон тасымалдау тізбегі арқылы келген электрондардың есебінен қойма үнемі Н
+
ионымен толықтырылып, жұмсалған энергия өз қалпына келіп отырады.
- ден электрон тасымалдау тізбегі арқылы келген электрондардың есебінен қойма үнемі Н
+
ионымен толықтырылып, жұмсалған энергия өз қалпына келіп отырады.
Сіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz