Микроағзалардың өнеркәсіптік штамдарын алу: селекция, индуцирленген мутагенез және рекомбинантты ДНҚ әдістері

СЕМЕЙ қаласының ШӘКӘРІМ атындағы МЕМЛЕКЕТТІК УНИВЕРСИТЕТІ
ПОӘК
«Биотехнология негіздері»
пәні бойынша оқу - әдістемелік бағдарламасы
№ 1 - басылым
18. 09. 2013ж.
«БИОТЕХНОЛОГИЯ НЕГІЗДЕРІ»
пәнінің ОҚУ-ӘДІСТЕМЕЛІК КЕШЕНІ
5В070100 - «Биотехнология» мамандығы үшін ОҚУ-ӘДІСТЕМЕЛІК КЕШЕНІ БАҒДАРЛАМАСЫ Семей 2013Дәріс № 3. Селекция әдістерімен алынған, микроағзалардың өнеркәсіптік штамдары. Индуцирленген мутагенез. Мақсатты өнімнің құрылымдық ұқсастықтарына орнықтылығы бойынша - продуценттерді іріктеу.
Селекция - ДНК нуклеотидті тізбектілігінде құрылымдық түрөзгергіштің салдарынан, тұқымқұалаушылығы секіріс тәрізді өзгерістерге ұшыраған, ағзаларды, яғни, мутанттарды бағытты іріктеу. Селекцияның басты жолы - бұл продуценттерді көз жұмып іріктеуден оардың геномдарын саналы құрастыруға дейінгі жол. Бірақ кенеттен мутацияларды іріктеуге негізделген, әдістер микроағзаларды пайдаланумен әртүрлі технологиялардың дамуында маңызды рөл атқарады.
Осындай жолмен ұзақ уақыт бойында сыра, шарап, наубайхана ашытқыларының штамдары, сіркеқышқылының, пропион-қышқылының бактериялары т. б. іріктелген болатын. Баспалдақты іріктеу туралы сөз қозғалып отыр: кезңдердің әрқайсысында микроағзалардың популяцияларынан ең жоғары тиімді клондар іріктеледі. Кенеттен түрөзгеруге негізделген, селекция әдісінің шектеулігі олардың төмен тиілігімен байланысқан, бұл үдерістің қарқындылығын мәнді түрде қиындатады. ДНК құрылымында өзгерістер сирек жүреді. Түрөзгергіштік туындау үшін, ген орташа алғанда - есе еселенуі керек.
Көптеген ұрпақтар бойында үлкен көлемдерде бионысанды үздіксіз егумен сәйкестікте, 1мл тамшыларға және одан артық жасушаларға жететін, микробты популяциялардың жоғары тығыздықтары мутанттардың жеткілікті үлкен мөлшерін алуға мүмкіндік береді. Үздіксіз тәртіпте егу барысында ең өнімділікті мутанттарды іріктеудің мысалы, ашытқылар тіршілігінің өніміне, этанолға төзімділік белгісі бойынша Saccharomyces uvarum ашытқыларды іріктеу болып табылады. Қышқылдарға және сілтілерге, метаболизм өнімдеріне, ауыр металлдардың иондарына, т. б. -әртүрлі факторларға бионысандардың төзімділігін жоғарылатуға жол алатын, бұл қадамның жаңашылдығы-дақылдың бөлуімен және биореакторға ингибирлейтін фактордың (бұл жағдайда этанол) түсуімен-дақылдың тіршілігін сипаттайтын, параметрдің арасында кері байланысты орнатуда. Осындай белгілеудегі ұзақ (650 сағаттық) егуде, 10%-ға дейін концентрацияларда этанолдық ингибирлейтін әрекетіне резистентті, мутант ашытқылар алынған.
Геномның жасанды зақымдануында бионысан мутациялары жиілігінің күрт ұлғаюы-индуцирленген мутагенез селекцияның мәнді түрде жақсаруына әкеледі. ДНК-ның бірінші құрылымының өзгерістерін тудыратын, кейбір химиялық қосылыстар, ультракүлгін рентген немесе усәулелері мутагенді әсерге ие болған. Өздерін ұсына білген мутагендер қатарына азотты қышқыл, алкилдеуші агенттер, акридинді бояғыштар, бром - урацил, т. б. жатады.
Алынған клондардың тоталді тексерейін (скрипинг) жүргізіді. Ең өнімділікті клондарды іріктеп алып, сол немесе өзге мутагенмен өңдеуді қайталайды, ең өнімділікті нұсқаны қайта таңдайды, т. с. с., яғни, мұнда да керекті белгі бойынша баспалдақты іріктеу туралы сөз қозғалады.
Еңбегінің күрделілігі - индуцирленген мутагнез және тізбекті баспалдақты іріктеу әдісінің негізгі кемшілігі. Сондай - ақ мутациялардың сипаты туралы мәліметтердің болмауы да әдістің кемшілігіне жатады, зерттеуші ақтық нәтиже бойынша іріктеу жүргізіледі. Егер, ауыр металдарға төзімді, бактерия штамдары туралы сөз қозғалса, онда төзімділік әртүрлі мутация топтарына байланысты болуы мүмкін: а) бактерия жасушасымен металдар катиондарының жұтылу жүйесін басумен; б) жасушадан жұтылған катиондардың шығарылуын белсендірумен; в) ауыр металдардың ингибирлейтін әсеріне сезімтал, жүйелердің қайта құрылуымен.
Молекулалық генетикалық жетістіктері, мақсатты өнімнің құрылымдық аналогтарына төзімділіктері бойынша, продуценттерді іріктеудің мақсатқа сай әдістерін тәжірибеге енгізуге мүмкіндік берді. Әдіс, биосинтетикалық жолдың ақтық өнімімен кері байланыстың қағидасы бойынша, ферменттерді реттеуге негізделген (В. Г. Дебабов, 1984) . Метаболит концентрациясының жоғарылауы, метаболит синтезіне қатысатын, ферменттің белсенділігін ингиьирлейді; немесе бұл ферменттің синтезін репрессиялайды. Осылайша, пен глюкоза болғанда көптеген бактериялардың жасушалары тіршілікке қажетті барлық азотқұрамды қосылыстарды синтездейді. Егер ортаға сол немесе өзге аминқышқылын қосса, онда оның

Сурет 2. Ақтық өніммен биосинтетикалық жолдың негативті реттелуі: A, B, C, D-Ea, Eb, Ec, Ed ферменттерімен, өршітілетін реакциялардың өнімдері.
Синтезі тез тоқтайды. Метаболиттің құрылымдық аналогтары да осындай әсер тудырады, бірақ олар метаболитті функционалды алмастыра алмайды. Мысалы, аминқышқылының аналогы ақуыз құрамына кіре алмайды, сондықтан аналогтың ақуыз құрамына кіре алмайды, сондықтан аналогтың қатысуында қалыпты жасушалардың өсуі, мақсатты өнімге ашығумен байланысты, басылады.

Бұл жағдайларда тек кейбір жасушалар ғана аман қалады. Бұл ферменттер синтезінің және белсенділігінің бұзылған реттелуімен мутенттар. 1) Еа ферменті (сурет 2) функционалдық белсенділігін сақтаған, бірақ ақтық өнімнің немесе оның аналогының ингибирлейтін әсеріне сезімталдылығын жоғалтқан; 2) Еа ферментінің синтезі табиғатта өсімдіктердің обыр ісіктерін тудырады, өйткені олар бақылаусыз көбейеді. Осындай клондардың негізінде құнды қосылыстарды алуға болады.
Дәріс № 4. Генетикалық инженерия әдістерімен алынған, микроағзалардың өнеркәсіптік штамдары.
1 Қажетті генді алу
2 Оны, репликацияға қабілетті, генетикалық элементке ( векторға) құру.
3 Реципиент - ағзаға, вектор құрамына кіретін, генді енгізу.
4 Қажетті ген ие болған, жасушаларды сәйкестендіру (скрининг және селекция)
Қазіргі биотехнологияны көбінесе генетикалық инженерия негізіндегі биотехнология ретінде сипаттайды. Шындығында, бұл жасанды жасалған генетикалық бағдарламаларды жүргізудің нәтижесінде, бионысандардың бағытталған түрөзгергіштігіне қолданылатын негізгі жол. Кейде генетикалық инженерияның үш деңгейін ажыратады: 1) гендік - жеке гендерден тұратын, рекомбинантты ДНҚ - ымен тікелей манипуляциялау; 2) хромосомдық - гендердіңүлкен топтарымен немесе тұтас хромосомаларымен манипуляциялар; 3) геномдық - генетикалық материалдың барлығын немесе үлкен бөлігін біржасушадан басқасына тасымалдау. Қазіргі түсінікте генетикалық инженерияға рекомбинантты ДНҚ технологиясы кіреді. Генетикалық инженерия облысындағы жұмыстар төрт негізгі кезеңнен тұрады: 1) қажет генді алу; 2) оны репликацияға қабілетті, генетикалық элементке (векторға) құру; 3) реципиент - ағзаға, вектордың құрамына кіретін, генді енгізу; 4) қажетті генге ие болған, жасушаларды сәйкестендіру (скрининг және селекция) . Әрбіркезеңді жеке - жеке қарастырайық. Гендерді алу. Қажетті генді келесі түрмен алуға болады: а) оны ДНҚ - дан бөлумен; б) химиялық - ферментатитвті синтез жолымен; в) РНҚ тәуеді ДНҚ полимеразаның (ревертозаның) көмегімен, оқшауланған матрицалық РНҚ - ның негізінде жаңғыртумен. Гендерді ДНҚ - дан бөлу. Оқшауланған ДНҚ бөлшектеуге ұшыратады. Ол үшін, нуклеотидтердің белгілі тізбектеріне ие болатын, үлескілерде ДНҚ ыдырауын өршітетін, рестрипциялық эндонуклеозаларды пайдаланады. Қазіргі уақытта, 85 әртүрлі нуклеотидті тізбектерді танитын, 400 - ден астам рестриктазалар белгілі. Ыдырау нуклеотидті жұптардың танылатын үлескісінің ортасында жүруі мүмкін, сонда ДНҚ - дың екі тармақшасы да бір деңгейде «қиылады». Түзілген бөлшектер екітармақты (тұйық) ұштарға ие болады. Басқа рестриктазалар ДНҚ - ның тармақшаларын ығыстырумен ыдыратады, сондықтан баспалдақ түзіледі. ДНҚ жіпшелерінің шығыңқы болады. Біржіпшелі (жабысқақ) ұштары түзіледі. Бір рестриктазаның әрекетімен алынған, ДНҚ - дың екі жабысқақ бөлшегі кездессе, онда ұштық тізбектерінің әсерінен өзара әрекетте оңай түседі. Қажет болғанда тұйық ұштары жабысқақтарға айналуы мүмкін. Ол үшін, тұйық ұштарға, жабысқақ ұштар беретін, рестриктазаның тану үлескілерімен екітізбекті тізбектілерді қосады. Жабысқақ ұштарымен нуклеотидті тізбектілік болуы мүмкін: а) алдын - ала сол рестриктазамен өңделген, векторға қосылуы; б) өзара комплементар ұштарды тігу жолымен сызықтық молекуладан сақиналыққа түрленуі мүмкін. Рестриктазалардың көмегімен ДНҚ - дан гендерді әдісі мәнді кемшіліктерге ие болған. Қажет генге сәйкес келетін, сол бірүлескіні ДНҚ - дан кесіп алуға мүмкіндік беретін, рестриктазаларды таңдап алу қиын. Қызықты генмен қатар, ДНҚ бөлшектреі, генді қолдануға кедергілер келтіретін, артық нуклеотидті тізбектіліктерге де ие болады. Рестриктаза нуклеотидті ген тізбектілігінің бөлігін ыдырата алады, нәтижесінде ген функционалдық толық құндылығын жоғалтады.
Эупариотты ағзалардың гендері күрделі құрылысқа ие болған: кодтайтын ақуыздан, мәнді (экзондар) және аралық, мәнсіз үлескілерден (интрондар) тұрады. Осындай ДНҚ - матрицада синтезделген, алғашқы РНҚ түрөзгеріске ұшырайды. Нәтижесінде интрондарға сәйкес келетін, үлескілер жойылады, ал, экзондарға сәйкес келетін үлескілер, қосыла отырып, жетілген матрицалық РНҚ түзеді. Интрондардың болуы трансплантацияланған гендердің қалыпты қызметіне кедергі болып табылады. Рестриктазалармен ДНҚ - ын өңдеу барысында бөлшектердің қоспасы түзіледі. Бұлқоспадан, қажетті генге ие болатын, бөлшектерді бөліп алу - оңай мәселе емес. Бактериялық жасуша шамамен бес мың гендерден, ал эукариоттық жасуша 10 - 20 мың гендерден тұрады. Гендердің химиялық - ферментативті синтезі. Бұләдіс - нативті ДНҚ - дан рестриктазалардың көмегімен гендерді «қиюға» маңызды альтернатива. Әдіс нуклеотидтердің арасында эфир байланыстарының кезеңдік түзілуі есебінен, қысқа (8 - 16 буынды) біртізбекті ДНҚ бөлшектерінің химиялық синтезінен және екі тізбекті поленуклеотидтердің түзілуінен, ДНҚ лигаза арқылы өзара олегонуклеотидтердің тігісінен тұрады.
Химиялық - ферментативтік синтез нуклоетидтердің минимальді қажетті тізбектілігін дәл жаңғыртуға және ДНҚ бөлшектерінде артық нуклеотидті тізбектіліктердің, сонымен қатар, интрондардың элиминирленуімен байланысқан, мәселелерді болдырмауға мүмкіндік береді. Сонымен қатар, әртүрлі рестриктазалардың реттегішіне тізбектіліктердің т. б. тану үлескілерін гендерге енгізу мүмкіндігі бар.
Гендердің химиялық - ферментативті синтезіне оның нуклеотидті тізбектілігі туралы толық ақпарат қажет, сондықтан әдістің қолданысы осындай ақпаратты алу мүмкіндіктерімен шектелген. Генде нуклеотидтердің тізбектілігі сәйкесті ақуыздың алғашқы құрылымы негізінде жаңғыруы мүмкін. Ген құрылымын талдаудағы триумф - кодталатын ақуызда аминқышқылды қалдықтар тізбегінің және ДНҚ нуклеотидті тізбектілігінің параллель жаңғыруы. Химиялық - ферментативтік синтез әдісімен ас - оператор Е coli, проинсулиннің, инсулиннің А және В тізбектеріндегі, соматостатиндегі гендер алынған.
Жасушадан бөлінген матрицалық РНҚ негізінде гендердің ферментативті синтезі. Бұл гендер синтезінің ең танымал әдісі. Кері транскриптаза (ревертаза), мРНҚ - на комплементарлы, ДНҚ жіпшелерінің синтездерін өршітеді. Комплементарлық ДНҚ және кДНҚ деп аталатын, алынған біртізбекті ДНҚ - лып, ДНҚ - полимеразаны немесе ревертазаны қолданумен, ДНҚ - ның екінші жіпшесін синтездлеуге арналған матрица ретінде пайдаланады. Қарастырылатын әдістің артықшылығы, ген интрондарсыз және басқа транскрибцияланбайтын тізбектілерсіз алынады. Сонымен қатар, генді ДНҚ бөлшектерінің қоспасынан іріктегеннен гөрі, мРНҚ - ның қажет түрін жасуша аккумуляциялайтын, жағдайларды жасау оңайырақ. РНҚ тәуелді ДНҚ синтезінде негізделген, әдісті қолдануда 1979ж. адамның өсу гормонының генін алу үлкен жетістік болып табылады. Генді векторға енгізу. Сол немесе өзге тәсілмен алынған, ген ақуыз құрылымы туралы ақпаратты жүзеге асыра алмайды. Геннің әрекетін басқаратын қосымша механизмдер қажет, сондықтан генетикалық ақпаратты жасушаға тасымалдау векторлар құрамында жүзеге асырылады. Векторлар - өз бетінше реплткацияға қабілетті сақиналы молекулалар. Ген вектормен бірге рекомбинантты ДНҚ - ын түзеді. Рекомбинантты ДНҚ - ды құрастыру in Vitro жүзеге асырылады. Вектордың сақиналы молекуласы рестриктазамен айырылады. Алынған сызықты ДНҚ молекуласы, ДНҚ - ны енгізетін ұштарға комплементарлы, жабысқақ ұштарға ие болуы қажет. Енгізілетін геннің және вектордың комплементарлы жабысқақ ұштарын ДНҚ лигазаның көмегімен, біріңғай сақиналы молекула түзумен, қайта тұйықтайды. Векторлардың екі негізгі класын ажыратады: вирустар және плазмидтер. Генетикалық векторлар ретінде вирустарды пайдалану барысында туындайтын, маңызды мәселе - аттеньюация болып табылады. Аттеньюация - вируспен зақымдалған жасушалар аман қалуы және ұрпаққа өзгерген генетикалық бағдарламаны беруі үшін патогенділіктің әлсіреуі. Бүкілағза бойымен басты инфекция қысқа мерзімде дамитындай, жануар немесе өсімдік ұлпасында тарай отырып, жасушадан жасушаға тез тасымалданатын вирустардың қабілеті биотехнология үшін үлкен мәнге ие болған. Вирустардың осындай қасиеті ересек ағзадағы соматикалық жасушалардың генетикалық түрөзгеру мүмкіндігін ашады. Бұл қатынаста барлық он адам ағзасының жасышаларымен жетіспейтін гендерді таситын, вирустарды енгізу жолымен, адамның тұқым қуалайтын ауруларын емдеу жолдары ашылады.
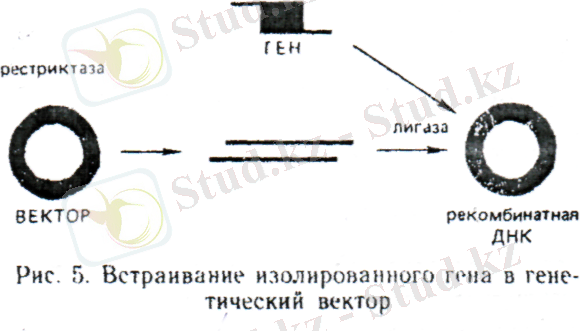
Плазмидтер - бактериялардан, саңырауқұлақтардан, өсімдіктерден және жануарлардан табыған, автономды өздігінен репликацияланатын генетикалық бірліктер. Генетикалық инженерияда бактериялық плазмидтер, әсіресе Е coli плазмидтер кең қолданыс тапқан.
... жалғасыСіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz