Клетканың құрылымы: плазмалық мембрана, цитоплазма, органоидтар мен клеткааралық байланыстар

ЖОСПАР
Кіріспе 3
Негізгі бөлім
І. Плазмалық мембрана
1. 1 Гиалоплазма 5
1. 2 Клеткалық мембраналар 6
1. 3 Клетка бетінің мамандалған құрылымдары 8
1. 4 Клеткааралық байланыстар 9
ІІ. Клетканың негізгі органоидтары
2. 1 Эндоплазмалық тор 12
2. 2 Гольджи кешені 13
2. 3 Лизосомалар 16
2. 4 Пероксисомалар (микроденешіктер) 18
2. 5 Рибосомалар 18
2. 6 Митохоңдриялар 19
2. 7 Хлоропластлар 23
Қорытынды 27
Қолданылған әдебиеттер тізімі 28
Кіріспе
Цитология жеке ғылым ретінде өткен ғасырдың соңғы ширегінде пайда болған. Бірақ клетка жөніңдегі ілім XVII ғасырдан басталады, оның тарихы үш ғасырды камтиды. Организмнің клеткалық құрылысының ашылуы микроскоптың шығуьша байланысты.
Микроскоптың шығу тарихы толық анық емес. Бірақ та микроскопты жасап шығаруца көзілдірік өндірісінің кейбір әсерінің болғаны күмәнсіз. Көзілдірік 1285 жьшы Италияда шыққан. Кейбір аңыздарға карағанда алғашқы микроскопты голландиялық оптиктер Янсендер 15. 90 жылы жасап шығарған. 1612 жылы Галлилей де микроскопты құрастырған. Алғашқы микроскоп ғылыми зерттеу құралы болмаған, оған ойыншық ретінде қараған. Ағылшын математигі, физигі және механигі Роберт Гук 1665 жылы өзі жасаған микроскоп арқылы тығынның құрылысын қарап, оньщ ұяшықтардан тұратынын анықтаған. Осы ұяшықтарды клетка деп атаған. Сонымен Р. Гук "клетка" деген терминді қалдырған. Бұл қазіргі түсініктегі клетканың ашылуы емес. Кейінірек, осы XVII ғасырда Грю мен Мальпиги микроскопты қолдана отырып өсімдіктердің құрылысын зерттеген.
Чех ғалымы Пуркинье 1840 жылы клетканың ішкі құрамын белгілеу үшін «протоплазма» деген терминді ұсынған болатын. Протоплазма - клетканы құрайтын тірі зат - кариотазма (ядро деген мағынада) мен цитоплазмаға бөлінеді. Бұл екі терминді ғылымға енгізген 1882 жылы полыналық ботаник Страсбургер. Ядродан басқа клетканың барлық ішкі құрамды бөлігін цитоплазма деп атайды.
І. Плазмалық мембрана
Плазмалық мембрана, немесе плазмалемма (гректің plasma - пішін, lemma - кабықша) мембраналардың ішінде ерекше орын алады; клетканы сыртынан қоршап, оның сыртқы ортамен тікелей байланысуын қамтамасыз етеді. Плазмалемманың қалыңдығы 7, 5 нм. Жарық микроскопының шешуші қабілеті кем болғандықтан, плазмалық мембрана электрондық микроскоп пен электрондық микроскопиялық зерттеулерге материалды дайындау әдістері шыққаннан кейін ғана байқалды
Плазмалық мембрананың негізгі құрамды белігі липидтер (40% жуық), белоктар (60% жуық) және көмірсулар (10% жуық) . Плазмалемма басқа мембраналарға қарағанда холестеринге бай келеді; оның фосфолиптерінде қаныққан май қышқылдары басым болады. Плазмалық мембрананың құрамына кіретін белоктардың құрамы атқаратын қызметтерінің көп түрлі болуына байланысты алуан түрлі болады. Плазмалық мембрананың құрамында 30-ға жуық түрлі ферменттер анықталған, ал жиі кездесетіндері Na + және К + иондарымен белсенділігін артыратын Mg-re тәуелді АТФ-аза, 5ä-нуклеотидаза, сілтілі және қышқыл фосфатаза, аденилатциклаза, РНК-аза. Эукариондық клеткалардың плазмалық мембранасында тыныс алу тізбектерінің фермеиттері мен гликолиздеуші ферменттер болмайды, ал прокариондық клеткаларда тотықтырып фосфорлау мен электронды тасымалдаушы тізбектің элементтері плазмалық мембранасында орналасқан. Аталған ферненттердің ішінде ең маңыздысы Na + және К + иондарымен белсенділігін артыратын Mg-re тәуелді АТФ-аза, себебі иондардың плазмалық мембранадан иондардың өтуінде маңызды рөл атқарады.
Плазмалемманың сыртқы бетінде мембрананың кұрамына кіретін белоктармен байланысқан күрделі қосылыстарды - гликопротеидтер мен гликолипидтерді құрайтын көмірсулардың молекулалары болады. Бұл мембрана үстіндегі комплекс, немесе гликокаликс (грекше kalyx - кабықша), бір және көпклеткалы жануарлардың клеткаларында жақсы жетілген. Сол сияқты өсімдіктердің клеткаларында да кездеседі. Гликокаликс сыртқы ортамен тікелей қатынаста болғандықтан, клеткалардың беткі аппаратының рецепциялық функциясында маңызды рөл атқарады. Плазмалық мембрананың бос беттерінде гликокаликстің көмірсу компоненті гликозильдік топтар антенналарға ұқсас тармақталған олигосахаридтік тізбектер кұрап, сырттан келген сигналдардың ажырауын камтамасыз етеді. Гликокаликс деген терминді 1963 жылы Беннет ұсынған болатын.
Гликокаликсте түрлі заттардың диффузиясының жылдамдығы кемиді. Са ++ немесе Mg + иондарының көмегімен гликокаликс зонасымен байланысқан, клеткадан бөлінген ферменттер болуы мүмкін. Бұл ферменттер түрлі заттардың (полисахаридтердің, белоктардың, майлардың тағы басқалардың) клеткадан тыс ыдырауына қатысады. Осы ыдырау өнімдері мономерлер күйінде плазмалық мембрана арқылы өтіп, клеткаға сіңеді.
Плазмалық мембрана клетканың ішкі кұрылымының құрамын да реттейді. Барлық клеткаларда плазмалық мембрана жаңарып отырады. Плазмалемманың ішкі бетіне жанасып, цитоплазманың беткі бөлігі немесе кортикалық (грекше cortex - қабық) қабат жатады. Кортикалық қабатта органеллалар болмайды, оның есесіне цитоплазманың фибриллалық элементтері - микрофиламенттер мен микротүтікшелер көп кездеседі. Кортикалық қабаттың негізгі фибриллалық компоненті актиндік микрофибриллалардың торы. Кортикалық қабат клетка бетінің түрлі өсінділерін құрайды. Жарық микроскопында тегіс болып көрінетін клеткалардың бос беті шынында ирек-ирек болады. Кейбір клеткалардың бетінде саңылау мен ойыстар, екіншілерінде псевдоподиялар (жалған аяқтар) деп аталатын өсінділер, үшіншілерінде микробүрлер мен кірпікшелер болады. Бетінде катпарлар болатын клеткалар да кездеседі. Тірі клеткалардың беті толқындалып үнемі козғалып тұрады, дірілдеп тұрған сияқты болып байқалады. Плазмалық мембрана көптеген маңызды биологиялық функциялар атқарады. Олардың ішіндегі ең бастылары - қорғау функциясы мен тасымалдау функциясы. Тасымалдау функциясына су мен иондарды және төменгі молекулалық заттары пассивті түрде тасымалдау мен осы заттарды концентрацияның градиенттеріне қарсы белсенді түрде өткізу және жоғары молекулалық қосылыстар мен комплекстерді тасымалдаудың түрлі формалары жатады; клеткада пайда болған өнімдерді плазмалық мембрана клеткалардан сыртқа шығаруға қатысады. Сонымен бірге биополимерлердің клетка ішінде ыдырау процесіне де қатысады. Плазмалемманың бетінде клетканың ішіне хабар өткізетін түрлі рецепторлар орналасқан.
1. 1 Гиалоплазма
Гиалоплазма (грекше hyaline - мөлдір), негізгі плазма, немесе цитоплазманың матриксі, клетканың маңызды бөлігі, оның шын мағынасындағы ішкі ортасы. Электрондық микроскопта цитоплазманың матриксі гомогенді немесе электрондық тығыздығы төмен ұсақ дәнді зат болып байқалады.
Гиалоплазманың құрамында түрлі глобулалық белоктар мен цитоплазмалық матрикстің ферменттері болады. Бұлар эукариондық клеткадағы белоктардың жалпы санының 20-25%-ін құрайды. Матрикстің маңызды ферменттеріне гликолиздің ферменттері, қанттардың, азоттық негіздердің, аминқышқылдарының, липидтердің және басқа маңызды қосылыстардың метаболизмінін ферменттері жатады. Матриксте РНҚ мен белоктың синтезделуі кезінде аминқышкылдарының белсенділігін арттыратын ферменттер де орналасқан. Клетканың осмостық және буфферлік қасиеттері де гиалоплазманың кұрамы мен құрылымына байланысты.
Гиалоплазма барлық клеткалық құрылымдарды біріктіріп тұрады және олардың бір-бірімен химиялық әрекеттесуін қамтамасыз етеді. Аминқышқылдарының, май қышқылдарының, нуклеотидтердің, қанттардың клетка ішіндегі тасымалдануын да іске асырады. Гиалоплазмада плазмалық мембранаға және онан кері, митохондрияларға, ядро мен вакуольдарға, иондардың ағысы үнемі жүріп отырады. Гиалоплазма АТФ молекулаларының жинақталатын орны. Мұнда гликоген, май тамшылары сияқты қоректік заттар да жиналады. Зерттеу әдістерінің дамуы цитоплазмалық матрикстің макромолекулалық құрылысының әртүрлі клетканың өзінде әркелкі ғана емес, бір клетканың өзінде де түрліше болатынын көрсетті.
1. 2 Клеткалық мембраналар
1955 жылдан бастап клетканы коршаушы мембраналардан басқа эукариондық клеткалардың бәрінде клетканың ішінде орналасқан субмикроскопиялық күрделі жүйе болатыны анықталды.
Цитомембраналар клетка құрылымының негізгі компоненттерінін бірі. Цитомембраналар жеке элементтерден тұратын, бір-бірімен өзара әрекеттесіп біртұтас күрделі жүйе құрайтын, клетканың тіршілігінде маңызды рөл атқаратын құрылым.
Шығу тегіне, құрылысына және атқаратын қызметіне қарай мембраналарды бірнеше түрлі топтарға бөледі: плазмалемма (плазмалық мембрана, сыртқы клеткалық мембрана), ядролық мембрана (ядро қабықшасы), миелин қабықшалары - мембраналары, вирустар мен бактериялардың мембраналары, эндоплазмалық тор, митохондриялар мен хлоропласидтердің мембраналары.
Құрылысы мен құрамы жағынан мембраналардың бәрінің көптеген жалпы қасиеттері болады: олардың орташа қалыңдығы 7 нм-ге тең, белоктар мен липидтерден тұрады және олар өткізгіштіктің тосқауылы; мембраналардың, үзінділерінің ұштары ұштасып көпіршіктер құрайды. Көптеген жалпы қасиеттерімен бірге мембраналардың белгілі айырмашылықтары да бар. Мембрананы құрауда негізгі рөл атқаратын глобулалық (шар) кұрылысты интегралдық және жартылай интегралдық белоктар. Белоктардың мөлшері мембраналардың типіне байланысты айнымалы болады. Миелиндік мембранада белок аз, ал көлденең салалы бұлшық ет талшықтарында 65% белоктар және 15% липидтер болады. Бауыр клеткаларының мембраналарының 85% белоктардан тұрады, ал фосфолипидтер мен холестерин 10% кұрайды.
Мембраналардың липидтері фосфолипидтер, гликолипидтер мен стеролдар. Фосфолипидтердің молекулалары полярлы басынан және екі бейполярлы кұйрықтардан тұрады. Гликолипидтер липидтер мен көмірсудың қосылысы болып есептеледі. Фосфолипидтер сияқты полярлы басы мен бейполярлы құйрықтардан тұрады. Стеролдар стероидтар класына жататын спирттер. Бұлардың ішіндегі ең көп тарағаны холестерол. Бұның молекуласы толықтай бейполярлы; осы белгісі жағынан фосфолипидтер мен гликолипидтерден өзге болады. Түрлі мембранадағы фосфолипидтердің саны әркелкі, олардың түрліше болуы мембраналардың кұрылысына әсерін тигізеді. Фосфолипидтердің химиялық құрамының маңызы зор, әдетте олар фосфатидилхолинге бай келеді. Сонымен бірге мембраналардың көпшілігінде көмірсулардың көптеген саны болады.
Белоктар өзіндік табиғат туғызады, ал ерекшелік өз кезеңінде мембраналардың бір-бірінен ажырататын белгі. Клеткалық мембраналарда мыңдаған түрлі белоктар кездеседі. Бұлардың ішінде таза құрылымдық белоктармен бірге қосымша функцияларды атқаратын белоктар да болады. Кейбіреулері әр түрлі заттарды тасымалдап тасымалдаушылардың, қызметін атқарады. Белоктың молекулаларда, я көршілес белоктық молекулалардың арасында гидрофильді каналдар, немесе поралар болады деп жорамалдайды. Бұл поралар мембрананы тесіп өтіп тұрады.
Мембраналарда ферменттік белоктар, рецепторлар, электрондарды тасымалдаушылар, т. т. болады. Сонымен қатар мембраналарда гликопротеиндер де болады. Белоктар липидтермен қосылып комплекстер кұрайды. Кайсыбір белоктар құрылымдық фосфолипидтермен қосылған кезде өткізгіштіктіңбарьері (тосқауылы) пайда болады; өткізгіштік барьерінің болуы клеткалық мембрананың маңызды функциялық ерекшелігі. Басқа белоктар өздеріне тән молекулалық конфигурациясының арқасында катализдің белсенді зоналарын құрайды.
Биологиялық мембраналардың кейбір маңызды касиеттері липидтік бикабаттың құрылысына байланысты. Олардың бірі мембраналардың қозғалмалығы, оның шегінде молекулалар биқабат жазықтығында еркін қозғалып жүре алады. Липидтік биқабаттың тағы да бір ерекшелігі суда ерігіш молекулалар олар арқылы өте алмайды.
Мембраналар клетканы компартаменттер деп аталатын бірнеше түйық бөлмелерге бөледі. Цитомембранамен шектелген клеткалық бөлмелер әркелкі пішінді болады - каналшықтар, вакуольдер, гранулалар, жалпақ қапшықтар т. б. Клетка ішіндегі мембраналар жалпы есеп бойынша клетканың барлық массасының үштен бірін немесе тең жартысын құрайды. Клеткалық мембраналардың негізін қос липидтік қабат құрайтыны жөніндегі түсінік Овертонның есімімен байланысты.
1895 жылы Овертон молекулалардың клетка ішіне өту жылдамдығы липидтер ерекшелігіне байланысты екенін байқаған. Осы физиологиялық байқаулардан ол мембраналарда липидтік қабаттың болатындығы жөнінде анатомиялық тұжырым жасаған. 30 жылдан кейін 1927 жылы голландиялық Гортер мен Грендель эритроциттердің мембранасын зерттеп бұл түсінікті қуаттаған болатын. 1935 жылы Давсон-Даниелли гипотезасы шыққан. Бұл гипотеза бойынша плазмалық және басқа мембраналар фосфолипид молекулаларының бір-бірінің үстінде орналасқан екі қабатынан тұрады. Осы күрделі липидтік молекулалардың әрқайсысында гидрофильдік (суда ерігіш полярлы) және гидрофобты топтар болады. Бір қабаттың гидрофобты беті екіншісінің гидрофобты бетіне беттеседі. Гидрофильдік полярлы топтар осы екі қабаттың бетін кұрайды. Олар екі белоктық қабаттармен байланысады.
Сонымен бұл гипотеза бойынша клеткалық мембрана үш қабаттан: фосфолипидтік қабаттардан тұратын бір орталық қабаттан және орталық липидтік қабаттық екі бетінде орналасқан екі белоктық кабаттан тұрады. Бұл гипотеза мембраналарды электрондық микроскоппен зерттеген кездегі көріністі түсіндіреді.
Давсон мен Даниеллидің моделі липидтердің орналасуын канағаттанарлық түрде түсіндіргенмен, белоктардың орналасуын түсіндірмейді. Белоктар липидтердің бетінде қабат құрап орналасады деп жорамалдаған болатын. Бірақ, мұндай түсінік егжей-тегжейлі байқауларға сәйкес келмеген, себебі мембраналық белоктардың көпшілігі липидтермен мықты байланыста болады. Мембрана көптеген заттарға диффузиялық тосқауыл кұрайды. Мембрана арқылы жүретін диффузияның жылдамдығы су ерітіндісіне қарағанда 10 8 -10 9 есе төмен. Мембрана арқылы кедергісіз өтетін молекулалар мен иондардың болатыны белгілі. Заттар алмасуының ингибиторлары (бөгеуші заттар) бұл молекулалар мен иондардың мембрана арқылы етуіне әсер етпейді. Осы фактілерді түсіндіру үшін Давсон мен Даниелли 1955 жылы мембраналарда поралар (каналдар) болуы керек деген жорамал жасаған. Бірақ бұл модель мембраналар туралы жиналған байкаулардық бәрін толықтай түсіндіре алмаған.
Электрондық микроскоп клеткалық мембрана екі сыртқы электрондарға тығыз қабаттар мен ақшыл аралық қабаттан тұратын үш қабатты құрылым екенін көрсетті. Үш қабатты клеткалық мембрана элементарлық (жабайы) мембрана деп аталды. Бұл құрылым Даниелли мен Давсонның сэндвич (белок-липид-белок) моделіне сәйкес келеді.
1. 3 Клетка бетінің мамандалған құрылымдары
Плазмалық мембрана және оған жанаса орналасқан цитоплазманың қабаты кейбір клеткаларда түрлі мамандалған құрылымдар құрайды. Бұндай кұрылымдар әдетте жалғасушы клеткалардың жанасу беттерінде (десмосомалар, синапстар т. б. ) немесе клетканың бос бетінде (мысалы, микробүрлер, кірпікшелер мен талшықтар) жиі байқалады.
Орналасу ерекшеліктеріне қарап, клеткалар бетінің мамандалған құрылымдарын клетканың бос (апикальдық) бетіндегі, жанасқан клеткалардың бетіндегі және клеткалардың базальдық бөлігіндегі мамандалған құрылымдар деп бөледі.
Жануарлардың клеткаларының көпшілігінің плазмалық мембранасы түрлі құрылымдық өсімділер құрайды. Солардың бірі - микробүрлер. Электрондық микроскопиялық зерттеулер микробүрлер плазмалық мембранамен шектелген цитоплазманың өсінділері екенін анықтады. Бұл құрылымдар әртүрлі қызметтер атқарады. Микробүрлердің негізгі функциясы - клеткалардың ауданын арттыру. Сондықтан да бұлар алмасуға белсенді қатысатын клеткалардың бетінде көп болады. Мысалы, асқорыту жолының кілегейлі қабатын астарлаушы клеткаларда, бүйректің каналшықтарында, секрет бөлуші клеткаларда. Дәнекер ұлпалық клеткаларда да байқалады (фибробласттарда, лейкоциттерде) . Өсімдік клеткаларында микробүрлер болмайды. Микробүрлердің калыңдығы 100 нм шамасында. Саны мен ұзындығы клеткалардың түрлі типінде түрліше. Әсіресе, ішек және бүйрек эпителийінің тарақша көмкермесінде (щеточная клетка) көп болып байқалады. Тарақша көмкерме дегеніміз, клетканың апикальдік бетіндегі микробүрлердің жиынтығы. Ішек эпителийінің бір клеткасында 3000 дейін микробүрлер болады. Ішектің 1 мм 2 бетіндегі микробүрлердің саны 200. 000. 000 дейін жетеді. Микробүрлердің аралығында гликокаликстің қалың қабаты орналасқан. Бұл аймақта сіңіру процесіне қатысатын ферменттер (пептидазалар, сілтілі фосфатаза, дисахарозалар) болады. Микробүрлердің жиырыла алатын қабілеті бар. Микробүрлер аз болса да клеткалардың басқа түрлерінде де кездеседі. Мысалы, мезотелийдің клеткаларында, өт қабының, жатырдың, сары уыз қапшығының, бауыр клеткаларының т. т. эпителийінде.
Плазмалық мембрананың өсінділерінің тағы да бір типі кірпікшелер мен талшықтар. Бұлардың жуандығы (диаметрі 0, 2 мкм жуық) мен ультрақұрылымы бірдей. Сондықтан, қимыл органеллаларының осы екі типін біріктіріп ундулиподия деп атайды. Кірпікшелер мен талшықтар плазмалық мембранамен қапталған, базальдық денешікпен байланысқан микротүтікшелердің жүйесінен тұрады. Кірпікшелер мен талшықтар жануарлар клеткаларында кең тараған; өсімдіктерде аталық жыныс клеткаларында кездеседі; жалаңаш тұқымдылар мен жабық тұқымдыларда бұлар болмайды. Инфузориялардың әрбір клеткасында жүздеген, мыңдаған кірпікше болады.
Кірпікшелер мен талшықтар ұзындығы және клеткадағы саны бойынша ажырайды. Кірпікшелердің ұзындығы 5-10 мкм, талшықтардікі 150 мкм және онан да артады.
1. 4 Клеткааралық байланыстар
Көпклеткалы жануарлар организмдерінің органдары мен ұлпаларының құрамындағы клеткалардың өзара байланысы клеткааралық байланыстар деп аталатын күрделі арнаулы құрылымдар кұрайды.
Клеткааралық байланыстардың бірнеше түрлерін ажыратуға болады: жай клеткааралық байланыс (құлып және саусақ тәрізді байланыстар), десмосомалық, тығыз байланыстар, лента тәрізді десмосомалар, санылау тәрізді байланыс (1-сурет) .
Жай клеткалық байланыстар. Қабат құрайтын клеткалар ерекше құрылымдарды құрамастан өзара байланысады. Бұндағы клеткалардың байланысы жай байланыс арқылы камтамасыз етіледі. Екі клетканың плазмалеммаларының арасында 15-20 нм клеткааралық кеңістік болады. Жай байланыстан басқа клеткаларды өзара байланыстырып бекітетін арнаулы құралдар болады. Ол «құлып» деп аталатын құрылым.
Тығыз байланыс. Екі көршілес клеткалардың плазмалық мембраналары мейлінше жақындап, бір-біріне жабысып, қалындығы 2-3 нм бір жалпы қабат кұрайды. Байланыстың аймағы макромолекулалар мен иондарды өткізбейді. Тығыз байланыс ми қылтамырларының арасында ми клеткаларына қаннан заттардың еркін диффузиялануына кедергі болып гематө-энцефальдық тосқауыл кұрайды. Тығыз байланыс клетканың апикальдік жиегінде болады.
Десмосомалар - эпителийлік клеткалардың көбіне тән. Десмосома (грекше desmos - байланыс, soma - дене) ара қашықтықтары 30-50 мкм екі көршілес клеткалардың плазмалық мембраналарының диаметрі 0, 5 мкм-ден екі дөңгелек немесе сопақша учаскелерінен түзілген. Мембрананың астында клетканың ішінде орналаскдн тоно-фибриллалар бекитін тығыз пластинка болады.
Саңылау тәрізді байланыс, немесе нексустар. Клеткадан клеткаға химиялық заттарды тікелей жеткізуге қатысатын құрылым. Байланыстың бұл түрінде екі көршілес клеткалардың плазмалық мембраналарының арақашыктықтары 2-3 нм-ден аспайды.
Көршілес клеткалардың мембраналары коннексондар деп аталатын құрылымдармен байланысқан. Коннексон - ортасында ені 2 нм каналы бар, молекулалық массасы 20 мындай коннектин белогінен тұратын цилиндр тәрізді агрегат.
Саңылау тәрізді байланыстың зоналарында клеткалардың функциялық ерекшелігіне байланысты 10-20-дан бірнеше мыңға дейін плазмалық мембрананы тесіп өтетін коннексондар болады. Көршілес клеткалардың плазмалық мембранасындағы коннексондар бір-біріне дәл қарсы орналасып біртұтас жүйе құрайды.
Плазмодесмалар . Клеткааралық байланыстың бұл типі есімдіктерде кездеседі. Бұл құрылымды алғаш рет И. Н. Горожанкин (1881) жазған болатын. Плазмодесмалар кершілес екі клетканы байланыстыратын жіңішке түтік тәрізді цитоплазмалық каналдар. Осы каналдарды шектейтін мембрана кершілес клеткалардың плазмалық мембраналарына ауысады. Плазмодесмалар клетканы беліп тұратын клеткалық кабырғаны тесіп өтеді. Сонымен, кейбір өсімдік клеткаларында плаз-модесмалар көршілес клеткалардың. гиалоплазмасын байланыстырып тұрады. Плазмодесмалардың маңызы зор. Қүрамында қоректік зат-тар, иондар және басқа косылыстар бар ерітінділердің клеткааралық ағысын камтамасыз етеді. Сонымен бірге плазмодесма аркылы клет-каларға өсімдіктер вирустары да таралады.
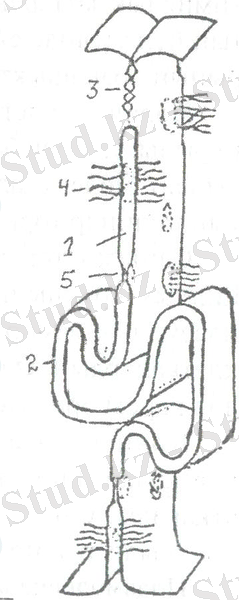
1-сурет. Клеткааралық байланыстар құрылысының схемасы (Ю. Ченцовтан) : 1 - жай байланыс; 2 - кұлып; 3 - тығыз тұйықтаушы байланыс;
4 - десмосома; 5 - саңылау тәрізді байланыс.
ІІ. Клетканың негізгі органоидтары
2. 1 Эндоплазмалық тор
1945 жылы Портер, Клод және Фуллман фибробластлердің өте жұқа кесіндісін электрондық микроскоппен қарап ерекше торды байғаған. Бұл құрылымды атау үшін эргастоплазма (Дальтон және басқалар), вакуольдық жүйе (Де Робертс), α және β - цитомембраналар (Шестранд) деген терминдер кездеседі. 1953 жылы Портер оны эндоплазмалықтор (эндоплазмалық ретикулум) деп атаған. Кейін эритроциттер мен бактериялардан басқа клеткалардың бәрінде де байқалған. Бұлшық ет клеткаларыңдағы эндоплазмалық торды саркоплазмалықтор деп атайды, ол миофибриллаларды қоршап тұрады, жиырылу процесіне тікелей қатынасы бар.
Эндоплазмалықтордың құрылысы мен жетілу дәрежесі түрлі клеткаларда түрліше болады. Эндоплазмалық тор белоктық алмасуы жоғары секрет белуші клеткаларда жақсы жетілген (ұйқы безінің ацинустарының клеткаларында, бауыр клеткаларында) . Сперматоциттерде, лейкоциттерде, ішек крипталарының эпителийінде, бүйрекүсті бездің қабық бөлігінің клеткаларында эндоплазмалық тор нашар жетілген. Көптеген байқаулар бойынша эндоплазмалықтордың жетілу жеделдігі клетканың жіктелу дәрежесіне тәуелді болуы да мүмкін (Портер, 1963) . Эндоплазмалық тордың жіктелу жеделділігі мен пішіні клетканың функциялық белсенділігіне байланысты өзгереді.
Эндоплазмалық тордың клеткалық кабырғасы, плазмалық және басқа клеткалық мембраналар сияқты, липопротеиндік мембранадан тұрады. Бірақ эндоплазмалықтордың мембранасы жұқа (5-6 нм), тегіс және өткізгіштігі түрліше болады, ал биқабаттағы фосфолипидтің құрылысы өзге, ал олармен байланысты холестерин жоктың қасында, сол сияқты белоктарда өзгеше.
Эндоплазмалық тор мен ядро қабықшасының мембраналарының өзара айырмашылығы ядро қабықшасында «поралар» деп аталатын тесіктердің болуы. Эндоплазмалық торда поралар комплексі болмайды.
Эндоплазмалықтордың екі түрін ажыратады:
1) Бұдыр, немесе гранулалық;
2) тегіс, немесе агранулалық.
Гранулалық эндоплазмалық тордың сыртқы қабырғасында рибо-сомалар жайғасқан . Гранулалық эндоплазмалық тор кейде эргастоплазма деп аталатын цитоплазманың базофильдік учаскесіне сәйкес келеді. Бұл клеткалардың цитоплазмасының базофильдігі ондағы РНК көп болуына байланысты.
Эндоплазмалық тордың мембраналарымен байланыскан рибосомалар клеткадан шығарылатын белоктарды синтездеуге қатысады. Сонымен бірге клетка ішіндегі астың корытылуына қажет белоктар - ферменттерді синтездейді. Эндоплазмалық тордың қуыстарында жиналған белоктар Гольджи аппаратының вакуольдарына жеткізіледі, онан олар басқа вакуольдарға ауысады немесе клеткадан шығарылады.
... жалғасыСіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz