Ұлпалардың пайда болуы және эволюциялық заңдылықтары

Марат Оспанов атындағы Батыс Қазақстан Мемлекеттік Медицина Университеті
Білім алушының өзіндік жұмысы
Факультет:Жалпы медицина
Кафедра:Гистология кафедрасы
Дисциплина:Гистология - 1
Курс: 2
Тақырыбы:Тканьдердің пайда болуы және эволюциялық заңдылықтары
Орындаған:Тәжіғос А. Т.
Тобы:207 Б
Тексерген:Бердалинова А. К.
Бағасы:
Оқытушықолы:
Күні:_ _
Ақтөбе 2016ж
Жоспар:
- Тканьдер(тін) - туралы түсінік
- Тканьдердің эволюциялық пайда болуы, болжамдары, Зарварзин бойыша тіндердің пайда болуы
- Тканьдер жіктелісі
- Тканьдік гомеостаз, эпидермистің тұрақтылығын қамтамасыз ететін факторлар
Тканьдер туралы түсінік
Тканьдер- арнайы мамандарлған, топтасқан және атқаратын қызметі бір-бірімен байланысты жасушалар жүйесі. Тканьдерді атқаратын қызметіне, шыққан тегіне, морфологиясына, т. б. қасиеттеріне байланысты бірнеше топқа бөледі. Егер тканьдер біркелкі жасушалардан құралған болса, оны жай ( колленхима), ал бірнеше жасушалардан тұрса - күрделі ( эпидерма) деп жіктейді.
Тканьдердің эволюциялық пайда болуы, болжамдары, Зарварзин бойыша тіндердің пайда болуы
Тканьдерде клеткалардың пайда болуы, мамандануы және бірігуі тарихи категория болып табылады, өйткені олар филогенез барысында пайда болды. Бірақ бұл категорияларды түсіндіру өте қиын, өйткені прокариоттық және эукариоттық организмдердің клеткаларының арасында маңызды айырмашылықтар болады. Соған қарамастан бірнеше болжамдар белгілі. Бактериялар мен цианобактериялардың қазба қалдықтарын зерттеу негізінде клеткалық форманың арғы тегі осыдан 3, 5 х 10 9 жыл бұрын пайда болған қарапайым прокариоттық клетка болуы мүмкін деп болжайды. Бұл типтің клеткалары өздерінің тіршілігін және көбеюін қамтамасыз ету үшін алғашқыда табиғаты биологиялық емес органикалық молекулаларды пайдаланды. Қарапайым клеткалардың қалыптасуына жасалған алғашқы қадам клетканың затын қоршайтын мембрананың пайда болуы еді.
Содан кейін қарапайым прокариоттық клеткаларда синтез бен энергетикалық қамтамасыз ету механизмдері дами бастады. Алғашқы прокариоттық клеткалар біршама қарапайым катализдік жүйелерге ие болған, нәтижесінде оларды энергйямен қамтамасыз ету ашуға негізделуі мүмкін деп болжайды. Әріқарай прокариоттық клеткалардың жекеленген түрлері ашудан тыныс алуға көшті, ол біршама тиімді энергия алуға мүмкіндік берді. Сонымен, прокариоттық клеткалардың эвоюциялық өзгерістері оларда әртүрлі метаболизмдік жолдардың дамуы бағытында жүрді. Олардың геномы ДНҚ-ның «жалаңаш» молекулаларының қалыптасу бағытында дамыды.
Экариоттық клеткалардың эвоюциялық өзгерістері пішіні, мөлшерлері, құрылымы мен қызметтерінің бір мезгілде биохимиялық жүйелердің өсуі және барлық клеткалар үшін жалпы аэробты метаболизмнің сақталуы бағытында жүрді. Экариоттық клеткалар бұдан 1 млрд жылдай бұрын пайда болған деп санайды және олардың шығу тегін түсіндіру үшін үш болжамды ұсынады.
Осы болжамдардың біреуіне сәйкес (клеткалық симбиоз болжамы), ол біршама кең тараған, эукариоттық клетка әртүрлі типтегі бірнеше клеткалардан тұратын, ортақ клеткалық мембранамен біріккен симбиоздық құрылым болып табылады. Мысалы, қазіргі жасыл өсімдіктердің клеткаларының пластидтері қазіргі цианобактериялардың ата-тегі болып табылатын және аэробты фотосинтезге қабілетті бактериялардан шыққан деп жорамалдайды, ал эукариоттық клеткалардың митохондриялары аэробты бактериялардан бастау алады, олар фотосинтезге қабілетті қарапайым анаэробты клеткалармен симбиозға түскен, оттек атмосферасында тіршілік етуге және оттекті тыныс алу жолымен пайдалануға қабілетті клеткалардың түзілуіне алып келді. Ядроға келсек, ол бастапқы клеткаға енгеннен кейін өзінің цитоплазмасын жоғалтқан, әйтәуір бір көне клеткаішілік симбионттың қалдығы болуы мүмкін деп болжайды. Бұл болжамның пайдасына қазіргі кейбір организмдердің симбиоздық өзара қарым-қатынасы жайындағы деректер дәлел бола алады. Мысалы, бірклеткалы жасыл балдыр хлорелла (Chlorella) жасыл парамецияның (Paramecium bussaria) цитоплазмасында тіршілік етеді. Фотосинтезге қабілетті болғандықтан ол парамецияны қоректік заттармен жабдықтайды. Пластидтер мен митохондриялар ДНҚ, мРНҚ, рРНҚ, тРНҚ және сэйкес ферменттер түрінде белоктар синтезі жайында өздеріне тән генетикалық ақпараттар жүйесіне ие болады. Хлоропластар, митохондриялар мен прокариот-клеткаларына репродукция (олардың барлығы бірдей қарапайым екіге бөліну жолымен көбейеді) тәсілерінің ұқсастығы тән. Ақырында, митохондриялдық гендердің мутациясы ядролық гендердің мутациясына тәуелді емес.
Екінші болжамға сәйкес эукариоттық клетка клеткалық мембранаға бекінген, бірнеше геномдары бар прокариоттық клеткадан пайда болған деп санайды. Клеткалық мембрананың инвагинациясы нэтижесінде бастапқыда фотосинтезге қабілетті мезосомалар түзілген. Бірақ әрі қарай бұл органеллалардың мамандануы жүрді, нәтижесінде олардың біреуі тыныс алу және фотосинтездеу қызметтерінен айырылып, ядроға айналды, басқалары, керісінше, осы қызметтерді әрі қарай дамытып жануарларда митохондрияларға және өсімдіктерде пластидтерге айналды. Бұл болжамның пайдасынаядроның мембраналарының, митохондриялар мен пластидтердің қос құрылысы жайындағы деректер дәлел бола алады.
Үшінші болжамға сәйкес барлық тірі формалар анаэробты ферментативтік гетеротрофтардың арғы тегінен шықты деген ойға негізделген, эукариоттар қабырғасыз (анаэробты) прокариоттардың сублиниясы болып табылады, олар эндоцитозға қабілеттілігін дамытты. Басқа прокариоттарды «жұту» арқылы бұлар оларға қосымша метаболизмдік қабілет берді және кейбіреулері, ең соңында, органелларға, ал қарапайым клетка (уркариот) экариоттық клеткаға айналды. Сонымен, прокариоттар эукариот-клеткаларға қарағанда көне, қарапайым.
Төртінші болжамға сәйкес эукариоттық клеткалар көптеген гендері бар прокариоттық клеткадан пайда болған, олар бөліктерге бөлініп әртүрлі қызмет атқаратын құрылымдардың бастамасын берген деп жорамалдайды. Әрі қарай ұқсас қызмет атқаратын құрылымдарды клондау жүрді, одан соң олар қос мембранамен жабылды, осылайша ядро, митохондриялар түзілуге, ал соңынан мембраналықтор құрылуға алып келді. Бұл болжамның пайдасына ядро мен митохондриялы ДНҚ-ның генетикалық кодының ұқсастығы, сонымен қатар тыныс алу қызметін ядролар мен митохондриялардың реттеуінің ұқсастығы жайындағы деректер дәлел болады.
Жоғарыда айтылғандай, қазір эукариоттық клеткалардың пайда болуының симбиоздық болжамы кең тараған. Бірақ бұл болжамды ескере отырып, митохондриялар мен хлоропластардың қазіргі аэробты бактериялар және цианобактериялармен (сэйкесінше) ұқсас болуына қарамастан, олардан біршама ерекшеліктері болатынын да естен шығармауымыз керек. Мысалы, митохондриялар мен хлоропластарда ДНҚ біршама аз. Демек, бұл жерде митохондриялар мен хлоропластар эволюция барысында өздерінің мөлшері бойынша біршама өзгерістерге ұшыраған.
Эукариоттық клеткалардың геномы әрі қарай ДНҚ молекулаларының белоктармен бірігуі және хроматин мен әртүрлі формадағы және әртүрлі санды хромосомалардың қалыптасуы бағытында дамыды. Хроматиннің мамандануы эухроматин мен гетерохроматиннің қалыптасуында да, аутосомалар мен жыныс хромосомаларының қалыптасуында да көрінеді. Хромосомалардың санына келсек, олардың эволюциялық беталысын түсіндіру әзірше қиын, өйткені көптеген қарапайым организмдердің клеткаларында, жоғарғы эволюциялық сатыда тұрған организмдермен салыстырғанда, көп хромосомалар болады. Бірақ сөз жоқ эволюция барысында кариотиптердің сандық және құрылымдық өзгерістері түр түзілуде маңызды рөл атқарды. Осымен бір мезгілде клеткалық компоненттер құрылымы мен қызметтерінің, реттеуші механизмдердің дамуының күрделенуі қоса жүрді.
Митоздың эволюциялық маңызы күмәнсіз. Митоздың нәтижесінде хромосомалардың бөлінуіні мен орналасуы дэлділігі көпклеткалықты қамтамасыз ететін жағдай болып табылады. Бірақ митоздың өзінің пайда болуының толықтай түсінігі жоқ. Тек ол қарапайым митоздан дамыған деп болжайды, репликацияланған хромосомалардың ажырауы ұршықтың созылуы және жыртылуынан кейін ядролық мембрананың еш зақымданбай-ақ жүзеге асатын механизм болып табылады (жоғарыда айтылған)
Ұлпалардың эволюциясын түсіндіру күрделілігі сол, олар эволюциялық баспалдақтың әртүрлі сатыларында тұрған тірі организмдерге тиесілі ұлпалардың құрылысының бірдей болуымен байланыстығында. Мысалы, буынаяқтылардың, кейбір моллюскалар мен омыртқалылардың бұлшық ет талшықтары бірдей құрылысты. Сонымен қатар бұл организмдер филогенетикалық жағынан өте «алыста» орналасқан. Осындай жағдай әртүрлі таксономиялық топтағы өсімдіктердің ұлпаларын салыстырғанда да орын алады.
Эволюция . Мысалы, вольвокстакейде 5-нан аса клеткалардантұратын, , . Мысалы, (колонияның) түзілуінежауапты. , , .
, , олардыңмамандануы суды . - , оларсыртқыжәнеішкі эпителий . Сыртқы эпителий бар атқышклеткаларболса, ал ішкі эпителий . эпителий мен .
A. A. Заварзинге (1886-1945) тиесілі, тек , де . . А. параллель . А. А. .
Тіндердің пайда болуы(Заварзин)
- Ежелгі тканьдер - Дәнекер және эпителий тканьдері
- Жаңа, мамандалған тканьдері - Бұлшық ет және жүйке тканьдері
Тканьдер жіктелісі
Тканьдердің жіктелуі бірнеше түрі бар.
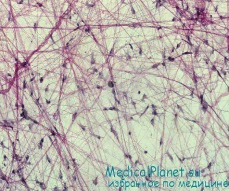
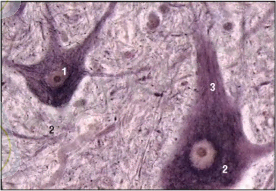
Эпителиалий тіндері пласттарға жасушалардың біріктірілуімен сипатталады. Бұл тіндер арқылы организм мен сыртқы орта арасында зат алмасу жүзеге асырылады. Эпителий тіндеріқорғаныш, сіңіружәне экскреция қызметтерінатқарады. Эпителий - эктодерма, мезодерма және энтодерма болыптабылады.
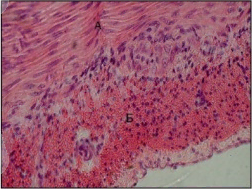
Ішкі орта тіндері (дәнекертін, қаңқа, қанжәне лимфа қосқанда) мезенхима . Ішкі орта . Олартрофикалық (қоректік), пластикалық, .
Бұлшықеттіндері . (көлденеңжолақтытін) жәнемезенхимадан () дамиды.
Жүйкетіні , (регуляторлық) қызметатқарады.
Тканьдік гомеостаз, эпидермистің тұрақтылығын қамтамасыз ететін факторлар
Әрбір тін өзінің құрылымдық бірлігімен, тұрақтылығымен(гомеостаз) және өзінің санымен ерекшеленеді. Тіндік гомеостаздың негізін жасушаның жаңаруы және оның өлімі қамтамасыз етеді.
Эпимидермістің тұрақтылығы мен құрамы осы механизмдер ақылы реттеледі. Алайда, эпидермис тіндердің саралау бірегей мүмкіндігі бар шекара болып табылады, бұл шын мәнінде, оған гомеостаз бетінен корниоциттер бағытталған жасуша көші-қон және дифференциалауы сияқты маңызды құбылыс болып табылады.
 Осылайша эпидермистің гомеостазын келесі процесстер қамтамасыз етеді:
Осылайша эпидермистің гомеостазын келесі процесстер қамтамасыз етеді:
Сіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz