Генетика: даму тарихы, зерттеу әдістері және тұқым қуалаушылықтың материалдық негіздері

Мазмұны
Кіріспе. Генетика ғылымының даму тарихы жəне зерттеу əдістері
1. Тұқым қуалаушылықтың материалдық негізі
1. 1Тұқым қуалаушылықтың цитологиялық негізі
1. 2Тұқымқуалау заңдылықтары. Г. Мендель ілімі
1. 3Аллелді емес гендердің өзара əрекеттесуі
2. Жыныс хромосомаларының жынысты анықтау механизмі
2. 1 Цитоплазмалық тұқымқуалау
3. Тұқымқуалаудың молекулалық негізі
3. 1 Геннің табиғаты
3. 2 Геннің құрылымы
3. 3 Генетикалық код
4. Өзгергіштік жəне оны жіктеу
4. 1 Мутациялар классификациясы
4. 2 Гендік мутация
4. 3 Хромосомалық мутация
4. 4 Геномдық мутация
4. 5 Цитоплазмалық мутация
5. Адам генетикасы. Адам генетикасын зерттеу əдістері
6. Популяциялық жəне селекциялық генетика негіздері
6. 1 Будандастыру түрлері
6. 2 Сұрыптау əдістері
8. Генетикалық терминдер сөздігі
Əдебиеттер
Кіріспе.
Генетика ғылымының даму тарихы жəне зерттеу əдістері
Генетика-организмдердің тұқым қуалаушылық жəне өзгергіштік қасиеттерін зерттеген ғылым. Тұқым қуалаушылық деп организмдердің өз белгілерін немесе қасиеттерін жəне даму ерекшеліктерін келесі ұрпаққа беру қабілетін айтады. Өзгергіштік дегеніміз организмдердің белгілерінде немесе қасиеттерінде айырмашылықтардың пайда болуы.
Осындай айырмашылықтарды белгілі бір түр ішіндегі өкілдер арасында немесе ата-аналық форма мен олардың ұрпақтары арасында байқауға болады. Тұқым қуалаушылық пен өзгергіштік организмнің қарама-қарсы қасиеттері екенін айта отырып оларды диалектикалық материализмнің даму заңын айқындайтын, бір-біріне байланысты категориялар ретінде қарау керек, өйткені органикалық дүние пайда болған жаңа өзгерістердің тұқым өуалауы арқылы дамиды. Организмнің негізгі 2 ерекшелігін тұқым қуалаушылықтың құрылымдық өлшемі - ген қамтамасыз етеді. Ал геннің материалдық негізі клетка ядросындағы хромосомаларда орналасқан ДНҚ молекуласымен анықталады. Ал оның белгілерді анықтауы жəне құрылысы генге яғни ДНҚ молекуласының тізбегіне байланысты.
Тұқым қуалаушылықтың материалдық негізі болып өзін-өзі өндіре алатын жəне бөліну процесіне жаңа клеткаларға тарала алатын қасиеттері бар клетканың барлық элементтері саналады. Осындай талаптарға жалғыз құрылым - хромосома ғана сай келеді, ол өзінің дəл көшірмесімен өндіріп клетканың бөліну процесінде заңдылықпен таралатын жəне өзінің гендері арқылы белгілердің дамуын анықтай алатын бірден-бір қосылыс. Сондықтан ДНҚ-сы бар клетканың кез-келген құрылымы тұқым қуалаушылық қасиетке ие бола алады. Зерттеулер ДНҚ-ның клетка ядросының хромосомасында ғана емес, сонымен бірге ол клетканың цитоплазмалық компоненттерінде де болып, бірқатар белгілердің тұқым қуалауына қатысатынын анықтады. Осыған байланысты тұқым қуалаушылық ядролық жəне цитоплазмалық болып екіге бөлінеді. Ген мен хромосоманың өзгеруі организмнің өзгергіштік қасиетінің материалдық негізін сипаттайды. Барлық белгілері бойынша бір-бірінен айнымайтын 2 дарақ мүмкін емес, өйткені олардың гендер құрамы ешқашан бірдей болған емес, бірдей болмайды да.
Тұқым қуалаушылық туралы алғашқы көзқарастар сол кезде басым болған идеялар мен философиялық жүйелерге тікелей байланысты болды. Грек мифологиясынан кейін ежелгі грек дəрігерлері тіршіліктің негізі, білгілердің тұқым қуалауы ұрықтың пайда болуы жəне ондағы əйел мен еркектің атқаратын рөлдері туралы байыппен толғады. Гиппократ былай деп айтқан болатын «Ұрықты организмнің барлық бөліктері бөліп шығарды, дұрыс ұрық дененің сау бөлігінде, бұзылған ұрық - ауру бөлігінде пайда болады. Егер тапалдан тапал, көк көздіден көк көзді, қысық көздіден қысық көзді туса, онда ұзын бастыдан ұзын басты тууына ештеңе бөгет бола алмайды». Гиппократ теориясы бойынша тұқым қуалаушылық информацияны таситын ұрық дененің барлық бөліктерінде пайда болады. Бұл қағида кейін пангенез теориясы деп аталды. Мұның мəні ұрық организмнің барлық дене бөліктерінде пайда болады да, қан тамырлары арқылы ұрық безіне келіп түседі. Ұрпақтың ата-анасымен ұқсастығын ұрықтың дененің əр түрлі бөліктерінің де пайда болып, солардың ерекшеліктерін өздеріне сақтауымен түсіндіруге болады. Тұқым қуалаушылықтың толық қалыптасқан теориясын идеалист философ Аристотель берді. Оның пікірінше организмнің дамуы тіршілік күші - энтелехияның бақылауында болады, ол организмде өтетін барлық материалдық процестерді анықтайды. Ол еркек жəне ұрғашы жыныстың ұрпаққа деген əсері бірдей емес деп есептеді. Оған алғашқы қозғалысты жіберетін - еркек жыныс, ал материалды беретін - ұрғашы болып көрінді. Егер еркек бастама күштірек болса ұл əкесіне көбірек ұқсайды жəне керісінше. Аристотелдің пікірінен қате теориялық елеске сүйенген бақылаудың теріс ұғым беретінін көреміз.
Аристотель ілімнің құндылығы ол пангенез теориясын жоққа шығарды, бұл ілім бойынша ұрпаққа дене бөліктерінің дайын үлгісі емес, эмбрионның дамуын бағыттайтын информация беріледі. Тұқым қуалаушылықтың мəні туралы Аристотель пайымдаған осындай терең ұғымға 23 ғасыр бойы мəн берілмеді. Сондықтан да тұқым қуалаушылық құбылысына материалистік тұрғыдан берілген түсініктемелер жəне кейін пайда болған көптеген идеялар пангенез теориясын анықтады. Демокриттің пікірі бойынша адам қабілетінің басым көпшілігі табиғи информация арқылы емес негізінен жаттығулар арқылы
дамиды. Демокрит бойынша еркек жəне ұрғашылардың ұрығында организмнің барлық органдарының материалдық бөліктері сақталды. Ол «адам бүкіл адамнан бөлінеді» деп айтқан болаты. Демокриттің тұқым қуалаушылыққа деген көзқарасында өте таңғырыттылық байқалса да, бірақ өзінің негізінде тұқым қуалаушылық құбылысын материалистік баяндауын пайымдады, еркек жəне ұрғашы жыныстың тұқым қуалаушылыққа қосатын үлесі тең деп саналды, өйткені олар қосылғаннан кейін жаңа ұрпақтың бастамасын беретін ұрпақ пайда болды. Платон, Аристотель жəне басқа да ежелгі грек философтары
Демокритпен қақпайласып, тұқым қуалаушылықтың материалдық сипатын үзілді-кесілді жоққа шығарды. Платон кез-келген организмнің қасиеттері мен ерекшеліктері ол туған кеде ғана оған қандай жанның орналасуына байланысты деп санады. Ол кезде Аристотельдің беделі орасан зор болатын, сондықтан да оның ойы əр уақытта дұрыс деп саналды. Тіпті қайта өрлеу дəуірі мен орта ғасырда Аристотельдің ойы бұрынғыдан да бағалы бола түсті.
Қайта өрлеу дəуірінде тұқым қуалаушылықтың ұғымына айтарлықтай жаңадан енбеді, бұл кезде жаратылыстану ғылымдары қайтадан жаңғырып догмалық наным жоққа шығарыла бастады. 17-18 ғасырларда преформациялық бағыт ерекше өріс алды. Оның теориялық негізі Гиппократ іліміне қарағанда əлдеқайда қарапайым болған еді, сондықтан ұзақ жылдар бойы əйгілі болып қала берді. Преформация теориясының негізін салушы Свамердам бойынша
титтей ұрық - гомонуклеус дайын күйінде жыныс клеткалардың ішінде орналасады екен. Дайын ұрықтың қай клеткада орналасқаны туралы Талас ұзаққа созылды. Дайын ұрық аналық жыныс клетканың ішінде болады дегенді жақтаушылардың тұжырымы бойынша сперматозоидтың əсері тек ұрықтың əрі қарай үлкейіп дамуына жағдай жасаумен ғана шектетеледі. Ал бұған қарсы ойды жақтаушылардың бірі Гартсекнер тіпті спертматозоидтың бас жағынан дайын ұрықты шамалап көруге болады деп абсурдты тезис айтқан. Мұндағы
аналық клетка ұрық көлемінің өсіп, ұлғаюына қажетті қоректік заттармен қамтамасыз етеді екен.
Орта ғасырдан кейін пангенез теориясы жаңа деңгейгде қайтадан кең өріс алды. Жан Батист те Ламарк панкезисті эволюциясының негізгі механизмі деп санады. Жеке мүшелерді жаттықтыру немесе сыртқы ортаның басқа факторларының əсерінен пайда болған өзгерістер Ламарк бойынша тұқым қуалай алады. Тұқым қуалаушылықтың жай-жапсарын ұғу үшін пангенез теориясын 19 ғасырда көптеген басқа да атақты ғалымдар тіпті Ч. Дарвин өзі қолдады. Ч. Дарвиннің уақытша пангенез гипотезасы бойынша ұрпақтың белгілері оның ата-анасының əр түрлі дəрежелерінен шығып жыныс клеткаларға келіп түсетін дискрет тұқым қуалайтын элемент - гемулла арқылы анықталады. 19 ғасырда тұқым қуалаушылықтың мəн-мағынасын сипаттайтын бірнеше ойша жорыту теориялар ұсынылды. Олардың ішінде генетика Авгус Вейсманның пангенез теориясына қарсы шығарған ұрық плазмасы теориясының маңызы өте зор, Вейсман жыныс клеткаларында тек ұрық плазмасынан ғана
пайда болады жəне ондағы тұқым қуалайтын факторлар уақытта өзгермейді жəне хромосомаларда орналасады деген дұрыс болжам жасады. Ал организмнің басқа бөліктеріннің клеткалары басқа бөліктерінің клеткалары сома плазмасына пайда болады олар өзгеруі мүмкін бірақ тұқым қуаламайды А. Вейсман көптеген ұрпақ көлемінде тышқандардың құйрығын кесіп олардан алынған ұрпақтардың ұзын құйрықты болатынын байқады. Демек тышқан құйрығының тұқым қуалау бастамасы сол құйрықтың ерекше бөліктерімен емес керісінше құйрық кесілген кезде өзгермейтін ұрық плазмасының клеткалары арқылы анықталады.
19 ғасырда əр түрлі ойша жорыту теорияларының авторлары арасында қызу таластар жүріп жатқанда ғылымда сол кездің өзінде-ақ мағынасы кемдеу сияқты болып көрінетін зерттеулер болған еді. Олар эксперимент жүзінде тұқым қуалау проблемаларының терең шынайы көріністерін ашты. Мұнда белгілердің тұқым қуалауын зерттеу өсімдік пен жануарлар гибридтерін алу арқылы жүргізілді.
Өсімдіктерді будандастыру арқылы гибридті зерттеу жұмыстарын алғаш рет Кельрейтер жүргізді. Ол əр түрге жататын өсімдіктерді будандастырудан алынған гибридтердің белгілері аралық формада яғни біркелкі болатынын байқады. Тозаң мен аналық белгілердің ұрпаққа тұқым қуалауына тең рөл атқарады деп дұрыс жорамал жасады. 1861 жылы Нодэн өз тəжірибелерінде бірінші ұрпақтың біркелкілігі ережесімен қатар тұқым қуалаушылықтың екінші ережесін белгілердің екінші ұрпақта ажырау сипатын ашты. Бірақ ол ережені дұрыс түсіндіре алмады Кельрейтер, Нодэн, Сажре жəне өсімдік пен жануалардың гибридтерін алумен айналысқан олардың басқа замандастары генетика ғылымының дамуына айтарлықтай үлес қосқаны мен олар тұқым қуалаушылықтың табиғи механизмін аша алмады, бір жағынан ол кезде бұл механизмнің цитологиялық негізі белгісіз еді, ең бастысы бұл ғалымдар барлық белгілердің тұқым қуалауын бір мезгілде зерттеуге тырысты. Тұқым қуалаушылықтың негізгі заңдылықтарын оның дискреттік табиғатын тұңғыш рет 1865 жылы Австрияның Брюн қаласының монахы Грегорь Мендель ашты.
Ас бұршақтың əр түрлі сорттарын шығылыстыру арқылы Г. Мендель белгілердің
Тұқым қуалаушылық факторларымен анықталатынының эксперименттік дəлелдемелерін ұсынды. Мендельдің табысқа жетуінің басты себебі - ол барлық белгілердің емес жеке альтернативті белгілердің тұқым қуалауын зерттеді, ұрпақтардың санын есептеп, тəжірибенің нəтижесін математикалық өңдеуден өткізді.
1. Бірінші ұрпақ белгілерінің біркелкілік ережесі.
2. Екінші ұрпақ белгілерінің 3\1 арақатынасына ажырау заңы.
3. Əр түрлі тұқым қуалау факторларының бір-біріне тəуелсіз тұқым қуалау заңы. Бұл заңдардың тұжырымдалуы биология ғылымының барлық саласының дамуына аса маңызды талпыныс əкелді. Алайда Мендель ашқан тұқым қуалау заңдылықтары оның замандастарына 35 жыл аралығында түсініксіз болды. Мендель заңдарын 1900ж Гуго де Фриз, Кар Коренс жəне Эрих Чермарк 2-ші рет ашты. Осы кезден бастап Мендель жұмыстарының тұқым қуалаушылық сырын ашудағы маңызы арта түсті. Генетикада əр салада жұмыс істейтін
биологтарды қызықтырып, жаңа ғылымның өркендеуінің ерекше талпынысы басталды. Сонымен генетика ғылым ретінде 1900 ж пайда болды, оның негізін 1865 ж Г. Мендель салды. Бұл екі арада генетика үшін маңызы зор клетканың бөліну заңдылықтары жəне оның материалдық заттары ашылды. 1874ж орыс ғалымы И. Д. Чистяков клетканың бөлінуін, 1882 ж Флеминг хромосомаларды ашты, Э. Ван-Бенеден 1883 ж аскариданың ұрықтану процесін зерттей отырып, жыныс клеткаларындағы хромосомалардың саны екі есе аз болатынын көрсетті, ал 1887 ж Т. Бовери ұрықтану кезеңінде хромосомалардың саны бұрынғы қалпына келетінін арнайы тəжірибелер арқылы дəлелдеді. Клетка ядросындағы хромосомалардың бөліну тəртібіне сүйене отырып 1887 ж А. Вейсман гендер хромосомаларда орналасады деген болжамды ұсынды. IX ғасырдың бас кезінде негізінен 3 ғалым Люсьен Кенс, Вильям Бэтсон жəне Вильем Кастл, Мендель принциптері тек өсімдіктерде ғана емес жануарларда да байқалатынын дəлелдеді. 1902ж Вальтер Сэттон жəне Теодор Бовери біріне-бірі тəуелсіз Мендель генетикасының митоз бен мейоз процестерімен тығыз байланысын атап айтқанда гендер мен хромосомалардың клетка бөлінуіндегі ажырауының ұқсастығын көрсетті. Гендер хромосомаларда шоғырланған деген болжам тек 1910ж эксперименттік дəлелге ие болды. Мұны атақты американ генетигі Томас Хант Морган жəне оның мектебі дрозофила шыбынына тəжірибелер жүргізіп анықтады. Сонымен қатар олар гендердің хромосома бойында тізбектеле орналасуын дəлелдеді. 1925-1943ж. ж аралығында гендер мен хромосомалардың сыртқы факторлардың əсерінен өзгеріске ұшырайтындығы дəлелденді. Сонымен Мендель заңдары қайтадан ашылған 1900 жылдан бастап 1953 жылға дейінгі кезең классикалық генетиканың дəуірі деп аталады. Осы мезгілде ген тек таза теориялық тұрғыдан ғана зерттелді.
Қазіргі заманғы яғни 1953 ж басталатын жаңа генетика дəуіріне тұқым қуалаушылық құбылысын молекулалық деңгейде зерттеу тəн. Жалпы тұқым қуалаушылық субстанциясының химиялық негізі нуклеин қышқылы ДНҚ-ға байланысты екенін 1944 ж О. Эвери, Мак Леод жəне Мак Карти көрсеткен болатын. 1953 ж ДНҚ молекуласы құрылымының ашылуы генетикалық зерттеулерді молекулалық деңгейде жүргізуге үлкен серпіліс берді. 50-60 ж. ж гендердің жұмыс істеу принциптері сондағы генетикалық кодтың концепциясы арнайы жүргізілген эксперименттер арқылы дəлелденді. Ал 70-жылдардан бері
генді химиялық жолмен синтездеу жолдары жəне генетикалық инженерияның теориялық жəне практикалық мəселелерін шешу мүмкіндіктерін зерттеу.
Генетиканы зерттеу əдістері.
Жоғарыда организмнің тұқым қуалаушылық жəне өзгергіштік қасиеттері молекулалық деңгейден популяциялық деңгейге шейін зерттелетіні атап өтілді. Зерттелетін белгілердің
тұқым қуалауы мен өзгеруін талдау үшін мал генетикасында мынадай əдістер қолданылады. Гибридологиялық талдауда организмнің белгілері мен қасиеттерінің тұқым қуалау заңдылықтарын зерттеу үшін оларды бір-бірімен
будандастырып, алынған бірінші, екінші жəне келесі ұрпақтарға талдау жүргізеді. Бұл əдісті чех ғалымы Г. Мендель қолданып, жетілтті.
Гибридологиялық əдіс генетикалық зерттеулерде негізгі əдіс болып саналады. Генеологиялық талдау бір-бірімен белгілі дəрежеде туыстық байлынысы бар мал тобына шежіре кесте құрастырып, онда белгінің тұқым қуалауын бірнеше ұрпақта бақылайды. Бұл əдіс ең алдымен адамның жəне малдың тұқым қуалаушылығын зерттеуде қолданылады.
Цитогенетикалық талдау хромосомалардың саны мен құрылысын жəне репликациясы мен қызметін зерттеу үшін қолданылады. Осы əдістің көмегімен хромосома құрлысының
жəне санының өзгеруі салдарынан малда əр түрлі генетикалық аурулар мен кемістіктер дамитыны анықталды. Популяциялық талдауда белгілердің тұқым қуалауын, өзгергіштік дəрежесін жəне олардың өзара байланысын анықтау мал тобында жүргізіледі. Əдіс негізінен математиканы қолдануды тірек етеді. Онтогенетикалық талдау организмнің жеке дамуында геннің əсерін зерттеу үшін қолданылады. Бұл əдістің көмегімен малды азықтандыру мен күту
жағдайларының өзгеруінің гендерге əсерін бақылауға болады. Кейінгі кезде малдың тұқым қуалаушылық қасиеттерін тереңірек зерттеу үшін жалпы генетиканың басқа да əдістері атап айтқанда биохимиялық, иммуногенетикалық, физиологиялық т. б. əдістері қолданыла бастады.
Тұқым қуалаушылықтың материалдық негіздері
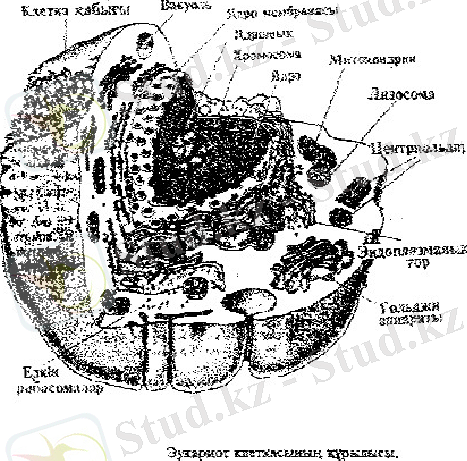
Клетка - тірі организмдердің негізгі құрылымдық бірлігі. Клетканың генетикалық информациясының материалдық негізін цитогенетика ғылымы зерттейді. Клетка күрделі биологиялық жүйе оның генетикалық құрылымы əр қилы жəне ол клетканың бас компоненттерімен тығыз байланысқан.
Организмнің кез-келген клеткасы 2 негізгі элементтен - ядро жəне цитоплазмадан тұрады.
Цитоплазманы құрамында органоидтар болады. Олар: митохондриялар, рибосомалар, лизосомалар, Голджи комплексі, эндомлазмалық тор жəне тек өсімдік клеткаларында ғана кездесетін пластидтер. Клетканың бұл құрылымдық элементтерінің əрқайсысының өзіне тəн құрылысы мен атқаратын қызметтері бар.
Митохондрия. Оның пішіні таяқша немесе түйіршік тəріздес, ұзындығы 5-7 мкм болып келеді. Оның іші-сыртын мембраналар қаптап жатады. Ішкі мембранасында криста деп аталатын қатпарлар болады. Əр организмдегі клеткалардың түріне қарай митохондриялардың саны шамамен 2-2, 5 мыңдай болады. Митохондриялардың негізгі қызметі клеткадағы заттардың алмасуы үшін қажетті энергенияның қорын жинақтайды. Ал
оның көзі - мейілінше энергияға бай қосылыс АТФ.
Рибосомалар негізінен эндоплазмалық тор мен ядро қабықшасының сыртқы қабатында орналасатын шағын денелер. Химиялық құрамы белок пен РНҚ -дан тұрады. Рибосомада ДНҚ мен РНҚ -ның қатысуында белок синтезі жүреді.
Лизосомалар - сыртқы липопртеидті мембраналармен қоршалған, диаметрі, 0, 4 мкм-дей бөлшектер. Олардың құрамында клетканың ішіндегі заттарды ыдыратуға қатысатын ферменттер жинақталады.
Гольджи комплексін ең алғаш рет 1898 жылы Италия оқымыстысы К. Гольджи тапқан, сондықтан ол соның есімімен аталады. Гольджи комплексі-мембраналар, гранулалар жəне вакуольдерден тұратын күрделі құрылым.
Онда заттар алмасуы процесінде бөлінетін жəне клеткадан сырқа шығарылуға тиісті ыдырау өнімдері - секреттер, кейбір улы заттар т. б. жинақталады.
Эндомлазмалы тор цитоплазманы торлап жататын əр түрлі ұзынды-қысқалы түтікшелерден тұрады. Ол клетканың ядросы мен бүкіл органоидтарын бір-бірімен байланыстырып тұратын заттардың алмасу процесіндегі бірден-бір реттеуші жүйе болып табылады.
Цитоплазмадағы аса маңызды органоид жасыл өсімдіктердің барлық клеткаларында кездесетін пластидтер. Ол үш түрлі болып келеді: фотосинтез процесіне қатысатын жасыл түсті түсті пигмент хлорофилл түзетін хлоропластар, түссіз - лейкопластар жəне əр түрлі пигменттер түзетін хромопластар.
Ядро - өзінің хромосомаларымен гендерінде организмнің негізгі генетикалық информациясын жинақтаған клетканың негізгі компоненті. Ядролы организмдер көп клеткалы балдырлар жануарлар мен өсімдіктер жəне бір клеткалы балдырлар мен қарапайымдар эукариоттар деп аталады. Ядросы қалыптаспаған организмдер немесе прокариоттар деп аталады. Ядро 2 түрлі күйде болуы мүмкін: тыныштық - интерфаза жəне бөліну стадиясы - митоз немесе мейоз.
Митоз. Бұл процесс кезінде тұқым қуалайтын материал - хромосомалар алдымен екі еселеніп алып, содан соң жана түзілген екі клеткаға тең мөлшерде бөлінедіМитоздың генетикалық мəнінің өзі бір организмге тəн тұқым қуалайтын информацияның жаңа түзілген екі клеткада бірдей, ұқсас болатындығында. Клетка өзінің бөлінуі барысында бірнеше кезеңнен өтеді, оларды біріктіріп клеткалық немесе метоздық цикл деп атайды. Клеткалық
циклдің өзі бірнеше фазалардан тұрады. Олар: интерфаза, профаза, прометафаза, метафаза, анафаза, жəне телофаза.
Интерфазалық клетка ядросында ядро қабығы, ядро шырыны, ядрошық жəне хромосомалар болады. Ядро қабығы ядроны цитоплазмадан бөліп тұрады, ядро қабығында көптеген ұсақ саңылаулар болады, олар арқылы ядро цитопалазма арасында байланыс қамтамасыз етеді. Ядро шырыны қоймалжың зат, онда
ядрошықтар мен хромосомалар болады. Ядрошықтың құрамында рибосома түзілуі үшін аса қажетті рибонуклеин қышқылы болады жəне олар хромосомалармен белгілі бір байланыс құрайтын денешік болып табылады. Хромосома тұқым қуалаушылық информацияға жауапты жалпы клетка жүйесінің негізгі жəне басты органоиды. Хромосомалардың морфологиялық құрылысы митоздың метафаза стадиясында жақсы байқалады. Осы кезеңде хромосома біршама тығыз консистенциялы негізгі бояғыштармен айқын боялатын қос таяқша тəрізді болып көрінеді. Хромосоманың формасы алғашқы немесе центромералық тартылысқа байланысты. Алғашқы тартылыстың белгілі бір бөлігінде клетка бөліну кезеңдерінде хромосома қозғалысын меңгеретін арнайы зат - центромера орналасады. Центромералық тартылыс хромосоманың денесін 2 иыққа бөледі. Центромерасының орналасуы əр түрлі хромосомалар үшін тұрақты жəне соған байланысты хромосоманың 3 морфологиялық типін ажыратады.
Метацентрлі хромосома центромера оның орта бөлігінде орналасқан яғни мұнда хромосоманың иықтарының ұзындығы бірдей немесе шамалас болып келеді. Субметацентрлі хромосома иықтарының ұзындығы əр түрлі хромосома.
Сіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz