Фотосинтездің С3 және С4 жолдары: молекулалық механизмдер мен қоршаған ортаның әсері

Аннотация
Курстық жұмыстың тақырыбы «Фотосинтездің С 3 және С 4 жолдары». Курстық жұмыс 34 беттен тұрады. Курстық жұмыста 10 сурет бар.
«Кіріспе» бөлімінде фотоситнзе процесінің пайда болуы және оның жүру жолдары туралы қысқаша қарастырылған.
«Фотоситнзе туралы ұғым» бөлімінде автотрофты қоректену түрлері, өсімдіктердің фотосинтез аппараты, хлорофил, хлорофилдің фотосинтезге процесіне қатысуы, каротиноидтар, пигменттер жүйесіндегі энергия миграциясы, пигменттер фотожүйесі, су фотолизі туралы толық қарастырылған.
«Фотосинтездің С 3 және С 4 жолдары» бөлімінде фотосинтездік фосфорлаудың циклды және циклсыз түрлері, фосфорлаудың хемиосмостық гипотезасы, фотосинтездің биохимиялық процестері, яғни фотосинтездің С 3 және С 4 жолдары, фотосинтез процесіне сыртқы жағдайлардың әсері келтірілген.
«Қорытынды» бөлімінде фотосинтездің жүру жолдары, оның ішінде С 3 және С 4 жолдары туралы қарастырылғаны туралы жазылған.
Жоспар
Кіріспе . . . 3
І Фотосинтез туралы ұғым
1. 1 Автотрофты қоректену түрлері . . . 4
1. 2 Өсімдіктердің фотосинтез аппараты . . . 5
1. 3 Хлорофил . . . 7
1. 3. 1 Хлорофилдің фотосинтезге процесіне қатысуы . . . 10
1. 4 Каротиноидтар . . . 12
1. 5 Пигменттер жүйесіндегі энергия миграциясы . . . 13
1. 5. 1 Пигменттер фотожүйесі. Реакция орталықтары . . . 13
1. 6 Су фотолизі . . . 14
ІІ Фотосинтездің С 3 және С 4 жолдары
2. 1 Фотосинтездік циклді және циклсіз фосфорлау . . . 16
2. 1. 1 Фосфорлаудың хемиосмостық гипотезасы . . . 18
2. 2 Фотосинтездің биохимиялық процестері . . . 19
2. 2. 1 Фотосинтездің С 3 жолы . . . 20
2. 2. 2 Фотосинтездің С 4 жолы . . . 22
2. 3 Фотосинтездік тыныс алу . . . 25
2. 4 Фотосинтез процесіне сыртқы жағдайлардың әсері . . . 27
2. 5 Фотосинтез және түсім . . . 32
Қорытынды . . . 33
Пайдаланылған әдебиеттер . . . 34
Кіріспе
Курстық жұмыстың көкейкестілігі. Фото . . . және синтез - жоғары сатыдағы жасыл өсімдіктердің, балдырлардың, фотосинтездеуші хлорофилл және басқа дафотосинтездік пигменттер арқылы күн сәулесі энергиясын сіңіруі нәтижесінде қарапайым қосылыстардан (көмірқышқыл газы, су) өздерінің және басқа организмдердің тіршілігіне қажетті күрделі органикалық заттар түзуі. Фотосинтез нәтижесінде жер жүзіндегі өсімдіктер жыл сайын 100 млрд т -дан астам органикалық заттар түзеді (мұның жартысынан көбін теңіз, мұхит өсімдіктері түзеді) және бұл кезде олар 200 млрд-тай СО 2 сіңіреді, оттегін бөледі.
Фотосинтез аппаратының негізі - клетка ішіндегі органелла-хлоропластар (көк жапырақ клеткасында 20-100 болады) .
Балдырлардың көпшілігінде фотосинтездік аппарат - клетка ішіндегі арнайы органелла-хроматофорлар, ал фотосинтездеуші бактериялар мен көк-жасыл балдырларда тилакоидтер. өсімдік фотосинтез процесінің негізі - тотығу-тотықсыздану. Мұнда квант энергиясы әсерінен 4 электрон мен протон су дәрежесінен (оның тотығуы) углевод дәрежесіне дейін көтеріледі. (СО
2
-ның тотықсыздануы) . Сөйтіп углеводтар фотосинтезі былай өтеді:
 СО
2
+Н
2
О
СО
2
+Н
2
О
 С(Н
2
О) +О
2
+120
ккал/моль
яғни СО
2
-ның бір молекуласының углевод дәрежесіне дейін тотықсыздануының бос энергиясы 120
ккал/моль
болады. Демек, өсімдік фотосинтезі кезінде кем дегенде 3 квант («қызыл» кванттар энергиясы 40
ккал/моль
) сіңірілуі қажет. әр түрлі жағдайда жасалған тәжірибе СО
2
-ның әр молекуласының тотықсыздануына 8-10 квант қажет екенін көрсетті. Көмірқышқыл газ да, су да, жарықты тікелей сіңірмейді, бұл қосылыстардың квантпен байланысқа түсуін хлоропласт не хроматофор структурасындағы хлорофилл
а
қамтамасыз етеді. Фотосинтездің биосферадағы маңызы да үлкен.
С(Н
2
О) +О
2
+120
ккал/моль
яғни СО
2
-ның бір молекуласының углевод дәрежесіне дейін тотықсыздануының бос энергиясы 120
ккал/моль
болады. Демек, өсімдік фотосинтезі кезінде кем дегенде 3 квант («қызыл» кванттар энергиясы 40
ккал/моль
) сіңірілуі қажет. әр түрлі жағдайда жасалған тәжірибе СО
2
-ның әр молекуласының тотықсыздануына 8-10 квант қажет екенін көрсетті. Көмірқышқыл газ да, су да, жарықты тікелей сіңірмейді, бұл қосылыстардың квантпен байланысқа түсуін хлоропласт не хроматофор структурасындағы хлорофилл
а
қамтамасыз етеді. Фотосинтездің биосферадағы маңызы да үлкен.
Курстық жұмыстың мақсаты: фотосинтез процесін бақылап, оның жүру жолдарын, әсіресе ішіндегі С 3 және С 4 жолдарын толық зерттеп, баяндау.
Курстық жұмыстың міндеті - фотосинтездің барлық жүру жолдарын анықтап, негізгілерінің реакцияларымен танысу.
І Фотосинтез туралы ұғым
1. 1 Автотрофты қоректену түрлері
Афтотрофты қоректенудің жоғары және төменгі сатыдағы жасыл өсімдіктер фотосинтезі, бактериялық фотосинтез немесе фоторедукция және хемосинтез түрлері болады. Біздің планетамыздағы органикалық заттардың барлық қоры осы үш процестің нәтижесінде түзілген. Жасыл пигменті бар жоғары және төменгі сатыдағы өсімдіктерде фотосинтез процесі жүреді. Хлорофилдің қатысуымен өсімдіктер күн энергиясына айналдырады. Хлорофилдің сіңірген энергиясы бірқатар аралық акцепторлар арқылы су мен көмірқышқыл газына жеткізіледі, осының нәтижесінде инертті болып саналатын бұл заттар күрделі органикалық қосылыстар түзе отырып әрекеттеседі. Жалпы алғанда, фотосинтез реакциясының жиынтық теңдеуін Ван-Ниль бойынша былайша жазуға болады:
6СО 2 + 12Н 2 О → 6О 2 + С 6 Н 12 О 6 + 6Н 2
Бұл теңдеу СО 2 және Н 2 О сияқты жай минералдық заттардан күн сәулесінің энергиясының есебінен органикалық зат - көмірсу түзілетінін көрсетеді. Фотосинтез кезінде бос оттегі бөлініп шығуының зор маңызы бар. Сол процестің арқасында тіршіліктің аэробты түрлері дамыды.
Соңғы кездегі зерттеулер көрсеткеніндей, фотосинтез процесінде күн сәулесінің энергиясы NADPH және АТР 4- түзілуіне жұмсалады. Сондықтан фотосинтез теңдеуін М. Кальвин былайша келтіреді:
6СО 2 + 12NADPH + 18АТР 4- + 12Н 2 О → глюкоза + 12NADP + + 18ADP 3- + 18Н 3 РО 4 + 6Н +
∆G (рh=7) = -378 кДж/моль -1 .
Эволюция барысында фотосинтез процесін жүргізуші организмдер хемосинтез процесін жүргізуші организмдер кейін пайда болды, алайда фотосинтез процесінде қоры іс жүзінде шексіз болып табылатын заттар мен күн энергиясы пайдалатындықтан, бұл процесс барынша дамыды. Жоғары сатыдағы өсімдіктер фотосинтезінің арқасында көмірдің, шымтезек пен мұнайдың қоры ерте замандағы геологиялық дәуірлерде қалыптасты. Жер бетінде тіршілік ететін гетеротрофтардың бәрі де қоректік заттарды өсімдіктер фотосинтезі есебінен қабылдайды. [2]
Бактериялық фотосинтез эволюция тұрғысынан алып қарағанда өте ертеде пайда болған. Бактериялық фотосинтез немесе фоторедукция деп аталатын бұл процесс мынадай бактериялар тобында жүзеге асырылады: пурпурлық (қанқызыл) және қызыл күкірт бактериялары, пурпурлық күкірт емес бактериялар, жасыл күкірт бактериялар жатады. Олардың құрамында хлорофиллден біраз айырмашылығы бар бактериохлорофилл пигменті және картиноидтар бар. Пигменттері болуының арқасында олар күн сәулесінің энергиясын сіңіреді. Бактериялық фотосинтез бен өсімдіктер фотосинтезінің ұқсастығы, міне, осында. Мұнда сутегі көзі ретінде су емес, көмірсутектер, күкіртсутектер мен май қышқылдары пайдаланылады. Сутегі көздері аз таралғандықтан, сондай-ақ олардың қоры өте шектеулі болғандықтан, фоторедукция процесі кең дамып, таралмаған.
Атмосферада бос оттегінің жиналуы салдарынан автотрофтық қоректенудің үшінші түрі - хемосинтез жүзеге асады. Хемосинтез жарық жоқ жердегі аэробты жағдайларда тіршілік ететін, пигментсіз бактериялар тобына тән. Көмірқышқыл газын тотықсыздандыру үшін бұл бактериялар күн энергиясын емес, химиялық тотығу реакцияларының энергиясын пайдаланылады. Күкірттің алуан түрлі қосылыстарын тотықтыратын күкірт бактерияларының үлкен тобы және нитрификациялаушы бактериялар хемосинтез бактерияларына жатады. Күкірттің тотығуы нәтижесінде энергия бөлініп шығады, оны бактериялар органикалық қосылыстар түзе отырып, көмірқышқыл газын тотықсыздандыру үшін пайдаланады. Хемосинтездің фотосинтезден энергия көзі жөнінен де, сондай-ақ сутегі көзі жөнінен де айырмашылығы бар. Бұл процесс оттегін бөліп шығармастан өтеді. Энергия мен сутегі көздерінің шектеулі болуы салдарынан хемосинтез кең таралмаған.
Жалпы алғанда, Жер бетіндегі өсімдіктер атмосферадан 170 млрд. т. СО 2 сіңіреді, 130 млрд. т. Су жұмсап, 2•10 21 Дж Күн энергиясы есебінен 100 млрд. т. органикалық зат түзеді.
1. 2 Өсімдіктердің фотосинтез аппараты
Фотосинтез процесі негізінен өсімдіктердің жасыл жапырақтарында өтеді. Жапырақ тақтасының анатомиялық құрылысынан палисадты және борпылдақ паренхима байқалады. Паренхима жасушаларында ерекше органоидтар - хлоропластар болады, онда фотосинтездің күрделі реакциялары өтеді.
Барлық жоғары сатыдағы өсімдіктер хлоропластарының пішіні мен шамасы белгілі дәрежеде бірдей болып келеді. Өсімдіктердің түрлеріне қарай олардың бір жасушадағы саны түрліше болады. Габерландтың мәліметі бойынша үрмебұршақ жапырағының 1 мм 2 ауданында 283 000, күнбағыс жапырағының 1 мм 2 ауданында 465 000 хлоропласт кездеседі. Mnium мүгінің бір жасушасында шамамен 106 хлоропласт болады. Балдырларда кездесетін хлоропластардың шамасы мен пішіні алуан түрлі. Хлореллада табақша пішінді бір ғана хлоропласт болады. Спирогира балдырындағы хлоропласт спираль тәрізді оралып жатады. Көк-жасыл балдырларда айқын білінетін хлоропласт жоқ, бірақ жасуша қабырғасында ламеллалар жақын орналасқан. Ламеллаларға бүкіл хлорофил шоғырланған, сондықтан цитоплазманың бұл учаскесі хлоропластың қызметін атқарады. [4]
Хлоропластың бетіндегі қос мембрана, ішіндегі құрылымсыз сұйық матрикс, әр жерінде граналар деп аталатын күрделенген қабатты түзінділері бар жұқа ламеллалар (оның да мембраналық құрылысы болады) хлоропластар құрылымының негізгі элементтері болып табылады. ХІХ ғасырдың соңында ғана Мейер мен Шимпер хлоропластарда граналардың болатынына алғаш рет назар аударды бұдан кейін Хейтц граналарды бірінің үстіне бірі жиналған теңгелер сияқты орналасқан жазық дискілер ретінде сипаттады. Фрей-Висслинг грананың әрбір жеке дискісі шеттері қосылған және қабысып тұйықталған қапшықболып табылатын екі ламелладан тұратынын көрсетті. қабысып тұрған мұндай қапшықтарды тиллакоидтар деп атайды. Грананың әрбі тиллакоиды көршілес грананың тиллакоидтарымен жұқа екі ламелла арқылы байланысқан. Хлорофил мен картоноидтар граналардың тиллакоидтарына шоғырланған. Граналары жоқ хлоропластарда хлорофилл мен сары пигменттер молекулаларының құрылысы мен қасиеттері ерекше. Олар граналардағы ақуыздар және липидтермен бірге құрылысы қатаң белгіленген жүйе құрады.
 1 - сурет. Өсімдік жапырағынан алынған хлороплаттың электрондық микрофотосуреті.
1 - сурет. Өсімдік жапырағынан алынған хлороплаттың электрондық микрофотосуреті.
Жапырақ жасушаларынан ерекше әдіспен бөлініп алынған хлоропластар фотосинтез реакциясының бүкіл комплексін жүзеге асыра алады. Тіпті грананың жеке бөліктері де фотосинтез реакциясының бір бөлігін, атап айтқанда, Хилл реакциясы деп аталатын жарықты сіңіру және суды айыру реакциясын, ал оларға қажетті ферменттерді қосқанда көмірқышқыл газын сіңіру реакциясын да жүзеге асыра алады. Хлоропластың күрделі құрылымы фотосинтез реакциясының күрделілігіне сай келеді. Құрылымының сипатталған ерекшеліктерінің арқасында фотосинтездің алуан түрлі реакциялары кеңістікте бөлінеді де, олар бір мезгілде, тез және тиімді түрде өте алады. Фотосинтездің көмірқышқыл газын сіңіру және органикалық заттарды синтездеу жөніндегі басқа да реакциялары хлоропластардың сұйық ортасында өтеді. [10]
Толық жетілген хлоропластардың құрылысы мембраналы болады, ал пропластидтерде мембраналы құрылым болмайды. Өсімдікке жарық қалыпты түсіп тұрған жағдайда ламеллалар үнемі жетіле береді, олардың қалыптасуы үшін хлорофиллдің болуы қажет. Бұл хлорофиллдің өзі ламеллалар үшін құрылыс материалы болып табылатынына байланысты. Өсімдіктерді қараңғыда өсіргенде, олардың жапырағында хлорофилл түзілмейді. Мұндай өсімдіктерді этиолданған өсімдіктер деп атайды. Этиолданған өсімдіктерде хлорофилл болмағандықтан, олардың пластидтері ламеллалар түзбейді. Мұндай өсімдіктерді жарыққа шығарып қойғанда, оларда тез арада хлорофилл синтезделіп, одан соң хлоропластың ламеллалық құрылымы жетіледі. Демек, ламеллалар мен граналардың түзілуі хлорофиллдің болуына байланысты. Хлорофиллдің синтезделуі жарық түсу жағдайларына байланысты. Су өсімдігі элодеяны қыста шашыраңқы жарықта өсіргенде хлоропластағы ламеллалар айқын білінбейді. Элодеяны қызыл жарықта өсіргенде айқын білінетін жұқа ламеллалар түзіледі, онда сондай-ақ граналар да болады. Көк жарықта өсіргенде элодея хлоропласты ісінеді. Жазда элодеяның граналы мен граналар арасындағы ламеллалары жақсы жіктеледі. Борпылдақ және палисадты паренхима жасушаларының айырмашылығы бар. Борпылдақ паренхимамен салыстырғанда палисадты жасушаларда хлоропластардың саны көбірек, фотосинтез қарқыны жылдамырақ, а хлорофилінің мөлшері көбірек болады. Мұнда крахмал, ал борпылдақ паренхима жасушаларында қант түзіледі. Борпылдақ жасушалар түзілген қантты флоэмаға жеткізуге арналған. Палисадты жасушалар фотосинтезді күшті жарық жағдайында, ал борпылдақ жасушалар - шашыраңқы жарықта жүргізуге бейімделген.
1. 3 Хлорофил
Күн спектірінің көрінетін бөлігінде жарық сәулелерін талдап сіңіретін заттар - пигменттер деп аталады. Барлық органикалық пигменттер молекулаларының атомдары арасында бір және қос байланыс кезектесіп келіп отыратын жүйелері болады. Мұндай байланыс жүйесі коньюгацияланған жүйе деп аталады. 1906 жылы М. С. Цвет өсімдіктердің көпшілігінде хлорофиллдің а және в деген екі түрі болатынын көрсетті, ал қазіргі кезде хлорофиллдің 10- ға жуық түрі белгілі, а хлорофилі фотосинтез процесін жүргізуші барлық өсімдіктерде болады. Жоғары сатыдағы өсімдіктер мен жасыл балдырларда а хлорофилімен қатар в хлорофилі де болады. Қоңыр және диатомды балдырларда - с хлорофилі, қызыл балдырларда - d хлорофилі болады. Пурпурлық күкірт бактерияларында бактериохлорофилл кездеседі, а хлорофиллдің формуласы: - С 55 Н 72 О 5 N 4 Мg. в хлорофиллдің эмпирикалық формуласы - С 55 Н 70 О 6 N 4 Мg. ал бактериохлорофиллдің формуласы - С 55 Н 74 О 6 N 4 Мg. хлорофилл құрамында фитол С 20 Н 39 Он және метанол СН 3 ОН спирті бар хлорофиллин қышқылының күрделі эфирі. Спирт тобының қалдықтары хлорофиллин қышқылының карбоксил тобындағы сутегінің орнын басады. Эфир ретінде хлорофилл сабындалу реакциясына қабілетті келеді. Қышқылдың әсер етуімен хлорофилл молекуласындағы магний өзінің орнын сутегінің екі атомына беріп, феофитин түзеді.
Жарық сәулесінің энергиясын сіңіру және оны басқа қосылыстарға беру немее флуоресценция кванттары түрінде қайтадан шағылыстыру қабілеті хлорофиллдің аса маңызды қасиеттері. Бұл қасиеттер тұтасымен алғанда атомдардың ерекшеліктеріне және молекуласының құрылымын неміс биохимиктері Вильштеттер мен Штоль анықтаған. Хлорофилл молекуласының орталық бөлігі төрт пиррол тобынан түзілген. Бұл топтар порфирин сақинасын түзеді.
Порфирин сақинасының ортасына магний атомы орналасады. Пиррол тобы рим цифрларымен (І, ІІ, ІІІ, ІV) нөмірленеді. Хлорофилл молекуласында пиррол тобына қосымша бесінші изоциклді циклопентан тобы болады, ол V цифрымен белгіленеді. Порфирин сақинасының сыртқы атомдары сағат тілі жүрісінің бағыты бойынша араб . Фитол қалдығының көміртектік ұзын тізбегі молекуланың порфирин бөлігіне қосылады. Қазіргі кезде хлорофилл молекуласының мұндай құрылысы анықталған деп есептеуге болады, өйткені ол а және в хлорофилін қолдан синтездеу арқылы дәлелденіп отыр. Неміс биохимигі Штрель мен американ биохимигі Вудворд 1960 жылы а хлорофилін қолдан синтездеу әдісін жүзеге асырады. 2-суретте а хлорофилі молекуласының құрылысы көрсетілген. Бұл хлорофиллден в хлорофилінің айырмашылығы мынада: в хлорофиліндегі көміртегінің үшінші атомына СН 3 тобының орнына - СОН тобы қосылған. Порфирин сақинасында тұйық цикл түзетін коньюгация байланыс жүйесі болуының маңызы зор. мұндай жүйеде π-электрондар еркін айналым жасайды. [5]
Азоттың әрбір атомындағы электрондардың үш жұбы көміртегінің көршілес орналасқан атомдарымен ортақтастырылған. Азот электрондарының тағы бір жұбы магний атомына қарай бағытталған орбитальда орналасқан. Тоғызыншы көміртегінің атомына қосылған оттегі атомының ортақтаспаған бұл электрондары мен азоттың магний атомына қарай бағытталған орбиталь бойындағы электрон жұбы π-электрондар деп аталады; олар жарықты сіңіріп, жарық кванттарының әсерімен орын ауыстырады. Азоттың магний атомына қарай бағытталған π-электрондардың арқасында хлорофилл қызыл сәулелерінің кванттарын сіңіреді. Хлорофилл сондай-ақ оттегі атомының ортақтаспаған 4 электрондарының арқасында күлгін-көк сәулелерді сіңіреді. Коньюгация байланыстарының тұйық циклінің болуы нәтижесінде хлорофилл сіңірген жарық толығымен бере алады. Осының арқасында хлорофиллдің көптеген жекелеген молекулалары сіңірген кванттар энергиясы бір орталыққа жиналады. Ал а және в хлорофилі сіңіру спектрлерінің біраз айырмашылықтары бар, бұл олардың құрылымындағы аздаған айырмашылықтарға байланысты. 660 нм аймағындағы қызыл сәулелер мен 429 нм аймағындағы көк сәулелерді а хлорофилі барынша сіңіреді. 643 нм және 453 нм аймақтағы қызыл және көк сәулелерді в хлорофилі сіңіреді. Бұл екі хлорофиллдің максимум сіңіру спектрлерінің өзара сәйкес келмеуі сіңіру сәулелерінің барынша кең диапазонын қамтуға мүмкіндік береді. а хлорофилінің ерітіндісі көк-жасыл түсті, ал в хлорофилінің ерітіндісі жасыл-сары түсті болады. Егер екі ерітіндіні де оған түскен жарық жағынан қарайтын болсақ, онда ашық-жасыл түсті жарқылды көреміз, бұл - флуоресценциялық сәуле шығару.
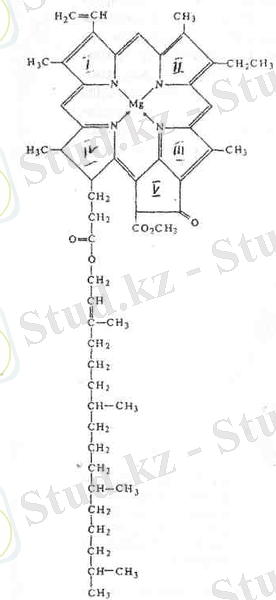
2 - сурет. Хлорофилл а молекуласының құрылымы.
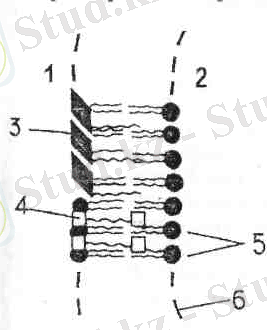
3 - сурет. Тиллакоид мембранасында пигменттердің орналасу схемасы.
Өсімдіктердегі хлорофилл күйі. Хлорофиллде порфирин ядросының гидрофильдік қасиеттері, ал молекуланың фитолдық ұшының липофильдік қасиеттері бар. Хлорофилл молекулалары ламеллалардың бетіне қалыңдығы бір молекуладай болатын тығыз қабатпен орналасады деп есептеледі. Липофильдік қасиеттері бар фитол липидтермен қосылыс түзуде белгілі роль атқарады. Хлорофилл молекуласының порфиринді бөлігі, керісінше, липидтерден қашықтайды, бірақ ақуыздарға жақындай алады. Сөйтіп, хлорофилл молекулалары арнаулы ақуыздармен байланысады. Осындай мәліметтердің негізінде хлорофиллдің фитолдық ұшы ламеллалардың липидтік қабатына батырылып тұратындай болып орналасқан деп жорамалдайды. Сонымен, ламеллалар мен граналардағы хлорофилл липидтермен және ақуыздармен байланысты болады. Бұл байланыс өте берік немесе нашар болуы мүмкін, ол хлоропластағы хлорофиллдің күйін күрделендіре түседі. Бір жасушалыбалдырлар мен жоғары сатыдағы өсімдіктердің а хлорофилі қызыл бөлікте 662, 670, 677, 682 нм-ге дейінгі спектрлерді сіңіреді. Бұл жағдай хлорофиллдің әртүрлі күйде болатынын білдіреді. Хлорофиллдің ақуыздармен және липидтермен байланысы ғана әр түрлі болмастан, сондай-ақ хлорофилл молекулаларының өзара қосылу дәрежесі де түрліше болуы мүмкін. Хлорофилл молекулалары өзара байланыспай тұрған күйін мономер түрі деп атайды. Ал өзара әрекеттесіп, байланысқан түрін агрегатты түрі деп есептейді. Оны Д. И. Сапожников, А. А. Красновский және т. б. атап көрсеткен. Хлорофиллдің бұл түрлерінің сіңіру спектрі өзгеше болады. Жасыл эвгленада а хлорофилінің С 695 белгісімен таңбаланған, ал жоғары сатыдағы өсімдіктерде - Р 700 белгісімен таңбаланған ерекше түрі бар екені анықталған. Бұл - бірінші хлорофиллдің 695 нм аймақта, ал екіншісінің 700 нм аймақта жарық сәулесін сіңіретінін білдіреді, а хлорофилінің бұл түрлері де олардың ерекше күйіне байланысты.
1. 3. 1 Хлорофилдің фотосинтезге процесіне қатысуы
Хлорофилл жеке алғанда активтігі шамалы қосылыс. Алайда оған жарық сәулесі түссе болғаны, хлорофилл солсәтте, 10
-15
с ішінде жарық энергиясын сіңіріп, қозған күйге келеді. Бұл - фотофизикалық кезең. Қозған күйдегі хлорофиллдің қозбаған күйдегі хлорофиллден айырмашылығы бар. Қозған күйдегі хлорофиллдің түсі өзгеріп, ол жасыл түстен қызыл-қоңыр түске енеді. Хлорофиллдің қозбаған күйі оның негізгі күйі деп аталады. Бұл кездегі электрондардың энергиясы нольге тең. Хлорофилл молекуласының жарық квантын сіңіруі және қозуы оның электрондарының негізгі күйден синглеттік қозу күйіне өтуіне себепші болады. Қызыл сәулелерді сіңірген кезде электрондар бірінші синглеттік қозу деңгейіне (S
 ), көк сәулелерді сіңірген кезде екінші синглеттік қозу деңгейіне (S
), көк сәулелерді сіңірген кезде екінші синглеттік қозу деңгейіне (S
 ) өтеді. Қозған молекула қозбаған молекуладан өзгеше, жұлдызшамен белгіленеді:
) өтеді. Қозған молекула қозбаған молекуладан өзгеше, жұлдызшамен белгіленеді:
хл + hυ → хл*
жарық кванты немесе фотон, жарық энергиясының дискреттік (бөлінбейтін) үлесі болып табылады. Квант энергиясының мөлшері мынаған тең: Е = hυ. Мұндағы h - Планк тұрақтысы, ал υ - толқын ұзындығына кері пропорционалды жарықтың жиілігі. Көк сәулелердің квант энергиясы 65 ккал/эйнштейнге, қызыл сәулелердің энергиясы 41 ккал/эйнштейнге тең. Хлорофиллдің жарық квантын сіңіру кезінде оның электрондарының энергиясы квант энергиясына тең болатын шамаға артады.
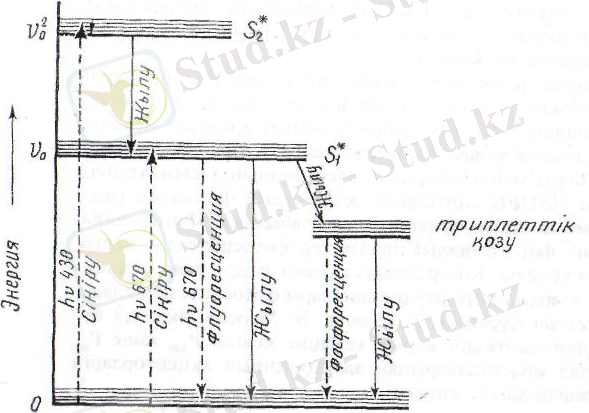
4 - сурет. Хлорофилл молекуласының энергияны сіңіріп, синглеттік S
 , S
, S
 және трилеттік қозу деңгейіне өту, флуоресцения және жылу бөліп шығару схемасы.
және трилеттік қозу деңгейіне өту, флуоресцения және жылу бөліп шығару схемасы.
Сіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz