Прокариоттар мен эукариоттардағы ген экспрессиясының молекулалық реттелу механизмдері

13
гендер экспрессиясының реттелуі
Гендер экспрессиясының дифференциалды реттелуін жүзеге асыру қабілеті прокариоттар мен эукариоттардың негізгі қасиеті болып есептеледі. Қандай гендердің экспрессиялануы керек, қайсыларына керегі жоқ екендігіне бақылау жасай отырып, сол сияқты түрлі гендер экспрессиясының деңгейін реттей келе клеткалар өз фенотипін белгілі бір сыртқы және ішкі орта жағдайларына бейімдей алады. Гендер көбінесе біртіндеп экспрессияланады: бір геннің активациясы басқа бірнеше гендердің экспрессиясын тудыра алады. Қайсыбір гендер немесе туыстас гендердің тобы үйлесімді түрде экспрессияланады, яғни реттеуші сигналға бір мезгілде және бірдей деңгейде жауап береді. Синтездің бір белоктан басқаларына ауысуы кез-келген дамудың, мейлі ол залалданған клеткадағы вирустардың репродукциясы, бактериялардағы спора түзілу, эмбриондардың дамуы немесе ұлпалардың жіктелуінің болсын негізін құрайды. Әртүрлі клеткаларда гендердің түрлі жиынтықтары транскрипцияланады. Бұл процестердің әр кезеңінде өзіндік ерекшелігі бар белоктар синтезделеді.
ДНҚ-дан белокқа алып баратын жолда мұндай бақылау кез-келген кезеңде жүзеге аса алады (13. 1-сурет) .
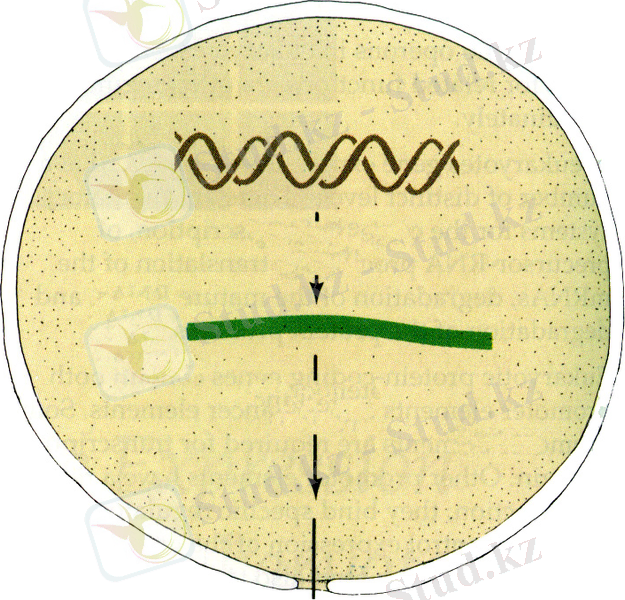
13. 1-сурет. Эукариоттарда гендер экспрессиясы бақылануының болуға тиісті деп ұйғарылған кезеңдерін көрсететін сызбанұсқа (11) .
Гендер экспрессиясының реттелуі мынандай деңгейлерде жүруі мүмкін:
- бақылау (геннің транскипциялану уақыты мен сипаты)
- бақылау (алғашқы транскрипт процессингінің сипаты)
- Транспортдеңгейіндегі бақылау (цитоплазмаға тасымалдауға арналған ядродағы жетілген мРНҚ-ны сұрыптау)
- бақылау (рибосомалардағы трансляция үшін қажетті цитоплазмадағы мРНҚ-ны сұрыптау)
- мРНҚ сұрыптау (цитоплазмадағы мРНҚ-ның белгілі бір түрлерін таңдап тұрақтандыру)
Транскрипция процесінде РНҚ құрамын реттеу генетикалық экспрессияны реттеудің аса маңызды және тиімді жолы болып табылады. Транскрипция деңгейіндегі бақылау әсіресе біршама тұрақсыз мРНҚ жағдайында аса қажет. Өз кезегінде посттранскипциялық құбылыстар да реттеліп отырады және клеткада РНҚ-ның функционалдық молекулаларының болуы да соған байланысты. Ген өнімдері экспрессиясының трансляциялық реттелуі рибосомаларда трансляцияланатын мРНҚ-ның сұрыпталуын қамтамасыз етеді. Ген әрекеттерін бақылау функционалдық деңгейде полипептидтердің посттрансляциялық модификациясы және ферменттер активтілігін аллостерикалық реттеу жолдарымен жүзеге асады. Эукариоттарда транскипция трансляциямен тікелей қатынаста болмайтындықтан цитоплазмадағы мРНҚ-ның белгілі бір түрлерінің деградациясы мен таңдамалы тұрақтануы деңгейіндегі гендер экспрессиясының цитоплазмалық бақылануы жүзеге асады.
Сонымен қатар гендер экспрессиясының реттелуі хроматин жағдайының, соның ішінде нуклеосомалар қатталуының өзгерісі жолымен жүруі мүмкін. Бұл экспрессия реттелуінің тағыда бір деңгейін туғызады. Нуклеосомалық реттеуші механизмдер бір ғана генге тән болмайды, көптеген гендердің экспрессиясына әсер етеді.
Бактерияларда көбірек зерттелген реттелудің транскрипциялық деңгейін біршама кеңірек қарастырайық.
Бактериялар қоршаған ортадағы өзгерістерге тез жауап беруге тиісті. Олардың өз тіршілігін сақтауы метаболизмді бір субстраттан екіншісіне ауыстыру қабілетіне байланысты, себебі, олардың тіршілік ортасына қоректік заттардың келіп түсуі тұрақты түрде өзгеріп отыруы мүмкін. Бактерия қандай да болмасын метаболиттік бағыттағы ферментті оған тиісті субстраты жоқ жағдайда синтездей алмайды, бірақ ондай жағдай бола қалса ферментті синтездеуін кез-келген уақытта бастап жіберуге қабілетті. Бактерия гендері қызметінің басты ерекшеліктері осы мен түсіндіріледі бұл олардың кластарға бірігуіне және экспрессия реттілігіне байланысты. Гендердің кластерлерге бірігетіндігі соншалықты, биосинтездің белгілі бір бағыты үшін қажетті ферменттердің барлығы бір-бірімен тіркескен гендер арқылы анықталады. Гендердің бүкіл тобы бір полицистронды (полигенді) мРНҚ-ға транскрипцияланады, ал одан әрі қарай біртіндеп рибосомаларда трансляцияланып түрлі белоктарды түзеді.
Прокариотты гендер экспрессиясының реттелуі. Оперон үлгісі
Прокариоттардағы генетикалық активтіліктің транскрипциялық бақылану құбылысын көрсетудің классикалық мысалына β -галактозидаза ферменті синтезінің реттелу жүйесін жатқызуға болады. E. сoli бактериялары көміртегі мен энергияның бірден-бір көзі ретінде лактозаны пайдалануы мүмкін, себебі олар лактоза дисахаридін галактоза мен глюкозаға дейін ыдырататын фермент β -галактозидазаны көп мөлшерде түзе алуға қабілетті. E. сoli- дегі лактозаны ыдырататын ферментті кодтайтын гендер қызметінің реттелу механизмдерін зерттеу 1961 жылы Франция ғалымдары Ф. Жакоб, Ж. Моно, және А. Львовқа құрылымдық гендер жұмысының үйлесімді түрде бақылануының моделін ұсынуға мүмкіндік берді, ол оперон моделі деп аталады. Бактериялардағы гендер активтілігінің реттелуі механизмдерін ашқандары үшін Ф. Жакоб, Ж. Моно және А. Львовқа 1965ж. Нобель сыйлығы берілді.
Оперон моделі бойынша қызметі жағынан бір-бірімен тығыз байланысты, полипептидтерді кодтаушы құрылымдық гендер тобының транскрипциясы екі бақылаушы элементтер - реттеуші-ген және оператор арқылы реттеледі. Құрылымдық немесе ферментативтік қызмет атқаратын клетка үшін қажетті белоктарды кодтайтын гендерді құрылымдық гендер деп атайды. Басқа гендердің экспрессиясын реттейтін белоктарды кодтайтын гендерді реттеушілер дейді. Оператор көбінесе транскрипцияның инициациясы үшін қажетті промотор мен құрылымдық гендердің аралығында шоғырланған құрылымдық гендерге келіп қосылатын нуклеотидтердің тізбегі болып табылады. Бір-бірімен тығыз байланысқан құрылымдық гендер, оператор және промотордан тұратын, үйлесімді түрде экспрессияланатын гендердің тобын оперон деп атайды. 13. 2-суретте E. сoli дің lac - оперон гендерінің құрылысы берілген, ол осы уақытқа дейінгі ең бір жақсы сипатталған оперон болып табылады. Осы lac -оперонға 6000 жұпқа шамалас ДНҚ нуклеотидтері сәйкес келеді.
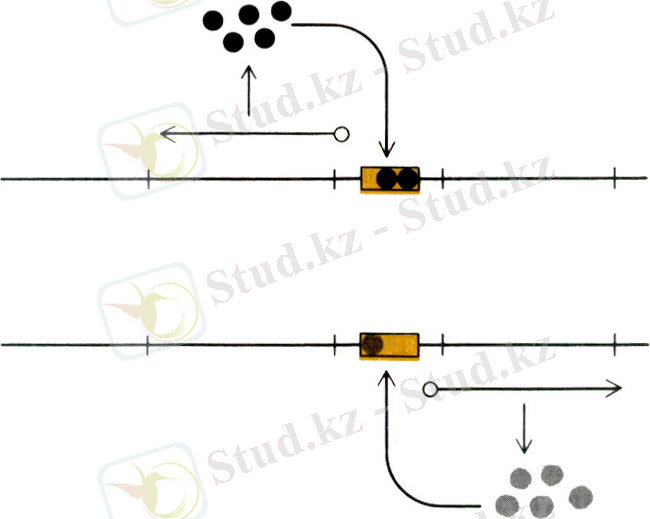
Реттеуші белоктар бұл қызметі ДНҚ-ғы белгілі бір сайттармен біріге отырып атқарады. Мұндай әсерлердің өзі негізінде реттелудің түріне байланысты болады. Егер реттеуші геннің өнімі белок - репрессор болып келсе, оның операторға келіп қосылуы құрылымдық гендердің транскрипциясына жол бермейді, сөйтіп РНҚ-полимеразаның промоторға қосылуына кедергі келтіреді. Керісінше, егер реттеуші белок қызметін активті апоиндуктор атқарса, оның операторға келіп қосылуы транскрипцияның инициациясына жағдай туғызады.

13. 2-сурет. E. сoli-дің lac - опероны (11) .
Реттеуші ген оперонның маңында немесе одан алысырақ орналасуы мүмкін. Мысалы, E. сoli- де галактозаның метаболизміне (алмасуына) жауапты ферменттерді кодтайтын үш ген - galE, galT және galK өзіне реттеуші сегмент-оператор арқылы келіп қосылатын промоторы бар оперонда орналасады. Арабинозаны пайдалану үшін қажетті ферменттердің синтезі екі жеке транскрипциялық бірліктермен реттеледі.
Оперонның жұмысын реттеуге сол сияқты төменгі-молекулалы заттар- эффекторлар да қатысады, олар оперонның құрылымына енетін құрылымдық гендердің индукторы не корепрессоры есебінде болады.
Эффектор-молекулалардың өз жұмысына әсер ету жолына қарай оперондарды индукциялаушы және репрессиялаушы деп ажыратады. Индукциялаушы оперондарда эффектор белок репрессорға келіп қосылады да оның оператормен бірігуіне жол бермейді. Сөйтіп құрылымдық гендердің транскрипциясына кедергі келтіреді. Оперон жұмысы реттелуінің мұндай түрін негатитвті деп атайды. Сонымен қатар индукцияланатын оперондар реттелудің позитивті бақылауында да бола алады, ол кезде эффектор реттеуші белокпен байланысып, оны активтендіреді. Позитивті реттелу арнайы белоктарды промотор аумағында орналасқан нуклеотидтік тізбектермен байланыстыру арқылы жүзеге асады. Активті апоиндуктор операторға келіп қосылып оперонның транскрипциялану мүмкіндігін тудырады. Реттелуді бақылаудың екі түрі репрессияланатын оперондар тұрғысынан да әсер етеді.
E. сoli lac - оперонының экспрессиясының реттелуі
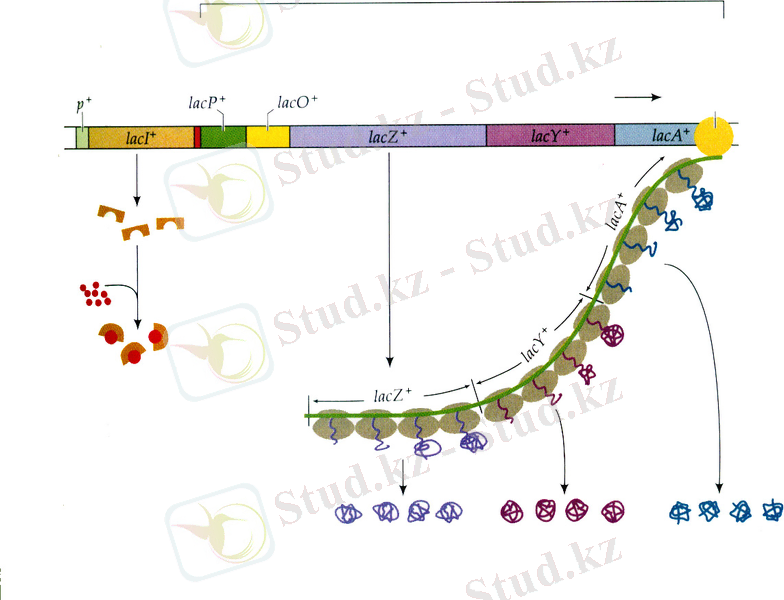
β -галактозидазаның ( lacZ ) синтезіне жауапты ген индуцибельді деп аталады, себебі ол кодтайтын фермент тек клеткада лактоза болғанда ғана синтезделеді. Екінші ген lacY - клеткаға лактозамен қоса қанттарды тасымалдауды қамтамасыз ететін галактозидпермеазаны кодтайды. Үшінші ген lacA - β -галактозид - трансацетилазаны кодтайды, ол фермент ацетильді топты ацетил коэнзимА-дан В-галактизидке алып барады, сөйтіп бактерияның өсуіне қолайлы жағдай туғызады. Осы үш генде - lacZ , lacY және lacA -да lac -оперонмен кодталынатын белоктар жайлы барлық ақпарат болады. Бұл үш lacZ, -Y, -A гендерінің кластері lacZ геніне жақын тұрған промотордан бір мРНҚ түрінде транскрипцияланады. Барлық үш ферментте әдетте бактерия клеткаларында шамалы мөлшерде болады. Алайда, бактерияларды энергияның бірден-бір көзі лактоза болатын ортада өсірген кезде аталған ферменттердің мөлшері 1000 есеге дейін артады.
Сол жақ бөлігінде өзінің жеке промоторы бар lacI гені орналасады және оның терминатор да болуы мүмкін. LacI генінің кодтаушы бөлігінің ұшы тікелей lacP - промоторымен түйіседі. Операторға lacZ генінің алғашқы 26 жұп негіздері сәйкес келеді. LacZ генінен соң lacY, lacA гендері мен терминатор орналасады.
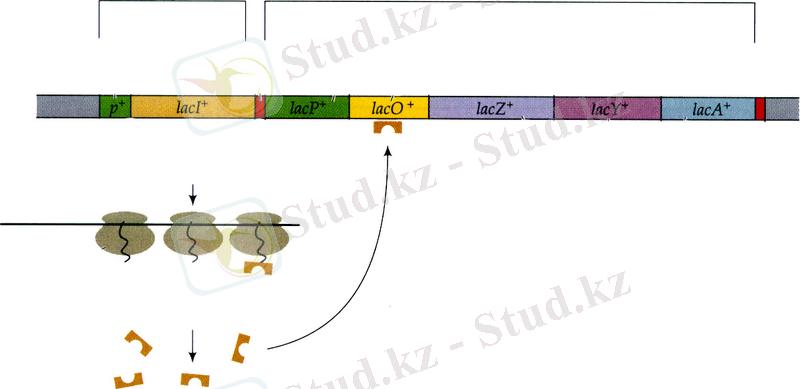
келіп қосылады.
13. 3-сурет. E. colі-дің lac оперонының қоректік ортада лактозаның жоқ кезіндегі функциональдық жағдайы (11) .
LacZ және lacY гендерінің мутациясы клетка лактозаны пайдалана алмаған кезде Lac фенотипін береді. LacZ мутанттарында β -галактозидазаның ферменттік активтілігі болмайды. LacY гені бойынша мутанттар сыртқы ортадан лактозаны өзіне сіңіре алмайды.
Лактозалық оперонның қызметі мынандай жолмен жүреді: lacZ, lacY және lacA гендерінің транскрипциясы lacI генімен кодталатын белок репрессормен бақыланады. LacZ, lacY және lacA гендері негатативті түрде бақыланады, яғни транскрипция белок-репрессордың көмегімен тоқтап қалғанша жүре береді. Репрессор геннің мутациясы lacZ, lacY және lacA гендерінің тұрақты транскрипциялық активтілігін тудырады. Lac - репрессор белогы оператормен lacZ, -Y, -A кластерінің транскрипциясы басталар кезде байланысып РНҚ-полимеразаның қозғалысы мен транскрипциясының инициациясын жояды (13. 3-сурет) .
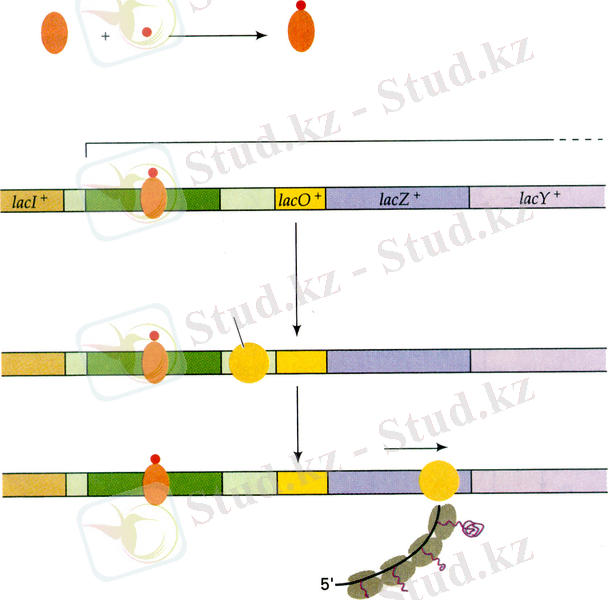
Белок репрессорда біреуі индуктормен, ал екіншісі оператормен байланысатын екі сайты болады. Индуктор (лактоза) клеткаға барып өз сайтымен байланысқан кезде, белоктың конформациясы өзгереді де басқа сайттың байланыстырғыш қабілеті бұзылады. Нәтижесінде репрессор транскрипцияны ұстап тұра алмайтындықтан ол басталып кетеді (13. 4-сурет, А) . Индуктор істен шыққан кезде репрессор оператордағы өз орнына қайтадан ие болып, транскрипцияны тоқтатады.
13. 4-сурет. E. coli-дің lac- оперонының позитивтік реттелуі (11) .
Қанттардың метаболизміне қатысатын ферменттердің синтезіне бақылау жасайтын
lac-
оперонының және тағы да басқа индуцибелді оперондардың экспрессиясы үшін тек оперонның репрессиясын алып тастау ғана емес, қайсы бір сигналды алу да қажет. Ондай сигнал үшін циклді АМФ-ты (цАМФ) катаболизмнің активаторы белогымен (САР, ағылшынша
catаbolite activator protein
) түзетін комплексі қызмет атқарады. Ол
lac
-промотордың ең басынан басталатын ерекше тізбектермен байланысады. САР-тың өзі молекулалық массасы 22 кДа болатын біртекті полипептидті тізбектердің димері болып табылады. Промотордың бас жағында САР-цАМФ комплексін ерекше жүйемен байланыстыру
lac
-оперон транскрипциясын 50 есеге жуық арттырады. САР өздігінен мұндай байланыстыруға және транскрипцияны жеделдетуге қабілетсіз. САР және цАМФ-ті реттейтін барлық гендердегідей, промотордың екі аумағы болады: РНҚ-полимеразамен байланыстыратын бөлімшесі және САР-цАМФ комплексімен байланыстыратын бөлімшесі
 (13. 5-сурет) .
(13. 5-сурет) .
13. 5-сурет. САР-цАМФ-тың қатысуымен lac- оперонының қызметінің реттелуі (11) .
Микроорганизмдердегі гендердің оперондық құрылымының негізгі ерекшелігі активтіліктің реттелуінің үйлесімділігі болып табылады, яғни бүкіл гендер экспрессияланса да немесе экспрессияланбаса да бірге қосылып қызмет атқарады. Матрицалық РНҚ-ның трансляциясы 5′- ұшынан біртіндеп жүзеге асырылады. Бұл неге индукция кезінде алдымен β- галактозидаза, содан соң β -галактозидпермеаза және ең соңында, β -галактозид-трансацетилаза пайда болатындығын түсіндіреді. Сол сияқты біртұтас мРНҚ-ның көмегімен үш геннің көрініс беруінің өзі барлық үш ферменттің салыстырмалы шамасы индукцияға қарамастан неге бірдей болып қалатындығын түсіндіреді.
Бір-біріне жақын бірнеше функцияларды кодтайтын гендер барлық жағдайда біртұтас оперон түзе бермейді. Мысалы, 30S және 50S-рибосомалық белоктарды кодтайтын гендер көптеген оперондарды құрайды.
Эукариоттардан оперондар табылмаған.
Триптофан оперонының экспрессиясының реттелуі
Оперон гендер тобын үйлестіре реттеуге мүмкіндік беретін құрылым болып есептеледі. Сонымен қатар жекелеген гендердің әртүрлі дәрежедегі экспрессиясын қамтамасыз ететін бақылаудың басқа да тәсілдері болуы мүмкін. Мысал ретінде триптофан оперонының реттелуін қарастыруға болады.
Триптофан E. сoli -де бес түрлі белоктан тұратын ферменттік комплекспен катализденетін бірінен соң бірі қатар келетін бес түрлі реакцияның нәтижесінде хоризм қышқылынан синтезделеді. Ферментативтік реакциялардың бұл тізбегі trp -оперонды құрайтын бес құрылымдық гендермен кодталады (13. 6-сурет) . TrpA және trpВ гендері өз кезегінде α- және β- триптофансинтетаза суббірліктерін анықтайды, trpЕ және trpD гендерінің өнімдері бірігіп антрацилсинтетаза ферментін түзеді, ал trpС гені кодтайды.
Trp -оперонымен кодталатын белоктар бірлескен түрде, шамамен бірдей мөлшерде промотормен транскрипцияланатын ұзындығы 7000 нуклеотидтерден тұратын полицистронды мРНҚ трансляциясы кезінде пайда болады. Ортада бактерияның өсуіне жеткілікті мөлшерде триптофан болған жағдайда E. coli клеткасы өте аз мөлшерде триптофан биосинтезіне қажетті ферменттерді түзе алады. Алайда егер клеткада экзогенді триптофан жоқ болса, онда барлық бес ферменттің де біршама қарқынды синтезі басталады. Клетка ішіндегі триптофанның мөлшеріне қарай бактерия клеткаларындағы фермент мөлшері 700 есеге дейін көбейе алады. Триптофандық оперонға талдау жасалғанда реттеуге промотор-оператор комплексінен бөлек тағы да басқа сайттың қатысатындығын көрсетті. Мұндай реттеуші сайт аттенуатор деп аталады. Ол 162 нуклеотидтен тұратын транскрипцияланатын басты тізбектің аумағында шоғырланған, ал бұл trpЕ генінің инициациялаушы кодонның алдында болады. Аттенуатор РНҚ-полимеразаның trp гендері аймағына қарай жылжуын қадағалайды және транскрипция үшін кедергі қызметін атқарады. Ол терминацияның Р-тәуелсіз сайты түрінде болады және терминациялық тізбектің құрамына ГЦ-ға бай аймақ енеді, ал олар терминаторлық полиндромды бізшелер (шпильки) түзеді. РНҚ-полимераза транскрипцияны осы бөлімшеде тоқтатады; нәтижесінде ұзындығы 140 негізден тұратын транскрипт пайда болады.
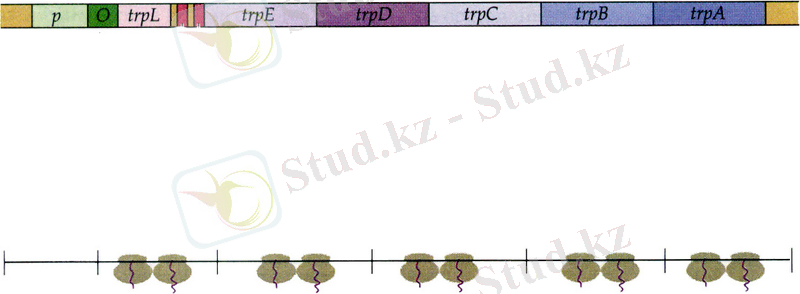
13. 6-сурет. E. coli-дің триптофан опероны. TrpL, trpЕ, trpD, trpС, trpВ және trpА құрылымдық гендерінің өнімдері және олардың көмегімен катализденетін реакциялар. Қысқартылған сөздер:PR-фосфорибозил, CdRP--фосфат (11) .
Бұл сайттағы терминация триптофанның болуына байланысты. Триптофанның деңгейі төмен болған жағдайда атенуаторлы тізбектің маңайындағы транскрипция терминациясы шектеледі де, РНҚ-полимераза құрылымдық гендерді транскрипциялауы әрі қарай жалғастыруға мүмкіндік алады. Триптофанның мөлшері жеткілікті болса терминация жақсы жүреді. Транскрипция терминациясының осылайша реттелуі аттенуация деп аталады.
Аттенуация E. сoli -дің және тағы да басқа бактериялардың көптеген гендері мен оперондарының экспрессиясының реттелуі кезінде қолданылады. Гистидиннің биосинтезіне қатысатын тоғыз ферментті кодтайтын оперон тек қана аттенуацияның көмегімен реттеледі. Олардың құрылымындағы ерекшелік мРНҚ-да терминациялаушы бізшелердің пайда болуына алып келетін полиндромның болуына байланысты. Промотордағы транскрипция инициациясы мен аттенуациясы нәтижесін өзгертетін репрессия механизмдерінің үйлесімі транскрипциялық деңгейдегі, сыртқы орта жағдайларына икемді ген экспрессиясы реттелуінің біршама тиімді болуын қамтамасыз етеді.
Гендер экспрессиясының фаг инфекциясы кезіндегі реттелуі
Фаг инфекциясы кезінде ген активтілігінің ауысуы ген қызметі реттелуінің тағы бір түріне жатады, ол жоғарыда қарастырылған оперондар жұмысының қосу және қайта алып тастау жолымен реттелуіне қарағанда өзгеше.
Бактериофагтардың ие-бактерияның метаболизмін фагтар репродукциясына ауыстыруына бірқатар мүмкіншіліктері бар. Ол фаг бөлшектері арқылы залалданған клеткаларда жаңа полимеразалардың, не олардың жеке суббірліктерінің, белок-репрессорлардың және белок активаторлардың пайда болуына байланысты. Лямбда (λ) бактериофагында тіршілік етудің екі альтернативті (қарама-қайшы) жолы бар. Бірінші - лизистік жолында барлық вирустық гендер белгілі бір уақыттық тізбекпен экспрессияланады, нәтижесінде шамамен жүздеген фаг бөлшектері пайда болып, залалды бактериялар ыдырайды. Инфекция тудыратын вирустардың дамуы екінші - лизогенді жолмен де жүруі мүмкін, ол кезде вирустық гендер ие-клеткасының ДНҚ-ның арнайы жерлеріне барып орналасады. Вирус геномының интеграцияланған түрі профаг деп аталады. Лизогенді клеткалардағы профагты ДНҚ клеткалық аппараттың көмегімен ондағы геномның бір бөлшегі сияқты болып бірнеше рет репликацияланады. Бірақ бұл жағдайда біреуінен басқа барлық гендер алынып қалады. Лизогенді клеткалар не кенеттен төменгі жиілікпен, не әртүрлі химиялық немесе физикалық факторлардың әсерінен лизистік тәсілге ауысады.
Лизистік цикл индукциясының табиғатына байланыссыз фагтың бұрынғы ″үндемейтін′′ гендері іске қосылып, инфекциялық фаг бөлшектерін тудыратын процестерді іске қосады.
13. 7-сурет. Лямбда (λ) фагтың инфекциядан кейінгі лизогендік (А) және лизистік (Б) даму жолдарын анықтайтын гендер транскрипциясы (8) .
Фагтардың көптеген геномдарының генетикалық картада орналасу реттілігі лизистік дамудың бір ізділігін дәл көрсете алады. Фагтар клеткалық метаболизмді екі топ гендердің көмегімен қайта өзгерте алады. Біреулері инфекциядан соң бірден-ақ көрініс береді, оларды алғашқы немесе қызметі ерте басталатын гендер деп атайды. Осындай гендердің бірінің өнімі бір гендердің іске қосылуын, ал басқалары өз кезегінде гендердің келесі ортаңғы және соңғы топтарының қызметтерін реттеуін қамтамасыз етеді. Гендердің біртіндеп көрініс беруінің осылайша реттелуі транскрипция арқылы жүреді. Мысалы, Т2, Т4, Т7 жағдайында гендер әсерінің осындай бірізділігі РНҚ-полимеразаның промоторлы ерекшелігінің модификациясы мен бақыланады.
λ фагта дамудың лизистік циклі лизогенияның тұрақтану циклімен түйіседі. Лямбда фагтың ДНҚ-сын жаңа клетка иесіне енгізгенде лизистік және лизогендік циклдер бірегей жолмен іске қосылады. 13. 7-суретте λ фагтың инфекциядан кейінгі лизистік және лизогендік даму жолдарын анықтайтын промоторлар мен операторлардың өзара бірігіп орналасуы көрсетілген.
Дамудың лизистік және лизогендік жолдарының арасындағы таңдау реттеуші белоктардың қорланып жиналу жылдамдығына байланысты олар: репрессорлы белок cI , антирепрессорлы белок Cro және реттеуші белоктар cII мен cIII . CI репрессоры мен Cro белогы активтілігінің арақатынасына қарай транскипцияның екі түрлі вариантының біреуі орнығады. Егер Cro -белоктың антирепрессиялық функциясы басым болып келсе, онда артықшылық лизистік жолға беріледі. Егер репрессия тұрақтанып үлгерсе, онда лизистік жолды жүзеге асыру үшін қажетті функциялар бірігіп ортақтасады да лизогения жүреді. Фаг инфекциясы кезінде ген активтілігін қайта ауыстырып қосу транскрипция деңгейінде жүреді және промоторлармен РНҚ арасында арнайы арақатынас болады.
Эукариоттардағы гендер экспрессиясы реттелуінің ерекшеліктері
Жоғарғы сатыдағы эукариоттардың арнайы клеткаларының арасындағы өзгешелік ең алдымен белоктардың әртүрлі жиынтығын синтездеуіне байланысты. Осындай өзгешеліктерге жауапты ген экспрессиясы бақылануының жүйесі негізінен ДНҚ транскрипциясы деңгейінде әсер етеді.
Әрбір ген транскрипциясының реттелуі ерекше орналасқан элементтердің арнайы жиынтығының транскрипцияға тиісті факторлардың және транскрипциясының инициациясына әсер етуші белокты факторлардың болатындығымен анықталады. Транскрипцисы РНҚ-полимераза II-мен басталатын әдеттегі гендерде транскрипция инициациясы нүктесінен жоғары орналасқан промотор болады. Онда бірнеше қысқа келген (10 ж. н. ) нуклеотидтер тізбегі бар, олармен транскрипция факторлары байланысады. Бұл тізбектер ұзындығы 200 ж. н. артығырақ бөлімшеде бытырап жатады (13. 8-сурет) . Олар позитивті және негативті болуы мүмкін. Энхансерде транскипция факторларын байланыстыратын жақын орналасқан бірнеше элементтер болады. ДНҚ-ның реттеуші тізбектері транскрипцияның инициациясы әрі оның жылдамдығымен жиілігін реттеу үшін де қажет.

13. 8-сурет. Эукариоттар гендерінің реттеуші бөлігі (14) .
Сондықтан промоторлы аймақ РНҚ-полимераза II және транскрипцияның жалпы факторларын құрайтын бөлімшеден тұрады, сонымен қатар әртүрлі реттеуші белоктарды немесе транскрипцияның ерекше факторларын байланыстыратын тізбектердің көптеген реттеуші элементтері де болады (13. 9-сурет) . Промотор әдетте транскрипцияның басталу нүктесіне дейін 20-30 нуклеотидтерді қамтитын А-Т-ға бай келетін тізбектерден тұрады (5′-ТАТ-3′) . Промотордың бұл бөлімшесі Гольдберг-Хогнесс боксы деп аталады (прокориоттардығы Прибнов-боксқа ұқсас) немесе жай ғана ″ТАТА″ тізбегі делінеді. ТАТА-тізбегі транскрипцияның басталу нүктесін бақылауы мүмкін. Мұндай тізбектің болуы транскрипциялық бірліктерге тән болып келеді, оның оқылуына РНҚ-полимераза II қатысады. Содан соң CААТ (5′-GGCCAATCT-3′) және GC (5′-GGCCGG-3′) домендері келеді. Шамасы CААТ РНҚ-полимеразаның промотормен алғашқы байланысын бақылайтын болуы керек. GC доменінің көмегімен РНҚ-полимеразаның транскрипцияның басталу нүктесі аймағымен байланысуы жүзеге асады. Промоторларда әртүрлі элементтердің комбинациясы болуы мүмкін, бірақ элементтердің бірде-бірі барлық промоторларда кездесе бермейді.

Сіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz